

서 언
재료 및 방법
식물재료 및 재배조건
CO2 고정률과 호흡률의 측정
OKJIP 분석 및 엽록소 형광변수 산출
생장량 측정 및 무름병 발생여부 조사
통계분석
결과 및 고찰
배추의 생육조건과 생육특성
CO2 고정률과 호흡률
OKJIP 곡선과 엽록소형광 변수의 변화
광계II에서의 에너지 흐름의 변화
고온에 의한 배추의 피해현황
서 언
식물은 동일 종이라도 생육 환경의 변화에 따라 형태적 또는 생리적으로 생장 상태가 크게 달라진다. 환경 요인 중에서도 온도는 식물의 지리적 분포와 생존, 그리고 생산량에 직결되는 중요한 요인으로 일정 수준보다 낮거나 높으면 저온 또는 고온 스트레스를 받게 된다. 즉, 온도가 내성의 한계에 가까워질수록 세포의 항상성을 저해하여 생명체의 성장 및 발달을 저해하게 되고, 심한 경우 결국은 죽음에 이르도록 한다. 특히 고등 식물에 있어서 종자나 화분과 같이 휴면 상태이거나 혹은 건조된 상태의 일부 특수한 조직들을 제외한 대부분의 조직들은 외부 온도가 45°C를 넘어가면 고온으로 인해 스트레스를 받게 되고 일정 시간 이상 노출되면 결국 죽게 된다(Levitt, 1980). 지구온난화는 식물의 광합성과 생장 기간을 변화시켜 식물의 생장과 발달에 직접적으로 영향을 주며 식량문제와도 연관이 있어, 현재 전 세계적으로 심각한 문제가 되고 있다. 기후 변화에 관한 정부간 패널 보고서(IPCC, 2007)에 의하면 지구온난화로 인하여 지구 전체의 평균 온도는 매 10년마다 0.3°C씩 상승할 것이며, 50-100년 후에는 현재의 기온보다 1.6-6.4°C가 높아질 것으로 예상하고 있다. 그러나 현 시점에서도 전 세계적으로 고온 스트레스에 의해 농작물들의 손실이 발생하고 있으며, 우리나라 역시 온대 기후에서 점차 아열대 기후로 변하면서 지역마다 재배되는 농작물이 점차 바뀌고 있다.
배추(Brassica campestris subsp. napus var. pekinensis)는 십자화과(Cruciferae)에 속하는 2년초 식물이며, 김치의 주재료로써 우리나라에서 생산 및 소비가 높은 채소 중에 하나이다. 최근 웰빙 붐과 더불어 김치의 식품영양학적 가치를 인정받아 국내뿐만 아니라 세계 시장에서도 그 수요가 증가하고 있다. 배추는 서늘한 기후를 좋아하는 호냉성 채소로 생육 적온인 20°C보다 높으면 결구 형성이 영향을 받고, 또한 무름병(soft-rot disease)을 발생시키는 세균(Erwinia carotovora subsp. carotovora)의 발생으로 인해 생산량이 감소하고 품질이 저하된다(Opeña et al., 1988). 더군다나, 배추는 Brassica속의 다른 식물종들에 비해 고온에 민감하며, 극심한 온도 스트레스는 되돌릴 수 없는 세포손상을 유발하여 세포 사멸을 초래한다(Yang et al., 2006). 따라서 안정된 배추의 생산을 위해서는 고온이 배추의 생육에 미치는 영향을 정량적으로 파악하고 고온 스트레스의 영향을 조기에 진단할 수 있는 기술을 개발할 필요가 있다.
최근 기후변화에 의한 식물의 반응을 파악하기 위하여 작물의 재배적지 변화, 수확량 변동예측 모형 개발 등에 관한 연구들이 활발하게 이루어지고 있다. 그리고, 폐쇄형 챔버(closed chamber) 또는 반폐쇄형 챔버(open-top chamber) 등을 이용하여 변화된 온도 또는 CO2 농도 하에서의 작물의 생장과 발달, 수확량 등의 변화 등에 관한 연구들도 진행되고 있다(Hadley et al., 1995; Porter and Semenov, 2005). 그러나 이러한 시스템은 광량 등의 미세 기후를 실제 대기조건과 유사하게 만들기에는 다소 어려움이 있다. 반면에 온도구배터널(temperature gradient tunnel system)은 온도를 제외한 다른 환경인자(광량, 대기 CO2 농도 등)를 자연 상태와 유사하게 유지할 수 있도록 고안한 반폐쇄형 장치이다. 즉, 입구를 개방하여 외부 공기가 터널 내부로 유입되도록 하고, 후미부는 6개의 소형 환기팬을 설치하여 배기 속도를 제어함으로써 공기 흡입구에서부터 후미부 쪽으로 연속적인 온도 구배가 형성되도록 설계되어 고온에 대한 작물의 반응을 연구하는데 유용한 시스템이다.
한편, 고온과 같은 환경 스트레스는 직・간접적으로 광계II(Photosystem II)와 같은 광합성 기구에 손상을 줄 수 있어(Takahashi and Murata, 2008; Yoshioka et al., 2006), 고온에 노출된 작물은 광합성량의 감소로 생산량이 감소될 수 있다. 광계II의 반응을 조사하기 위해서는 엽록소형광 분석이 다양하게 이용되고 있으며, 이는 환경 스트레스에 대한 식물의 생리적 반응 및 광합성 기구에 대한 다양한 정보를 제공한다(Strasser and Strasser, 1995). OKJIP 곡선은 빛을 조사한 잎에서 방출되는 형광의 시간대별 밀도를 나타내는 엽록소형광곡선으로 O, K, J, I, P 단계에서의 형광밀도로부터 환경 스트레스 하에서 일어나는 광계II에서의 에너지 흐름의 변화와 전자수용체의 환원 등 다양한 정보를 정량적으로 얻을 수 있다(Strasser et al., 2000).
따라서 본 연구에서는 온도구배터널에서 재배되고 있는 배추의 잎에서 광합성적 CO2 교환과 엽록소형광을 분석함으로써 배추의 생장에 미치는 고온의 영향을 정량적으로 분석하고, 고온의 영향을 진단하는 도구로서 엽록소형광 변수들의 활용 가능성을 살펴보았다.
재료 및 방법
식물재료 및 재배조건
본 실험에 사용한 식물재료는 봄재배용 배추인 ‘대통’(B. campestris subsp. napus var. pekinensis cv. Detong)이며, 2월 하순에 배양용 상토가 들어 있는 105구 육묘용 플러그 트레이에 종자를 1립씩 파종하여 자연광 하에서 20 ± 1°C (day)/15 ± 1°C(night)의 조건으로 재배하였다. 파종 30일 후(3월 하순)에 본엽이 3-4매 나온 균일하게 자란 개체들을 선발하여 온도구배터널에 정식하고, 정식된 배추는 농촌진흥청 표준영농재배법에 준하여 관리하였다. 온도구배터널(25m × 3m × 3m) 내의 토양은 정식 3일 전에 밑거름(퇴비와 복합비료)을 시비하여 개량하였다. 토양수분은 조사기간 동안 2-3일 간격으로 관수하여 포장용수량 정도를 유지하였으며, 배추좀나방, 진딧물 등 병충해 방제를 위하여 재배기간 중 4회에 걸쳐 저독성 약제를 살포하였다. 정식 후 5주째(4월 하순)에 각 처리구에서 자라는 배추의 8-10번째 잎 중에서 엽색이 비슷하고 균일한 상태의 잎을 실험재료로 사용하였다. 온도구배터널 내부의 온도는 터널의 입구, 중앙부, 후미부에 지상부로부터 2.5m 높이에 설치한 온도센서(1400-101, LI-COR Inc., Lincoln, USA)를 이용하여 측정하였다. 시험기간 동안 대기온도를 유지하는 터널 입구에 비해 중앙부, 후미부에서는 온도가 각각 4°C와 7°C 더 높게 유지되도록 복합환경제어시스템(TGC-Soldan, Soldan Crop., Korea)을 이용하여 제어하였다. 일사량은 지상부로부터 2.5m 높이에 설치한 광센서(LI200X, LI-COR Inc., USA)를 이용해 측정하고 일일적산일사량으로 나타내었다. 재배기간 동안의 온도와 일사량은 데이터로거(CR1000, Campbell Scientific Inc., Logan, USA)에 연결하여 1시간 간격으로 기록하였다.
CO2 고정률과 호흡률의 측정
CO2 교환은 휴대용 CO2 분석장치(LCpro+ Portable Photo-synthesis System, ADC BioScientific Ltd., Hoddesdon, UK)를 사용하여 맑은 날 14시부터 16시 사이에 측정하였다. CO2의 공급은 4m 높이의 대기 유입 안테나를 사용하여 220 mol・m-2・s-1의 유속으로 깨끗한 공기를 공급하는 것으로 대신하였다. CO2 고정률(photosynthetic rate)은 잎 챔버 내 CO2 농도가 대기의 CO2 농도와 비슷해지면 배추의 잎을 챔버에 물리고 5분간 안정화시키고 1,200μmol・m-2・s-1의 포화광을 비추어 측정하였다. 호흡률(dark respiration)은 CO2 고정률과 같은 방법으로 측정하였다. 다만, 포화광을 비추는 대신에 암조건을 유지하였다. 아울러 기공전도도(stomatal conductance), 증산률(transpiration rate) 등을 측정하고, 기공전도도에 대한 CO2 고정률로 수분이용효율(water use efficiency)을 산출하여 제시하였다.
OKJIP 분석 및 엽록소 형광변수 산출
OKJIP 곡선은 엽록소형광분석기(Plant Efficiency Analyzer, Hansatech Instrument Ltd., King’s Lynn, UK)를 이용하여 분석하였다(Strasser and Strasser, 1995). 15분간 광을 차단하여 암적응시킨 배추의 잎에 1,500μmol・m-2・s-1의 광량을 5초간 조사하여 OKJIP 곡선을 유도하였다. Time scale은 10μs에서 1s의 범위로 설정하고, 10μs에서 2ms까지는 1μs 단위로, 2ms에서 1s까지는 1ms 단위로 인식하도록 하였으며, 50μs(O-단계), 300μs(K-단계), 2ms(J-단계), 30ms(I-단계), 500ms(P-단계)에서 엽록소형광 밀도를 분석하였다(Srivastava et al., 1997). OKJIP 곡선의 분석으로 기본적 형광변수(FO, FK, FJ, FI, FM 등)들을 추출하고, 이로부터 다양한 형광변수들을 산출하였다(Table 1). 그리고, 에너지의 현상학적 흐름을 나타내는 변수들을 이용하여 광계II의 반응중심을 통한 에너지 흐름을 pipeline model로 제시하였다.
생장량 측정 및 무름병 발생여부 조사
배추의 생장량은 정식 후 5주째에 대기온도를 대조구로 하고 대기온도 + 4°C, 대기온도 + 7°C 조건에서 각각 10개체를 선정하여 엽수와 엽길이를 측정하였으며, 엽록소계(SPAD-502, Minolta Co., Ltd., Osaka, Japan)를 사용하여 SPAD값을 측정하였다. 엽수는 잎의 길이가 1cm 이상인 것을 조사하였으며, 엽길이와 SPAD값은 각 개체의 8-10번째 잎을 대상으로 조사하였다. 그리고, 고온에 의한 피해는 결구 형성 시(정식 후 7주, 9주와 10주째) 배추 포기를 세로로 2등분하여 내부 단면을 관찰하여 무름병의 발생여부를 확인하였다. 정식 후 10주째에 수확한 배추는 결구 부분과 비결구 부분으로 분리하여 생체중을 측정하였다.
통계분석
통계분석은 SPSS 통계 패키지 Release 7.5(SPSS Inc., Chicago, USA)를 이용하여 수행하였다. OKJIP 곡선의 분석으로부터 산출된 형광변수들은 Duncan의 다중검정(p < 0.01)으로 평균치간의 차이에 대한 유의성을 검정하였다.
결과 및 고찰
배추의 생육조건과 생육특성
배추의 생육 기간 동안 온도구배터널 내부의 일일적산일사량은 비가 내리거나 흐린 날을 제외하고는 전반적으로 4,664W・m-2(로제트 단계 4,187W・m-2, 결구형성 단계 4,989 W・m-2)를 상회하였으며, 일평균 대기 온도는 터널 입구의 대기온도를 기준으로 중앙부, 후미부의 온도가 주야를 통하여 각각 4°C, 7°C가 높게 유지되고 있는 것을 확인 할 수 있었다(Fig. 1). 배추의 생장 및 발달에 있어서 로제트 단계에서는 터널 입구의 일평균 대기온도가 11.4°C(일최저평균 5.0°C, 일최고평균 20.7°C)를 나타내었으며, 터널의 중앙부에서는 일평균 15.0°C(일최저평균 7.7°C, 일최고평균 24.3°C), 후미부에서는 일평균 17.5°C(일최저평균 9.7°C, 일최고평균 25.4°C)로 터널 입구에 비해 각각 3.6°C와 6.1°C가 높았다. 그리고 결구형성 단계에서는 터널 입구의 일평균 대기온도는 18.5°C(일최저평균 11.8°C, 일최고평균 23.5°C)로 로제트 단계보다 7.1°C나 높았다. 터널의 중앙부에서는 일평균 22.2°C(일최저평균 15.8°C, 일최고평균 26.9°C), 후미부에서는 일평균 24.6°C(일최저평균 17.7°C, 일최고평균 29.2°C)로 로제트 단계에 비해 각각 7.2°C, 7.1°C가 높았다. 그리고 재배기간 동안 터널 입구에는 일평균 25°C 이상을 넘는 날이 없는데 반하여 중앙부는 25°C 이상의 일수가 7일, 후미부는 16일이었다.
배추의 생장은 정식 후 4주째까지는 결구를 형성하지 않고 잎이 밖으로 펼쳐지고, 재배기간이 길어지면서 점차 엽수가 증가하고 잎이 안쪽으로 포개지면서 서서히 결구가 형성되었다. 결구는 대기온도에서 자라는 배추보다는 대기온도 + 4°C 조건에서 자라는 배추에서 오히려 빨리 형성되는 것을 확인할 수 있었다. 온도구배터널에 정식한 후 5주째인 4월 하순에 엽수와 엽길이 등의 생육에 미치는 온도의 영향을 살펴보았다(Table 2). 그 결과, 대기온도보다 대기온도 + 4°C와 대기온도 + 7°C 조건에서 엽수가 다소 많아지고, 엽길이도 더 길었다. 즉, 정식 5주째에 대기온도에서 자란 배추의 엽수는 14.2개인데 반해 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 각각 16.2와 16.1개로 많았으며, 엽길이도 대기온도에서 자란 배추가 16.2cm이나 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 각각 21.9cm와 24.1cm로 더 길게 자라 빠른 신장을 확인할 수 있었다. 그리고, SPAD값은 대기온도에서 44.0이고, 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 각각 45.3과 41.7로 온도가 높아졌을 때 다소 감소하나 통계적으로 유의한 차이는 없었다.
CO2 고정률과 호흡률

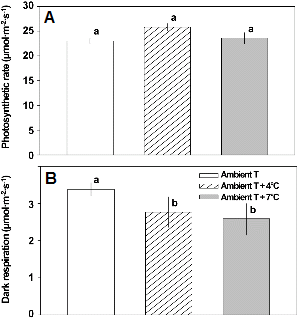
식물 잎의 CO2 고정률은 광합성 수준을 직접적으로 반영한다고 할 수 있으며, 작물의 재배나 생육 환경에 있어 스트레스로 작용할 수 있는 온도, 광 등의 환경 스트레스에 의한 식물의 광합성능을 평가하는 지표로 중요하다. 그리고 CO2 고정률과 더불어 호흡률은 고온에 대한 민감성 식물을 선별하는데 있어서도 중요하다(Prange et al., 1990). 배추의 CO2 고정률은 대기온도에서 자란 잎에서 25.8μmol・m-2・s-1로 다소 높았으나, 온도에 따라 통계적으로 유의한 차이는 없었다(Fig. 2A). 반면에 호흡률은 대기온도에서 다소 높았으며, 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 낮았다(Fig. 2B). 이와 유사한 결과로 상승된 온도에 순화된 활엽초본, 목초, 상록성 교목이나 관목 등에서도 일정 범위(14-28°C)까지는 CO2 고정률이 차이가 없거나 증가하는데 반해, 호흡률은 전반적으로 감소하는 것으로 보고된 바 있다(Campbell et al., 2007). 그리고, 포플러(Populus tremuloides), 자작나무(Betula papyrifera), 미국낙엽송(Larix laricina), 방크스소나무(Pinus banksiana), 가문비나무(Picea mariana) 등의 한대성 식물도 상승된 온도에서 자랐을 때 18°C보다 30°C에서 호흡률이 더 감소하였다(Tjoelker et al., 1999). 많은 식물 종들에 있어서 호흡률은 대기온도에 의존적으로 변화하는데, 특정 온도보다 따뜻한 조건에서 자랐을 때에는 호흡률이 감소하고 차가운 조건에서 자랐을 때에는 증가하는 양상을 보인다(Atkin and Tjoelker, 2003). 본 연구의 대기온도 + 4°C와 대기온도 + 7°C 조건에서 배추의 바깥 쪽에 붙어있는 잎에서 관찰되는 호흡률의 감소도 상승된 온도에 순화된 결과인 것으로 보인다.
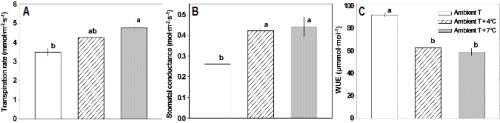
한편, 온도에 대한 기공의 반응은 식물 종이나 동일 종이더라도 재배 환경에 따라 매우 다양하게 나타난다. 다시 말하면 온도 변화에 따라 기공전도도가 증가 또는 감소하거나 다소 둔감하기도 한다(Silim et al., 2010). 본 연구에서는 대기온도보다 + 4°C와 + 7°C 상승하였을 때 증산률과 기공전도도가 더 높았으며, 수분이용효율은 오히려 낮았다(Fig. 3). 양구슬냉이(Camelina sativa)와 담배(Nicotiana tabacum)도 높은 온도에서 자랐을 때 CO2 고정률은 감소하나 증산률과 기공전도도가 증가하는 것으로 나타났다(Carmo-Silva and Salvucci, 2012). 특히, 양구슬냉이는 기공전도도와 증산률이 순화 온도의 상승에 따라 점차 증가하여 최적온도(30-32°C) 이상의 다소 높은 온도(38°C)에서도 높게 유지되었다(Carmo- Silva and Salvucci, 2012). 그리고 수분이용효율은 고온과 같은 스트레스 하에서 감소하는데, 배추와 같은 속 식물인 갓(B. juncea L.)에서도 30°C와 40°C의 고온에서 수분이용효율이 감소하는 것으로 알려지고 있다(Hayat et al., 2009). 이러한 결과는 배추를 다소 높은 온도에서 재배할 때에는 기공전도도와 증산률이 증가하기 때문에 수분이용효율이 감소하여 수분요구량이 많아질 수 있음을 나타내주고 있다.
OKJIP 곡선과 엽록소형광 변수의 변화
식물체가 빛, 온도, 수분 또는 화학적 스트레스 등에 의해 영향을 받았을 때, OKJIP 곡선의 패턴이 변하게 된다(Lu and Zhang, 1999; Mathur et al., 2011; Oh and Koh, 2013). 온도구배터널에 정식한 후 5주째인 4월 하순에 배추 잎에서 OKJIP 곡선의 변화와 이를 토대로 형광변수들을 산출하여 제시하였다(Figs. 4 and 5). 대기온도 + 4°C와 대기온도 + 7°C 조건에서 자란 배추의 잎에서는 대조구인 대기온도에 비해 O와 K 단계에서의 형광밀도 FO와 FK가 증가하고 J, I, P 단계에서의 형광밀도 FJ, FI, FP가 감소하고 있으며, 특히 P-단계에서의 형광밀도가 크게 감소하였다(Fig. 4). 이와 같이 온도가 높아짐에 따라 FJ, FI, FP가 크게 낮아지는 것은 고온에 의해 배추 잎의 광계II 반응중심의 전자수용체 quinone이 환원상태에 있고 광계II의 광화학적 효율이 감소하고 있음을 의미한다(Lu and Zhang, 1999, 2000). 그리고, 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 특이적으로 FK가 뚜렷하게 증가하는 것을 확인할 수 있었다. 이는 산소발생복합체(OEC; oxygen-evolving complex)의 불활성으로부터 기인한 광계II의 이차 전자공여체에 전자전달이 저해된 결과로 판단된다(Guisse et al., 1995; Strasser, 1997). 이러한 결과는 밀(Tritium aestivum L.)에서도 25°C에서 생육한 잎에서는 FK가 나타나지 않지만 45°C의 고온에 노출된 잎에서는 FJ, FI, FP값의 감소와 더불어 FK가 뚜렷하게 나타나고, 37.5°C 이상의 온도에서 FK가 점차 증가하는 것으로 보고된 바 있다(Lu and Zhang, 2000; Mathur et al., 2011).
OKJIP 분석을 통해 산출된 형광변수들 중에 FM, FV/FO는 온도가 높아졌을 때 점차적으로 감소하였고, FO, FO/FM, MO, SM, VJ는 증가하였다(Fig. 5A). 광계II의 광양자 효율을 나타내는 ΦPO, ΨO와 ΦEO는 온도가 높아졌을 때 크게 감소하였다. 그리고, 식물의 활력도를 나타내는 SFIabs, PIabs, KP, RC/ABS도 온도가 높아질수록 점차 감소하였으며, 역의 식물 활력도를 나타내는 KN은 증가하였다(Fig. 5B). 형광변수들 중 FV/FO는 활성상태인 반응중심의 크기와 밀도를 나타내는 지표이므로 Fig. 5A에서 대기온도 + 7°C 조건에서 FV/FO가 크게 감소하는 것은 고온에서 불활성상태의 광계II 반응중심이 증가하고 있음을 암시한다(Kriedemann et al., 1985). 40°C 이상의 고온 장해를 받은 밀(T. aestivum L.)에서도 FV/FO가 뚜렷하게 감소하였다(Mathus et al., 2011). 그리고, 40°C 이상의 고온 스트레스를 받은 완두(Pisum sativum L.)의 잎, 사과(Malus domestica Borkh.)와 토마토(Solanum lycopersicon cv. Mini Carol)의 잎과 과피에서도 본 연구결과와 유사하게 FO, MO, SM, VJ 등의 형광변수들이 증가하는 것으로 보고되었다(Chen and Cheng, 2009; Strasser et al., 2000; Zushi et al., 2012). 뿐만 아니라 광계II의 광양자 효율을 나타내는 ΦPO, ΨO와 ΦEO도 온도가 높아졌을 때 감소하였으며, 대기온도 + 7°C에서 급격하게 감소하였다. 이러한 결과는 식물이 고온 스트레스에 처했을 때 불활성상태의 반응중심이 많아지고 포획된 에너지의 대부분이 전자전달로 보내어지지 않고 있음을 간접적으로 보여주고 있다고 할 수 있다. 온도변화에 있어서 배추와 유사한 CO2 고정률과 기공반응을 보인 양구슬냉이에서도 35°C 이상의 온도에서 Fv/Fm값이 급격하게 감소하였으며, 이때 Fo의 증가를 동반하는 것으로 나타났다(Carmo-Silva and Salvucci, 2012). 그리고, 고온에 민감한 온주밀감(Citrus unshiu Marc.)도 감수성 식물인 네블오렌지(C. sinensis Osbeck)보다 38°C의 고온 조건에서 FV/FM의 감소와 FO의 증가가 확연하게 나타나는 것으로 알려졌다(Guo et al., 2006). 따라서 본 연구에서 배추가 대기온도 + 7°C 조건에서 ΦPO(= FV/FM)값이 급격하게 감소하였으며, 이때 Fig. 5A에서 보는 바와 같이 FM의 감소와 FO의 증가를 동반하고 있어 고온에 의해 만성적인 광억제가 일어나고 있음을 알 수 있다.

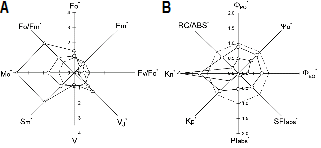
Table 2에서 5주째에 측정한 배추의 생장은 대기온도보다 + 4°C 또는 + 7°C 높은 온도에서 더 좋았다. 이러한 배추의 생장은 온도구배터널에 정식한 후 각각의 온도에서 5주 동안 자란 결과임을 감안해 보았을 때에 배추의 초기 생장 시, 즉 정식 후 5주째까지는 높은 온도가 오히려 잎의 생장에 도움을 준 것으로 보인다. 하지만, 5주째에 이르러서 상승 온도가 광합성에는 영향이 없었지만 호흡을 감소시키고 증산률이나 기공전도도의 증가, 그리고 수분이용효율의 감소를 야기하고(Figs. 2 and 3), 상승 온도에서 고온 스트레스 하에서 특이적으로 나타나는 현상인 FJ, FI, FP의감소,FK의 증가와 FV/FO값의 감소 등이 나타나고 있어(Figs. 4 and 5), 5주째에 이르러서는 고온이 광합성 기구, 특히 광계II에 악영향을 미치고 있음을 알 수 있었다. 이러한 결과는 OKJIP 곡선 패턴에서 상승 온도에서의 FJ, FI, FP의감소,FK의 증가와 FV/FO값의 감소를 비롯하여 고온에서 뚜렷하게 증가하거나 감소하는 MO, SM, RC/CS, ETo/CS, PIabs, SFIabs 등의 형광변수들이 배추의 생육 시 고온 스트레스를 감별하는 지표로 활용될 수 있는 가능성을 보여주고 있다. 이들 형광변수들은 기존에 사용되고 있는 변수인 FO, FV/FM와 FV/FO 등과 더불어 식물이 스트레스를 받았을 때 그 증상을 육안으로 확인할 수 없을 때에도 정량적으로 판정할 수 있기 때문에 유용하게 쓰일 것으로 보인다.

광계II에서의 에너지 흐름의 변화
대기온도를 기준으로 4°C와 7°C 상승하였을 때의 RC/CS와 광계II에서의 에너지 흐름의 변화를 비교하여 보았다(Table 3). RC/CS는 대조구에 비해 온도가 높아지면 크게 감소하여 엽면적당 활성형 반응중심의 밀도가 낮아져서 대기온도 + 4°C와 대기온도 + 7°C에서 대조구에 비해 각각 61.3%와 13.6%로 감소하였다(Table 3). 본 결과와 유사하게 수수(Sorghum bicolor)의 재배에 있어서도 고온에 의해서 RC/CS의 감소가 나타나는 것으로 보고된 바 있다(Yan et al., 2012). 그리고 ABS/CS, TRo/CS와 ETo/CS도 온도가 높아짐에 따라 점차 줄어들었으며 엽면적당 흡수되는 빛에너지 양과 광계II에서 포획되는 에너지, 전자전달로 전해지는 에너지가 모두 감소하고 있음을 알 수 있다. 이에 반해 DIo/CS는 온도가 높아짐에 따라 증가하였으며, 대조구인 대기온도에 비해 대기온도 + 4°C와 대기온도 + 7°C 조건에서 각각 1.1배, 1.41배 증가하였다. 이는 전자전달로 사용되지 않는 에너지가 많음을 나타내주고 있다. 이러한 광계II에서의 에너지 흐름을 pipeline model로 나타내면 Fig. 6과 같다. 본 연구에서 배추가 고온 스트레스에 노출되어 전자전달에 쓰이는 에너지가 줄어들고 있음을 한눈에 알아볼 수 있다.

고온에 의한 배추의 피해현황
배추는 서늘한 기후를 좋아하는 호냉성 채소로 생육기에 고온이 되면 결구가 되지 않을 뿐만 아니라 여러 병과 해충들에 의해 피해를 입게 된다(Opeña et al., 1988). 특히 고온에서 가장 큰 문제가 되고 있는 질병들 중에 하나가 E. carotovora subsp. carotovora에 의해 발병하는 세균성 무름병이다. 무름병균은 펙틴분해효소를 생산함으로써 배추의 세포벽을 와해시켜 식물조직을 무르게 하며 결국에는 세포를 죽게 만든다(Chaterjee et al., 1994). 본 연구에서 온도구배터널에 정식한 후 7주, 9주와 10주 동안 자란 배추를 수확하여 고온의 피해여부를 조사하였다(Fig. 7). 대조구인 대기온도에서는 7주, 9주, 10주에 수확한 배추 내부에서 병징이 관찰되지 않았으나, 대기온도 + 4°C와 대기온도 + 7°C에서는 재배기간이 길어질수록 점차 무름병에 의한 피해가 두드러지게 관찰되었다. 특히 7주간 재배하였을 때 대기온도 + 7°C 조건에서 결구 안쪽의 식물조직에 황갈색의 반점이 생겨 조직이 물러지는 것을 확인할 수 있었으며, 9주 후에는 그 범위가 점차 확대되면서 포기 전체가 엷은 누런색으로 변하며 흐물흐물하게 썩어 심한 악취를 발산하였다. 이는 결구형성 단계에서 고온에 의해 야기되는 피해로 보이는데, Fig. 1에서 보는 바와 같이 대기온도 + 7°C 조건에서 결구형성 단계에서 25°C 이상의 일수가 16일로 가장 많을 뿐 아니라 온도가 최고 29.2°C까지 높은 사실로 유추해 볼 수 있다. 그리고 대기온도 + 4°C 조건에서도 9주 후에 병징이 나타났다. 이러한 결과는 그리 높지 않은 고온이라 할지라도 장기간 동안 노출되면 결구가 형성되는 시기에 무름병의 발생을 야기하고, 이로 인해 품질이 저하될 수 있음을 나타내주고 있다. 따라서 결구가 형성되는 시점부터는 일 최고 온도가 25°C를 벗어나지 않는 환경에서 재배하는 것이 중요한 것으로 보인다. 감자 재배에 있어서도 대기온도보다 3°C 높아지면 역병이 증가되어 감자 수확량이 큰 폭으로 감소하는 것으로 알려지고 있다(Kaukoranta, 1996).
온도구배터널에 정식한 후 10주째에 배추를 수확하고 생체중과 무름병의 발생여부를 살펴보았다(Table 4). 결구 부분의 생체중은 대기온도에서 높고 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 다소 낮았다. 반면에 비결구 부분의 생체중은 대기온도에서 다소 낮고 지상부 전체의 생체중은 대기온도 + 7°C 조건에서 다소 높았지만, 처리간에 유의성을 보이지는 않았다. 무름병의 발생은 대기온도에서는 나타나지 않아 배추가 건전한 상태를 유지하였으나, 대기온도 + 4°C와 대기온도 + 7°C 조건에서는 무름병 발생률이 각각 70%와 100%로 고온에 의한 피해가 나타나고 있음을 확인할 수 있었다. 일반적으로 온도의 상승은 식물의 초기 생장과 발달에 있어서 우호적으로 작용한다(Ge et al., 2012). 배추 역시 서늘한 기후를 좋아하는 호냉성 채소이기는 하나 생육초기에는 높은 온도에서도 비교적 잘 견디는 것으로 알려지고 있다(Opeña et al., 1988). 본 연구에서도 Fig. 1과 Table 2에서 살펴본 바와 같이 대기온도에서보다 대기온도 + 4°C와 대기온도 + 7°C 조건이 배추 생육의 초기 단계에서는 우호적으로 작용하여 엽수와 엽길이, 결구형성 등에 있어서 긍정적인 효과를 나타내었다. 하지만 수확단계에 이르러서는 Table 4에서와 같이 대기온도 + 4°C와 대기온도 + 7°C 조건이 생체중의 감소와 더불어 무름병 발생률이 증가하는 등의 악영향을 미치고 있음을 확인할 수 있었다.
이상의 결과를 종합해 보았을 때 급격하게 변화하는 미래의 기후 환경 하에서 다른 작물들에서도 예측되는 바이나 배추도 역시 고온에 노출될 가능성이 매우 높다고 할 수 있다. 따라서 배추의 안정적인 생산을 위해서는 고온 적응성 품종, 특히 결구 시점에서 내고온성이 강한 품종을 육성하거나 고온의 피해를 최소화할 수 있는 재배기술이 확립되어야 할 것으로 보인다. 그리고 결구형성 시 고온 스트레스의 영향을 조기감별하기 위해서 OKJIP 곡선에서 FK의 증가를 비롯하여 기존에 사용되고 있는 변수인 FO, FV/FM와 FV/FO 이외에도 MO, SM, RC/CS, ETo/CS, PIabs, SFIabs 등의 형광변수들을 유용하게 활용할 필요가 있을 것으로 보인다.
