

서 언
제비동자꽃은 Lychnis속 식물로 석죽과(Caryophyllaceae)에 속하는 다년생 초본류이다. 세계적으로 약 30여종이 분포하며(Bae et al., 2014), 동아시아와 유럽전역에 걸쳐 북반구에 자생한다(Magnus et al., 2008). 우리나라에 자생하는 Lycnis속 식물은 제비동자꽃[ L. wilfordii (Regel) Maxim], 동자꽃(L. cognata Max.), 털동자꽃(L. fulgens Fisch.), 그리고 가는동자꽃(L. kiusiana Makino) 등 4종이 보고되어 있다(Lee, 1974). 특히 제비동자꽃은 산림청 지정 위기종(endangered species, EN)으로 지정되어 있으며, 강원도 일부 지역에서 자생하지만 개체수가 매우 적기 때문에 자생지에 대한 지속적인 모니터링 및 대량증식을 통한 현지 내외 보전대책이 필요한 실정이다(Lee et al., 2012).
제비동자꽃은 7-8월에 짙은 주황색의 꽃이 개화하며, 관상용으로서 가치가 높아 분화 및 정원용으로 유망한 식물이다(Lee and Yoon, 1998). 그러나 제비동자꽃의 자생지 내 개체수가 매우 적고 산업적인 활용에 한계가 있어 대량증식법 개발을 위한 몇 가지 연구가 이루어져 왔다. Bae et al.(2014)는 제비동자꽃의 기내번식에 대한 연구에서 100mg·L-1 GA3가 첨가된 MS배지에서 발아율이 50% 미만이었고, 발아소요기간은 2-3개월 정도였다고 발표하였다. 이 연구에서 조직배양을 통한 제비동자꽃의 기내번식에 성공하였으나 배양방법이 까다롭고 숙련된 기술을 요구한다는 단점이 있다. 제비동자꽃의 실생번식 연구에서 Baskin and Baskin(2014)이 북미지역에 자생하는 동일 속의 L. alba 종자가 생리적 휴면을 가지고 있으며, 암조건에서는 발아가 저조하지만 명조건에서는 발아율이 높다고 보고하였다. 또 다른 종인 L. flos-cuculi는 주야간 온도가 20/8°C의 조건에서 일년 동안 배양하였을 때 약 10%가 발아되었다고 보고된 바 있다(Milberg, 1994). L. senno Siebold et Zucc.의 경우 종자나 근경번식을 하는데, 종자가 발아하기까지 5-6개월의 긴 시간이 소요될 뿐만 아니라, 발아율 또한 매우 낮다. 기내배양에서 250mg·L-1 GA3 처리를 통하여 19.4%의 낮은 발아율을 보였다고 보고된 바 있다(Chen et al., 2006).
종자휴면은 식물의 생존전략으로써 생육에 적합하지 않은 환경에서는 발아를 억제하고 생육에 적합한 환경에서 발아시켜 추후 식물의 생존율을 높일 수 있게 한다(Baskin and Baskin, 1998; Fenner and Thompson, 2005). 현재 종자휴면의 종류는 크게 다섯 가지로 분류되어 있는데, 생리적 휴면(physiological dormancy, PD), 형태적 휴면(morphological dormancy, MD), 형태생리적 휴면(morphophysiological dormancy, MPD), 물리적 휴면(physical dormancy, PY), 그리고 조합 휴면(combinational dormancy; PY+PD)으로 나뉘며, 각각의 휴면유형 내에서도 깊이에 따라 세분화되어 있다(Baskin and Baskin, 2004). Baskin and Baskin (2014)는 Lychnis속 식물들을 PD로 분류하였다. PD는 다섯 가지의 휴면유형 중 온대지방에서 가장 일반적인 유형이며 나자식물문(Coniferales, Gnetales)과 대부분의 피자식물문 계통에서 주로 나타난다(Baskin and Baskin, 2004). 이러한 PD는 저온층적(cold stratification), 후숙(after-ripening) 또는 GA(gibberellic acid) 처리에 의해 휴면이 타파되며(Lee et al., 2015), 휴면 깊이에 따라 deep PD, intermediate PD, 그리고 non-deep PD 총 세 가지로 분류된다(Nikolaeva, 1977). Nikolaeva (2001)에 따르면 deep PD는 Malus속, Sorbus속, Acer속 등에서 나타나며, GA 처리는 종자의 휴면을 타파시키지 못하지만 3-4개월의 저온 또는 고온습윤처리를 통하여 휴면타파가 가능하다(Baskin and Baskin, 2004). Intermediate PD에는 Fraxinus lanceolate, A. negundo, Polygonum convolvulus, Stachys alpine 등에서 주로 나타나는 휴면 유형이며(Nikolaeva, 2001), GA 처리를 통한 휴면 타파는 일부 종에 국한되며, 주로 2-3개월의 저온층적처리를 통하여 휴면타파가 가능하다(Baskin and Baskin, 2004). 또, intermediate PD를 갖는 종자들은 후숙처리와 같은 건조저장을 통해 휴면 타파를 위한 저온요구도를 일부 충족시킬 수 있다(Baskin and Baskin, 2004). Non-deep PD는 가장 흔한 휴면 유형으로 1-2개월의 저온층적처리, 후숙, GA3 처리 등 모든 처리에 의해 종자의 휴면타파가 가능하다고 알려져 있다(Baskin and Baskin 2004; Geneve 2003).
산림청 지정 위기종인 제비동자꽃은 근경을 분주하거나 조직배양으로 번식할 수 있으나, 실용적인 측면에서는 종자번식이 가장 적합하다고 할 수 있다. 따라서 본 연구는 제비동자꽃의 종자휴면 유형을 분류하고 휴면타파 및 발아 촉진법을 개발하여 유전자원 보존 및 증식기술을 확립하고자 수행하였다.
재료 및 방법
실험 재료
본 연구에서 사용된 제비동자꽃[L. wilfordii (Regel) Maxim] 종자는 2015년 8월 4일 경기도 양평군 소재의 국립수목원 유용식물증식센터에서 현지외 보존되고 있는 식물체에서 채종하였다. 채종한 종자는 빛이 없고 통풍이 잘되는 곳에서 5일 동안 상온 건조 후 정선하여 바로 실험에 사용하였다.
종자 기본 특성조사
종자의 내외부 형태관찰을 위해 양면면도칼(stainless blade, Dorco, Seoul, Korea)을 이용하여 종자를 반으로 잘랐다. USB현미경(AM 3111 Dino-Lite premier, AnMo Electronics Co., Taiwan)을 이용하여 50-60배율로 촬영하였으며, 종자의 탈리시점과 발아과정의 내외부 형태를 관찰하였다.
그리고 제비동자꽃 종자가 불투수성과 물리적 휴면을 가지고 있는지 알아보기 위하여 수분흡수율을 측정하였다. Petri dish에 여과지(Whatman No.1, GE Healthcare, Buckinghamshire, UK) 두 장을 깔고 증류수를 첨가한 후 100립씩 3반복으로 배양하였다. 수분 흡수 전 초기 무게와 배양 후 3, 6, 9, 12, 24, 36시간이 경과되었을 때 무게를 측정하였다. 그리고 수분 흡수 정도는 %Ws = [(Wh-Wi)/Wi] x 100의 계산식을 이용하여 산출하였 다(Lee et al., 2015). Ws = 증가된 종자 무게, Wh = 수분 흡수 후 경과 시간 때의 종자 무게, Wi = 종자 초기 무게.
종자 소독 및 배양
종자는 petri dish에 치상하기 전 500mg·L-1의 소독제(Benomyl, FarmHannong, Seoul, Korea)에 30분간 침지 소독한 다음 증류수로 5회 이상 수세한 후 치상하였다. 모든 처리구는 90×20mm petri dish에 여과지 2매를 깔고 여과지가 마르지 않을 정도의 증류수를 주입 후 30립씩 3반복으로 치상하였고, 명조건과 암조건으로 나누어 처리하였다. 명조건은 26.13μmol·m-2·s-1 PPFD의 광을 16시간 조사하였고, 암조건은 petri dish를 aluminum foil로 싸서 완전히 광을 차단하여 암상태를 유지하였다. 모든 처리가 끝난 후에는 20°C 항온의 생장상(Multi-Room Incubator, WiseCube, Wonju, Korea)에 배양하면서 발아율을 조사하였다. 발아기간 동안 건조하지 않도록 증류수를 보충해 주었으며, 발아는 유근이 종피를 뚫고 1mm 이상 돌출하였을 때를 기준으로 판단하였고, 2일 간격으로 발아율을 조사하였다. 조사 시 부패한 종자들은 발아율 계산에서 제외하였다.
휴면타파 처리에 따른 종자의 발아 특성
후숙처리(after-ripening): 후숙처리는 채종한 종자를 1.5ml micro tube(Axygen, Union City, USA)에 넣고 상온(20-25°C)에 0, 3, 6, 9주 동안 건조 저장하였다. 처리 기간이 끝나면 종자를 소독한 뒤 20°C 항온의 생장상에서 각 처리 별로 명조건과 암조건으로 나누어 32일 동안 배양하였다. 즉, 암조건에서 후숙처리한 종자들은 암조건에서, 명조건에서 후숙처리한 종자들은 명조건에서 배양하였다. 저온층적처리와 GA3처리 실험도 동일하게 진행되었다.
저온층적처리(cold stratification): 소독한 종자는 90×20mm petri dish에 여과지 2매를 깔고 여과지가 마르지 않을 정도의 증류수를 주입한 후 5°C의 저온저장고에서 0, 3, 6, 9주간 처리하였다. 저온저장 기간 동안 명조건과 암조건으로 나누어 저온층적처리를 하였으며, 명조건에서 층적처리한 종자는 명조건에, 암조건에서 층적처리한 종자는 암조건에 배양을 하였다. 각각의 처리기간 후 20°C 항온 생장상에서 32일간 배양한 뒤 발아율을 계산하였다.
GA3 처리: 정선한 종자를 0(대조구), 10, 100, 1000mg·L-1의 GA3용액을 이용하여 1.5mL brown micro tube(Watson, Kobe, Japan)에서 24시간 동안 침지 처리하였다. 처리한 종자를 재차 소독한 뒤 30립 5반복으로 치상하였고, 명조건과 암조건으로 나누어 20°C 생장상에서 32일간 배양하였다.
통계 분석
후숙처리, 저온층적처리, 그리고 GA3 처리와 광조건에 대하여 각각의 수집된 자료는 SAS 9.4(SAS Institute Inc., Cary, USA)을 이용하여 분산분석(ANOVA)을 실시하였으며, 처리평균간 통계적인 유의차 비교는 Duncans multiple range test (p = 0.05)를 사용하였다. 그래프는 Sigma Plot 10.0(Systat Software Inc., California, USA)을 이용하여 작성하였다.
결과 및 고찰
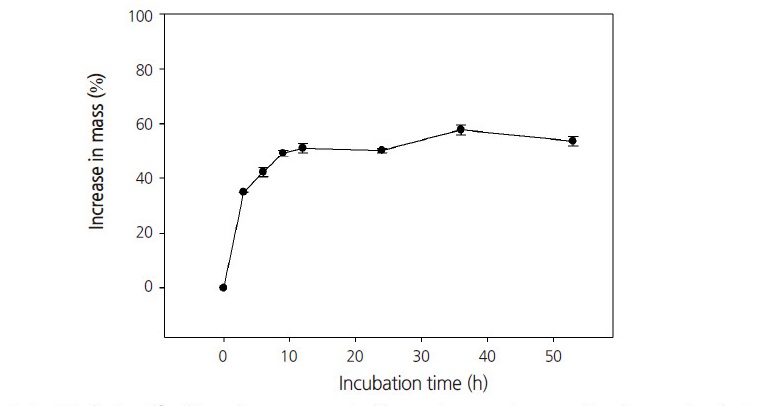
제비동자꽃 종자의 불투수성을 확인한 결과 배양한 후 24시간만에 초기 건조상태 무게보다 50% 이상 무게가 증가하였다(Fig. 1). 종피나 과피의 불투수성으로 인하여 야기된 휴면은 PY로 분류하는데(Baskin et al., 2000), 종자 초기 무게의 20% 이상 수분을 흡수하면 투수성을 갖는다고 판단한다(Baskin and Baskin, 2003). 따라서 제비동자꽃 종자는 PY가 없다고 판단된다. 그러나 실험실 내(20-25°C)에서 30일간 배양하였을 때 전혀 발아하지 않았다(data not shown).
제비동자꽃 종자의 외부형태를 관찰하였을 때 약 1mm 크기의 종자로 미세한 돌기가 종피를 둘러싸고 있음을 확인하였다(Fig. 2C). 내부형태를 관찰한 결과 완전히 자란 형태의 배와 배유 조직이 구분되어 있으며(Fig. 2D), 추가적인 배의 신장 없이 유근이 돌출하는 것을 확인할 수 있었다(Fig. 2E). Baskin and Baskin(1998)은 종자가 탈리되는 시점에 배가 완전히 자라 있으나 발아가 30일 이상 지연될 경우 이를 PD 유형으로 분류하였으며, 일반적으로 온대 낙엽수림의 초본류 중 약 60% 정도가 이러한 휴면을 갖는다고 보고하였다. 이러한 휴면 유형은 주로 비름과(Amaranthaceae), 국화과(Asteraceae), 지치과(Boraginaceae), 십자화과(Brassicaceae), 석죽과(Caryophyllaceae), 명아주과(Chenopodiaceae), 사초과(Cyperaceae), 대극과(Euphorbiaceae), 꿀풀과(Lamiaceae), 벼과(Poaceae), 장미과(Rosaceae), 그리고 현삼과(Scrophulariaceae)에서 나타난다(Baskin and Baskin, 1998).

휴면을 타파하고 발아를 촉진시키기 위해 후숙, 저온층적, GA3 처리를 수행하였다. 실험결과는 각 처리에 따라 최종 발아율과 발아소요일수가 다르게 나타나는 경향을 보였다(Table 1). 제비동자꽃 종자를 상온(20-25°C)에서 0, 3, 6, 9주 동안 후숙처리한 후 petri dish에 파종하여 20°C의 항온 조건에서 32일간 배양하면서 발아율을 조사하였다. 그 결과 광의 유무와 관계없이 0, 3, 6주 후숙처리구에서 발아가 거의 이루어지지 않았으며, 9주 후숙처리구에서 다른 처리구보다 발아율이 증가하는 것을 확인하였다(Fig. 3). 3주 후숙처리구의 최종발아율은 명조건과 암조건에서 각각 0.7%, 1.9%로 나타났다. 6주 후숙처리구의 경우 명조건에서 6일만에 발아하였으나 발아율은 2.3%에서 더 이상 증가하지 않았으며, 암조건에서는 명조건과 비슷하게 5.3%의 저조한 발아율을 보였다. 각각의 처리구를 지속적으로 관찰하였으나 더 이상 발아율이 증가하지 않았다. 반면에 9주 후숙처리구에서 명조건과 암조건의 평균발아소요일수가 10일로 3주와 6주 처리구에 비하여 빠르게 발아가 진행되었고, 명조건에서 33.2%, 암조건에서 22.6% 상대적으로 높은 발아율을 나타냈다. Geneve(2003)에 의하면 PD는 온대지역에서 가장 일반적인 휴면 유형으로 광조건, 저온층적처리 또는 후숙(건조저장)처리에 의해 휴면을 타파 할 수 있다고 하였다. 제비동자꽃의 경우 후숙처리 기간이 길어짐에 따라 발아율이 증가하는 경향을 보였다. 다만 최종 발아율이 50% 미만이었던 점을 감안하면 9주 이상의 긴 기간 처리에 대한 효과를 검증할 필요가 있다고 판단된다.
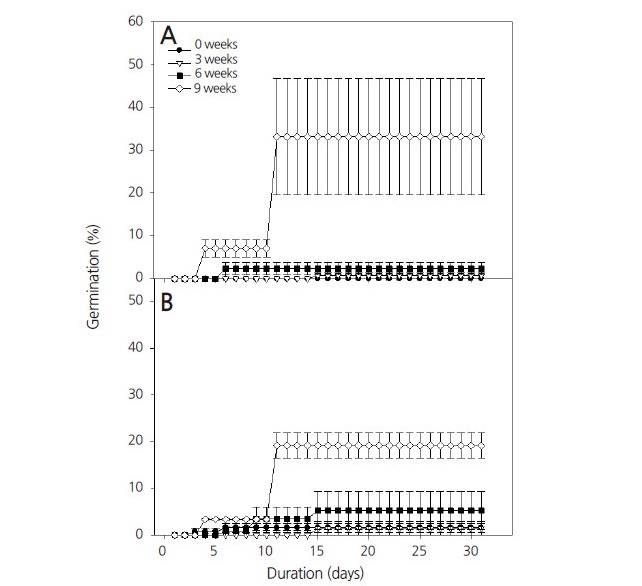
PD의 타파 방법으로 저온층적처리 실험을 수행한 결과 대조구의 발아율은 광조건에 관계없이 2% 미만이었으나, 3주간 저온층적처리 하였을 때 최종 발아율이 명조건에서 80.5%, 암조건에서 77.8%로 대조구 대비 명조건은 약 40배, 암조건은 약 37배 이상 향상되었다(Fig. 4). 6주간 저온층적처리를 하였을 때 최종발아율은 각각 명조건에서 77.0%, 암조건에서 58.4%로 나타났다. 그러나 9주 처리 시 명조건과 암조건 각각 83.3, 83.1%까지 향상되었다. 3주간 저온층적처리 후 배양하였을 때 명조건과 암조건에서 17일만에 발아하였지만, 9주 저온층적처리 시 명조건과 암조건에서 각각 5일과 3일로 처리기간이 길어질수록 발아소요일수를 단축시킬 수 있었다. 또, 저온층적처리 시 광조건의 경우 최종발아율에서는 유의적 차이가 없었다. 일반적으로 대부분의 식물들의 경우 저온처리를 통하여 종자발아가 촉진되며, 1-10°C(Bewley and Black, 1994) 범위 내의 저온층적처리는 대부분의 식물 종자의 휴면을 타파할 수 있는 가장 중요한 방법으로 알려져 있다(Baskin and Baskin, 1998). Lee et al.(2006)은 일반적으로 아무 처리를 하지 않은 개맥문동(Liriope spicata) 종자는 4.6%의 낮은 발아율을 나타내었지만 저온층적처리 이후 파종 30일만에 75.9%의 발아율을 나타내었다고 보고하였다. 또 갓 성숙한 섬현삼(Scrophularia takesimensis) 종자의 발아율은 매우 저조하지만, 5°C에서 저온층적처리 후 45일만에 66.5%의 발아율을 보인다는 연구결과도 있다(Ahn et al., 2006). 따라서 제비동자꽃 종자는 저온 6주 이상 처리 후 명조건에서 배양이 가장 적합하다고 판단된다.
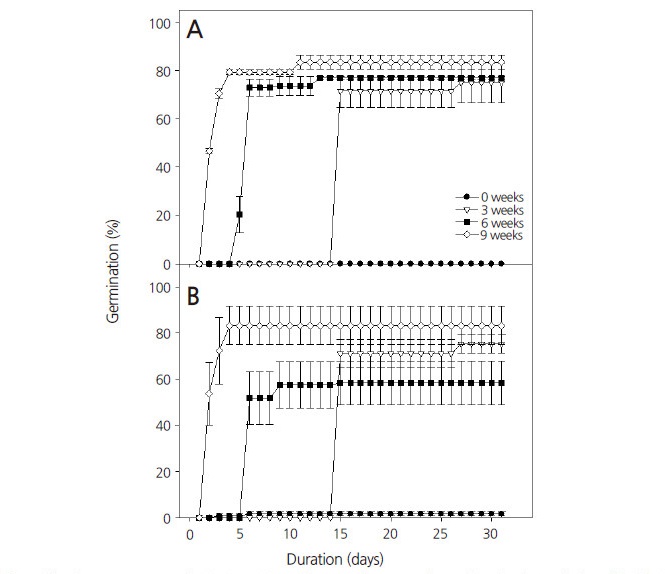
PD를 가지고 있는 종자들은 대부분 ABA(abscisic acid)와 같은 억제물질과 GA와 같은 생장촉진물질의 균형에 의해 발아가 조절된다(Çetinbaş and Koyuncu, 2006). Mehana et al.(1985)에 따르면 휴면을 갖는 종자의 내부에 GA 농도가 높으면, ABA의 비율이 낮아진다고 하였다. GA는 주로 식물의 생장을 조절하고 줄기의 신장, 개화 등 다양한 발달 과정에 영향을 미치며, 종자의 발아를 촉진하고 휴면을 타파한다(Baskin and Baskin, 1998). 이 연구에서는 제비동자꽃 종자의 휴면을 타파하고 발아를 촉진하기 위하여 0, 10, 100, 1000mg·L-1 농도의 GA3용액에 24시간 침지한 후 20°C의 생장상에서 32일 동안 발아율을 조사하였다. 대조구를 포함하여 10, 100mg·L-1 GA3처리구에서 각각 광조건에 관계없이 2% 미만의 낮은 발아율을 보였으나, 1000mg·L-1 GA3처리구에서는 명조건에서 56.3%, 암조건에서 73.2%로 발아율이 크게 증가하였다(Fig. 5). 평균발아소요일수는 명조건에서 5일, 암조건에서 3일로 발아를 시작한 후 일주일 이내에 최대 발아율에 이를 정도로 발아 속도가 빨라졌다. 1000mg·L-1 GA3처리구에서 명조건보다 암조건에서 높은 발아율을 나타내었으며 평균발아소요일수도 감소하는 경향을 보였다. GA는 대부분 종자들의 발아율을 향상시키며, 특히 미성숙배를 갖는 종자의 휴면을 타파하기 위해 주로 사용된다(Çetinbaş and Koyuncu, 2006). Bewley and Black(1994)는 종에 따라 종자의 휴면을 타파하기 위한 GA의 적정 농도가 각각 다르다고 하였으며, 딸기나무(Arbutus unedo L.)의 경우 300mg·L-1 GA3처리 시 광의 유무와 관계없이 휴면을 타파하였으나(Tilki, 2004), 눈잣나무[Pinus pumila (Pall.) Regel]의 경우 GA3를 0, 100, 500, 1000mg·L-1 농도로 처리하였을 때, 각각 48.0, 79.0, 75.7, 65.8%의 발아율을 나타내었다(Lim et al., 2015).
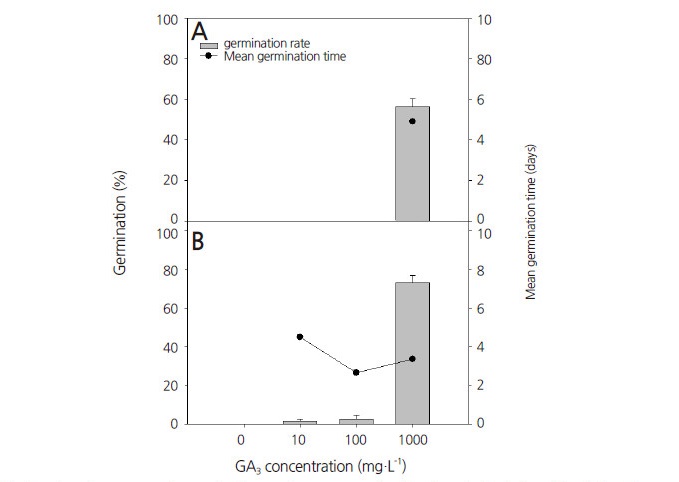
Fig. 5.
Effect of GA3 treatment on percent germination and mean germination time in Lychnis wilfordii. Seeds were incubated at 20 ± 1°C in light (16/8 h) (A) or dark conditions (B) durforing 32 d after soaking the seeds in 0, 10, 100, or 1000 mg·L-1 GA3 solution for 24 h. The control was soaked in distilled water for 24 h. Error bars indicate the mean ± SE of five replicates (n=30).
PD는 휴면의 깊이에 따라 세 가지(non-deep, intermediate, deep PD)로 나눌 수 있다(Baskin and Baskin, 2014). 이러한 휴면의 깊이는 저온처리, 후숙처리 및 GA처리에 대한 휴면타파 및 발아촉진 반응을 통하여 분류할 수 있다. 보통 non-deep PD를 보이는 종자들은 GA처리를 통하여 휴면이 타파되고, intermediate PD는 휴면이 타파되는 종들과 타파되지 않는 종들이 다양하게 존재한다. 그러나 deep PD를 보이는 종자들은 GA처리로 휴면이 타파되지 않는다. 뿐만 아니라, non-deep PD의 경우 1-2개월 정도의 저온층적처리를 통해 휴면이 완전히 타파되는 경우가 대부분이다(Baskin and Baskin, 2004). 본 연구에서, 제비동자꽃 종자는 1000mg·L-1 GA 처리, 3주 이상의 저온처리 등을 통하여 휴면이 완전히 타파되었다. 따라서 제비동자꽃 종자의 휴면유형은 non-deep PD로 분류할 수 있었다.
결론적으로 한반도에 자생하는 제비동자꽃의 종자는 모체에서 탈리되는 시점에서 성숙배를 가지고 있고, 대부분의 종자는 30일 내 전혀 발아하지 않는 생리적 휴면(PD)을 가지고 있다. 제비동자꽃 종자의 수분흡수 실험을 통하여 수분흡수에 장애가 없다는 것을 알 수 있었고, 이를 통하여 물리적 휴면(PY)은 없다는 결론을 내릴 수 있다. 또한 저온(cold stratification), 후숙(after-ripening) 및 GA처리에 의해 휴면이 타파되며, 이상의 결과를 바탕으로 제비동자꽃 종자는 non-deep PD로 분류할 수 있다. 본 연구를 통하여 제비동자꽃 종자의 휴면에 대한 학술적인 결론을 얻을 수 있었으며, 이러한 결과는 상업적인 활용을 위한 대량증식법에 매우 유용한 자료가 될 것이다.
