

서 언
감초(Glycyrrhiza spp.)는 콩과(Leguminosae)에 속하는 다년생 초본식물로, 아시아와 유럽의 건조지대를 중심으로 널리 자생하고 있는 약용식물이다. 감초의 뿌리 및 지하경에는 글리시리진(Glycyrrhizin)을 포함한 사포닌, 플라보노이드, 쿠마린류 등 다양한 생리활성물질이 풍부하여(Zeng et al. 1990; Zang 2020; Wu et al. 2022), 항염, 항바이러스, 항알러지, 간 보호, 면역 조절 등의 약리효과가 보고되어 있다(Asl and Hosseinzadeh 2008; Fiore et al. 2008). 감초는 한방에서 가장 자주 처방되는 핵심 약재로 오랫동안 이용되어 왔으며(MOHW 2001; Wang et al. 2013), 최근에는 건강기능식품, 화장품, 식품첨가물 등의 원료 소재로도 각광 받고 있다(Yan et al. 2019).
대한민국약전에는 만주감초(G. uralensis), 유럽감초(G. glabra), 창과감초(G. inflata), 코르신스키(G. korshinskyi) 등 네 종이 약용 감초로 등재되어 있으며(MFDS 2025), 이 중 산업적 중요성이 높은 종은 만주감초와 코르신스키 감초이다. 만주감초는 전통 한의학에서 가장 먼저 사용되어 왔으며, 동북아시아 각국 공정서에서도 기원(起源)식물로 널리 인정받고 있어(Choi 2015) 높은 가격으로 유통되고 있는 감초이다. 코르신스키는 만주감초와 유럽감초의 자연교잡으로 유래된 종으로 추정되는데(Hantemirova et al. 2020; Kim et al. 2025), 강우량이 많고 사계절이 뚜렷한 한국의 기후에 만주감초보다 비교적 잘 적응하는 특성이 있다(Park et al. 2014; Lee et al. 2020). 최근에는 수량 및 품질이 우수한 ‘원감’ 품종이 개발되어, 기존 만주감초를 대체할 국산 감초로 주목받고 있다(Kim et al. 2020).
감초는 오랫동안 중국, 몽골, 중앙아시아 등지에서 야생 채취 방식으로 생산되어 왔으나, 자원 고갈로 인해 최근 수십 년간 재배 중심으로 급속히 전환되고 있다(Hayashi and Sudo 2009; Huang et al. 2010; Li et al. 2013; Alamgir 2017; Jia et al. 2023). 그러나 현재까지도 재배 감초로부터 종자를 지속적으로 생산ㆍ공급하는 시스템이 마련되지 않아 완전한 품종 재배 체제는 갖추지 못하고 있다. 이는 품종개발의 부재 때문이 아니라 감초의 번식 방식 및 생식 특성이 품종 활용을 어렵게 하기 때문이다.
감초는 지하경과 종자 두 가지 방식으로 번식이 가능하다. 지하경을 이용하여 재배할 경우 초기 생육이 빠르고 유전적 균일성이 높아 품질 표준화에 유리한 장점이 있으나, 수확, 보관, 이식 과정에서 부패가 심하여 관리가 어렵고 종자에 비해 증식률도 낮아 대량 보급에는 제한이 따른다. 반면 종자는 관리의 편리성과 작업 효율성이 높아 농업인들이 선호하지만, 충분한 종자량 확보까지 장기간 재배해야 하는 어려움이 있다(GWARES 2013). 이로인해 중국 등 주요 산지 국가들에서도 실제 재배 시에는 뿌리 생산에 집중하고, 종자는 여전히 야생 개체에서 채취하고 있어 품종 순환이 이루어지지 않고 있다. 따라서 품종 재배 체제로의 전환을 위해서는 재배 과정에서 안정적으로 종자를 확보할 수 있는 기술이 필요하며, 이를 위해 장기 재배 조건에서의 생식 생장 특성에 대한 명확한 이해가 요구된다.
종자 증식을 위해 감초를 장기 재배하게 되면 약재 생산과 관련된 변화들도 새롭게 검토하여 그에 맞는 자원화 전략이 필요하다. 대한민국약전에서는 감초의 이용 부위를 “뿌리 및 뿌리줄기(지하경)로서 그대로 또는 주피를 제거한 것”으로 규정하고 있다(MFDS 2014). 그런데 한국과 중국의 재배 현장에서는 저연령의 감초의 지하경은 충분히 비대하지 않아 상품 가치가 낮고, 뿌리 비대만 방해한다(Park et al. 2002; Park et al. 2003; Wani et al. 2022; Jia et al. 2023)는 이유로 대부분 생육 초기에 제거된다. 이에 유통되는 감초는 사실상 뿌리에 한정되고 있어, 실제 섭취 부위도 과거 야생 감초의 ‘뿌리+지하경’에서 재배 감초의 ‘뿌리’ 중심으로 변화하고 있다. 이러한 지하부 조성비의 변화는 약효 구성이나 품질 특성에도 잠재적인 영향을 미칠 소지가 있지만, 해당 조성비를 실험적으로 정량화하여 보고한 연구는 아직까지 전무하다. 또한 장기 재배 시에는 지하경의 비중이 야생에 가깝게 증가할 수 있어 약재로의 이용 가능성도 재고할 여지가 있다. 한편 감초는 지하경을 통해 유전적으로 동일하면서도 잠재적으로 독립된 자손인 영양개체(ramets)를 생산할 수 있다. 온도 등 환경 조건에 따른 개화, 결실 및 지하경과 뿌리 발달 등은 각 영양개체를 중심으로 상호 영향을 미치므로, 관련 특성에 대한 이해는 재배 및 품종육성 전략을 세우는데 중요한 기초자료가 될 수 있다.
이러한 배경에도 불구하고, 감초에 대한 기존 연구는 대부분 유효성분 및 약리효과 분석에 국한되어 있으며, 장기 재배 및 온도와 같은 환경 요인이 감초의 영양 및 생식 생장에 미치는 영향을 통합적으로 다룬 연구 사례는 매우 드물다. 이에 본 연구에서는 산업적으로 중요한 감초 두 종(G. uralensis, G. korshinskyi)을 대상으로, 특별히 설계된 온도구배터널 내에서 5년간 재배하며 온도변화에 따른 지상부 생육, 개화·결실, 그리고 뿌리-지하경 구조 발달 양상을 종합적으로 분석하였다. 이를 통해 감초의 안정적인 종자생산 체계 구축 및 품종 활용, 나아가 재배 조건 최적화를 위한 생리학적 기초자료로서의 활용 가능성을 모색하고자 한다.
재료 및 방법
재배장소 및 온도처리
본 연구는 충청북도 음성에 위치한 국립원예특작과학원 인삼특작부 시험포장(36°56'N, 127°45'E) 내 설치된 온도구배터널(TGT, temperature gradient tunnel system)에서 수행하였다. 해당 터널은 가로 2.6m × 높이 2.5m × 길이 27m 크기의 비닐하우스로, 구간별 온도 차이를 유도하는 반폐쇄형 복합환경제어시스템(TGC-Soldan, Soldan Crop., Seoul, Korea)을 갖추고 있다(Fig. 1). 터널 입구는 개방되어 있어 외부 기온과 유사한 환경을 유지하고, 후방으로 갈수록 공기가 가열되어 기온이 점차 상승하도록 설계되었다. 후미부에는 6개의 대형 팬이 설치되어 있어 입구에서 유입된 공기가 뒤쪽으로 자연스럽게 빠져나갈 수 있도록 흐름을 조절하고, 흐린 날이나 야간에는 온풍기를 통해 따뜻한 공기가 후방에서 전방으로 이동하며 24시간 정교한 온도구배를 형성할 수 있도록 하였다. 터널에는 입구 외부 상단(높이 2.5m)과 내부 약 1/4 지점마다 온도센서(1400-101, LI-COR Inc., Lincoln, NE, USA)가 설치되어 있으며, 센서에서 측정된 온도 데이터는 제어기로 전달되어 6개의 팬이 선별적으로 작동하여 공기 흐름을 조정하도록 구성되어 있다. 또한 각 구간의 센서로부터 수집된 온도 데이터는 데이터로거(CR 1000, Campbell Scientific Inc., Logan, UT, USA)에 1시간 간격으로 기록되었다.
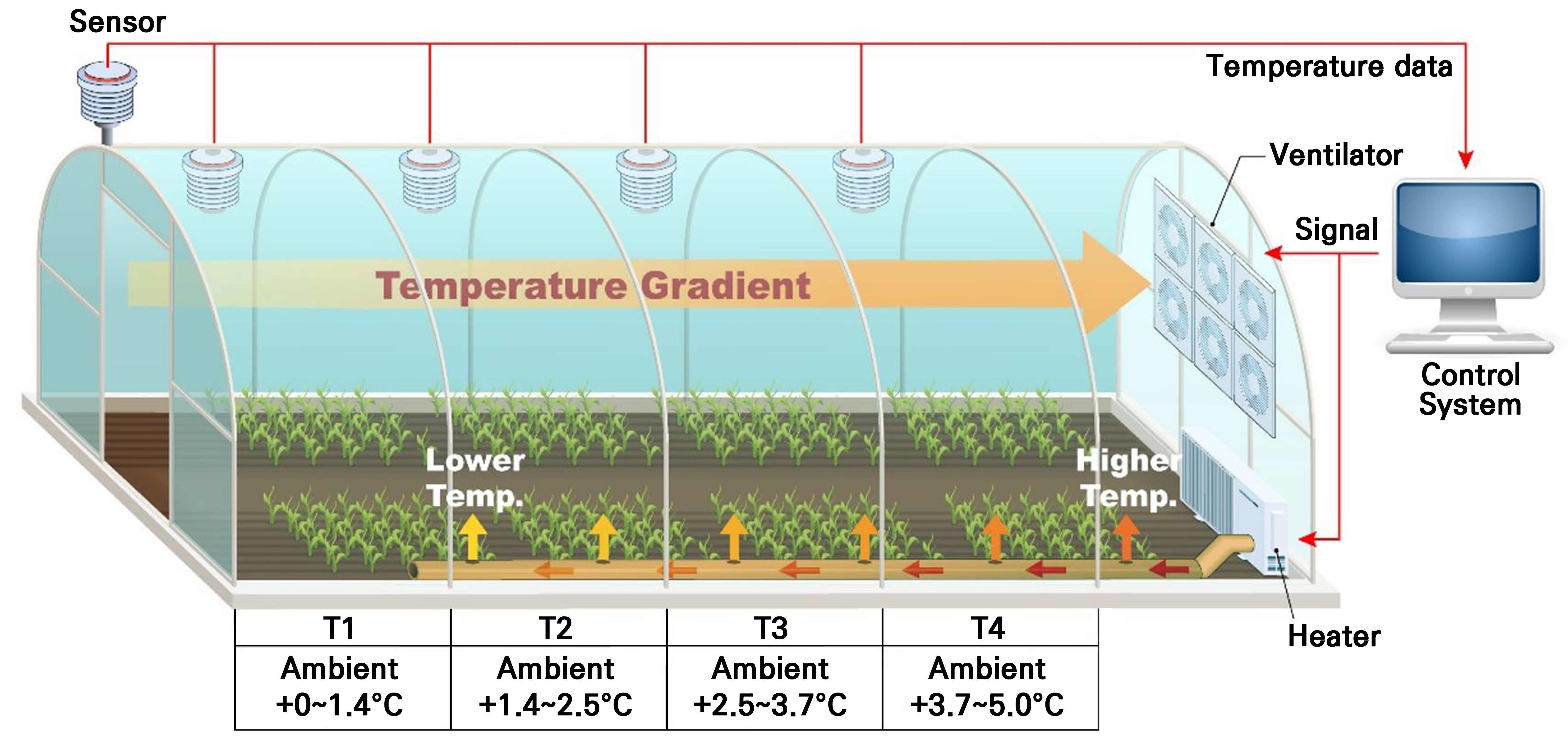
감초의 생육초기에는 안정적인 활착을 유도하고 생육 장해를 방지하기 위해, 정식일인 2017년 5월 17일부터 동년 8월 8일까지는 측창을 모두 개방하여 구간 간 온도 차이 없이 동일 조건에서 재배하였다. 이후, 일부 겨울철을 제외한 생육기를 중심으로 온도구배 장치를 24시간 연속 가동하여 온도 차이를 유지하며 5년간 재배를 지속하였다. 저온 구간와 고온구간의 온도 차이는 최대 약 5.9°C로 설정하였다.
시험재료 및 재배방법
재배지 조성은 온도구배터널 바닥 토양에 10a당 퇴비 2,000kg, 질소 17kg, 인산11kg, 칼리14kg을 시용한 후 관리기로 혼합하고, 약 35cm 높이의 두둑을 터널 양쪽에 각각 1줄씩 조성하였다. 두둑의 중앙에서 약 한 뼘 간격의 양옆에 점적 호스를 1줄씩 설치하고, 그 위를 검정색 비닐로 멀칭 하였다. 시험재료는 만주감초(G. uralensis Fisch.)와 G. korshinskyi 종인 원감(G. glabra x G. uralensis F.) 품종을 이용하였다. 만주감초는 2017년 봄 제천 감초영농조합법인에서 구입한 중국산 수입 종자로, 3월 중순 200공 트레이에 파종 후 비닐하우스에서 육묘하여 약 2개월 후에 정식에 이용하였다. 원감 품종은 국립원예특작과학원 시험포장에서 유지되고 있는 모본에서 2017년 봄 수확한 지하경을 1–2개 눈이 포함되도록 약 5cm로 절단하여 정식하였다. 재식은 두둑 중앙에 20cm 간격으로 1열씩 식재하였고, 터널 내 양쪽 두둑에 각각 만주감초와 원감 품종을 배치하였다. 정식일은 2017년 5월 17일이며, 정식 후 생육 초기 약 한 달간은 가뭄 시에만 임의로 관수하였고, 그 이후에는 별도 관수 없이 자연조건에 의존하여 재배 하였다.
생육, 개화, 결실 및 지하부 특성 조사
1년차인 2017년에는 개화가 거의 관찰되지 않아 조사에서 제외하였으며 처리구별로 초장, 줄기수, 분지수만을 조사하였다. 2년차(2018년)부터 4년차(2020년)까지는 온도 구간별로 생육특성(초장, 줄기수)과 더불어 생식특성(개화, 결실수 등)을 조사하였다. 3년차와 4년차는 모주와 신초를 연차별로 구분하여 개화 및 결실 특성을 평가하였다. 신초는 모주에서 발생한 지하경에서 이듬해 새롭게 자란 줄기로, 정식 구멍에서 발생한 줄기를 모주로, 그 외 위치에서 발생한 줄기를 신초로 간주하였다. 5년차(2021년)에는 주근 및 지하경의 길이, 굵기, 중량 등 지하부 생육 지표와 전체 지하부 생체량을 조사하였다.
통계분석
연구 결과는 SAS 프로그램(SAS Enterprise Guide 4.3, SAS Institute Inc., Cary, NC, USA)을 이용하여 분석하였으며, 데이터는 평균 ± 표준편차(means ± SD)로 제시하였다. 각 온도처리 간 생육 및 개화·결실 특성의 유의차 검정은 Duncan’s Multiple Range Test(DMRT, p < 0.05)로 수행하였다. 또한 개화수 및 결실수와 생육 지표(초장, 줄기수), 기온(5–10월 월별 및 생육기 평균기온) 간의 관계를 분석하기 위해 3–4년차 감초를 대상으로 피어슨 상관분석을 하였다.
결 과
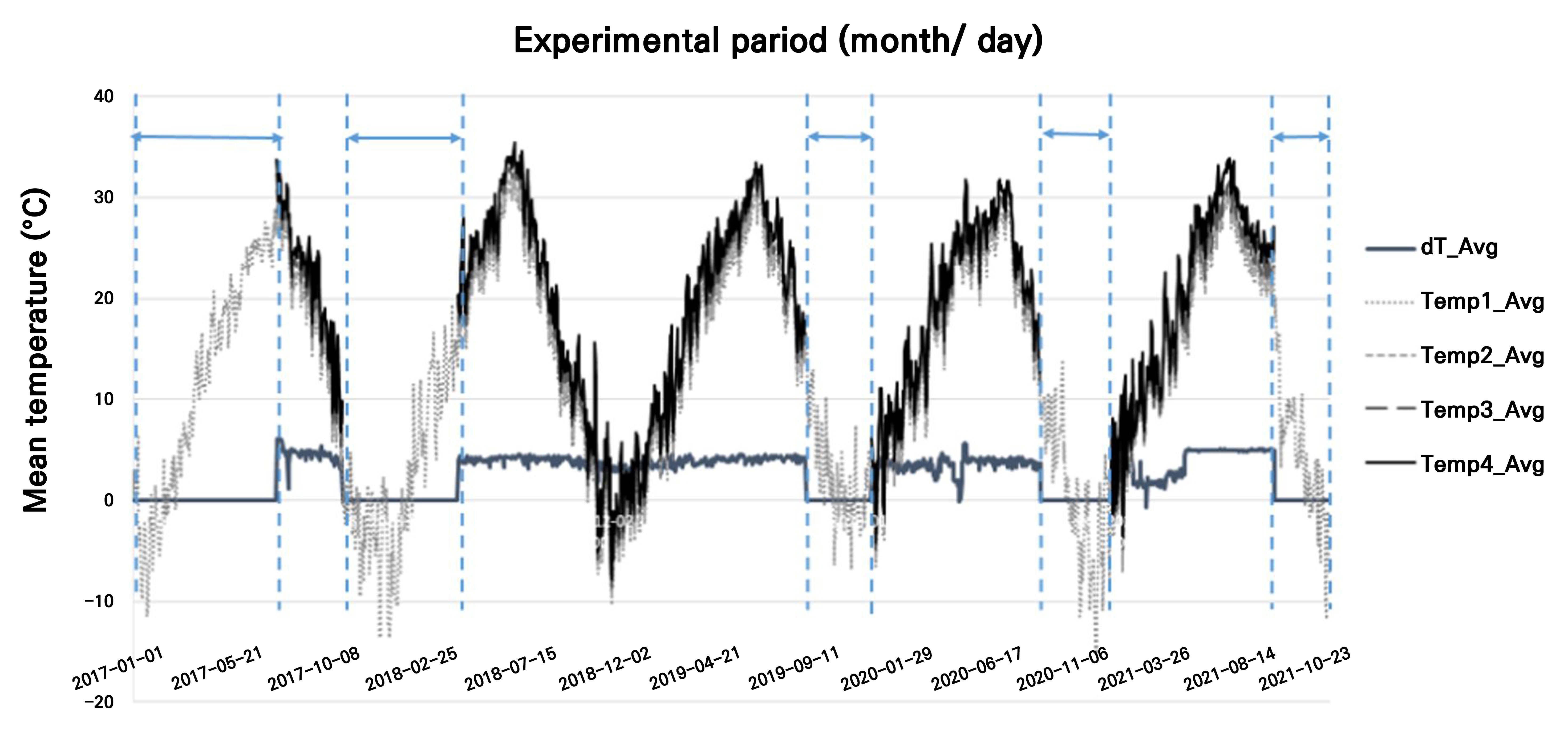
온도구배터널 내 5년간 구간별 온도변화는 Fig. 2와 같다. 외기온도와 유사한 T1 구간부터 T2, T3, T4 구간으로 갈수록 온도 높여 최대 약 4.0–5.9°C 정도 구배 차이를 유지 하였다.
1년생 감초의 종별 지상부 생육은 온도구간에 따라 뚜렷한 차이를 보였다(Table 1). 만주감초의 초장은 온도가 낮은 T1구간에서 가장 컸고 온도가 높은 T4구간으로 갈수록 작았다. 줄기수 또한 T1구간에서 가장 많았고, T4 구간에서는 가장 적었다. 원감 품종도 대체로 온도가 낮은 구간일수록 초장과 분지수가 우세하였다. 종별 비교 결과, 원감은 만주감초보다 초장이 더 컸으며, 줄기수는 다소 적었으나 분지수는 더 많았다. 1년차에는 만주감초에서 개화가 전혀 관찰되지 않았으며, 원감 품종에서는 일부 개체에서 극소수의 개화가 확인되었다.
Table 1.
Aerial growth characteristics of One-year-old licorice species in the temperature-gradient tunnel
| Species |
Temperature range sectionz | Mid-stage (Aug. 23) | Late-stage (Oct. 12) | |||||
|
Plant height (cm) |
No. of stems (stems/plant) |
No. of branches (per plant) |
Plant height (cm) |
No. of stems (stems/plant) |
No. of branches (per plant) | |||
| G. uralensis (Yr1) | T1 | 101.2 ± 16.1 a | 3.5 ± 1.6 | 39.5 ± 11.0 | 106.9 ± 17.4 a | 5.1 ± 2.1 a | 30.5 ± 4.2 a | |
| T2 | 80.1 ± 19.1 b | 2.4 ± 1.1 | 36.0 ± 8.7 | 89.3 ± 21.9 b | 3.3 ± 1.8 b | 27.0 ± 6.0 a | ||
| T3 | 67.3 ± 25.0 b | 3.3 ± 2.2 | 23.1 ± 10.7 | 77.1 ± 22.3 b | 3.7 ± 2.4 ab | 20.9 ± 6.0 b | ||
| T4 | 73.7 ± 22.4 b | 2.5 ± 1.4 | 18.3 ± 6.0 | 79.1 ± 27.4 b | 2.3 ± 1.6 b | 22.3 ± 6.6 b | ||
| Average | 80.6 ± 14.7 | 2.9 ± 0.6 | 29.2 ± 10.1 | 88.1 ± 13.6 | 3.6 ± 1.2 | 25.2 ± 4.4 | ||
| Significancey | * | ns | ns | * | * | * | ||
| Wongam (Yr1) | T1 | 119.5 ± 12.5 a | 1.6 ± 0.9 | 36.5 ± 12.2 | 122.3 ± 12.0 a | 1.9 ± 1.1 | 38.5 ± 6.6 | |
| T2 | 102.9 ± 27.7 ab | 1.1 ± 0.4 | 27.4 ± 14.0 | 111.4 ± 24.3 a | 1.6 ± 1.0 | 33.7 ± 6.6 | ||
| T3 | 96.1 ± 23.6 b | 1.1 ± 0.4 | 25.6 ± 10.4 | 100.7 ± 25.5 ab | 1.1 ± 0.4 | 32.4 ± 8.9 | ||
| T4 | 84.6 ± 17.6 b | 1.4 ± 0.5 | 27.4 ± 9.7 | 82.1 ± 36.4 b | 1.4 ± 0.5 | 29.0 ± 7.1 | ||
| Average | 100.8 ± 14.6 | 1.3 ± 0.2 | 29.2 ± 4.9 | 104.1 ± 17.1 | 1.5 ± 0.3 | 33.4 ± 3.9 | ||
| Significancey | * | ns | ns | * | ns | ns | ||
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
2년차 감초의 생육 및 생식생장 특성은 Tables 2, 3에 제시된 바와 같이 품종 및 온도구간에 따라 상이한 양상을 나타냈다. 만주 감초의 초장은 T1구간에서 141.5cm로 가장 컸고, T2 117.9cm, T3 102.3cm, T4 94.6cm 순으로 온도 상승에 따라 점진적으로 감소하였다. 줄기수는 평균 3.3개로 온도 구간 간 뚜렷한 변화 경향은 나타나지 않았다. 원감 품종의 초장은 T1구간에서 158.1cm로 가장 컸고, T4 구간에서도 135.2cm로 고온 환경에서도 생장이 크게 위축되지 않았다. 줄기수는 평균 3.2개로 만주감초와 유사하였다. 만주감초의 주당 평균 개화수는 2.2개로 매우 낮았으며, 특히 고온 구간인 T4에서는 개화가 전혀 발생하지 않았다. 결실은 T1에서 8.0%, T2에서 38.7%의 결실률을 보였으며, T3에서는 결실이 관찰되지 않았다.
Table 2.
Growth and flowering characteristics of Two-year-old licorice under temperature variations
| Temperature range sectionz |
Plant height (cm) |
No. of stems (stems/plant) | Flowers per stem |
Total flowers (flowers/plant) |
Flowering rate (%) | |
| G. uralensis (Yr2) | T1 | 141.5 ± 24.3 a | 3.4 ± 2.2 | 0.6 ± 1.0 | 3.1 ± 7.0 | 31.3 |
| T2 | 117.9 ± 27.2 b | 2.7 ± 1.4 | 0.6 ± 1.4 | 1.6 ± 3.7 | 20.0 | |
| T3 | 102.3 ± 24.3 bc | 3.7 ± 1.9 | 0.5 ± 1.5 | 3.2 ± 10.3 | 10.0 | |
| T4 | 94.6 ± 29.7 c | 3.3 ± 1.9 | 0.0 | 0.0 | 0.0 | |
| Average | 115.7 ± 30.5 | 3.3 ± 1.8 | 0.5 ± 1.2 | 2.2 ± 6.9 | 16.9 | |
| Significancey | *** | ns | ns | ns | ||
|
Wongam (Yr2) | T1 | 158.1 ± 17.7 a | 3.5 ± 2.8 | 8.5 ± 8.6 b | 38.3 ± 43.0 | 91.7 |
| T2 | 142.9 ± 20.1 ab | 3.9 ± 2.4 | 25.0 ± 12.9 a | 95.3 ± 88.2 | 100.0 | |
| T3 | 125.4 ± 16.3 c | 2.4 ± 1.7 | 17.9 ± 15.0 ab | 40.4 ± 38.4 | 92.9 | |
| T4 | 135.2 ± 2.7 bc | 2.7 ± 1.4 | 17.7 ± 10.5 ab | 44.0 ± 30.6 | 100.0 | |
| Average | 140.5 ± 21.0 | 3.2 ± 2.2 | 17.4 ± 13.5 | 56.2 ± 61.4 | 95.6 | |
| Significancey | *** | ns | * | ns |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
Table 3.
Growth and fruit set characteristics of Two-year-old licorice under temperature variation
| Temperature range sectionz | Fruits per stem | Fruits per plant |
Fruit set per flower (%) |
Fruit set rate by section (%) | |
| G. uralensis (Yr2) | T1 | 0.1 ± 0.2 | 0.3 ± 0.7 | 8.0 | 12.5 |
| T2 | 0.4 ± 1.2 | 0.6 ± 2.3 | 38.7 | 10.0 | |
| T3 | 0.0 | 0.0 | 0.0 | 0.0 | |
| T4 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Average | 0.1 ± 0.7 | 0.2 ± 1.3 | 11.7 | 5.6 | |
| Significancey | ns | ns | |||
| Wongam (Yr2) | T1 | 3.6 ± 2.5 | 14.9 ± 16.1 b | 39.0 | 75.0 |
| T2 | 12.3 ± 6.9 | 45.5 ± 38.3 a | 47.7 | 100.0 | |
| T3 | 12.4 ± 24.8 | 20.4 ± 26.8 b | 50.6 | 92.9 | |
| T4 | 3.7 ± 2.0 | 8.7 ± 4.1 b | 19.7 | 100.0 | |
| Average | 8.9 ± 14.7 | 24.6 ± 29.5 | 39.3 | 92.0 | |
| Significancey | ns | * |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
반면, 원감 품종은 주당 평균 개화수가 56.2개로 만주감초(2.2개)에 비해 약 25.5배 많았고, 결실수도 24.6개로 만주감초(0.2개)에 비해 약 123.0배 높은 수준이었다. 개화수는 T2 구간에서 95.3개로 가장 많았으며, T1·T3·T4 구간에서도 각각 38.3개, 40.4개, 44.0개로 비교적 안정적인 수준을 유지하였다. 결실률은 T1–T3구간이 38.9–50.5%로 비교적 일정하였으며, T4구간에서는 19.8%로 상대적으로 낮았다. 대체로 원감 품종은 생육, 개화, 결실 모두에서 만주감초에 비해 고온 적응도가 높았다.
재배 3년차(2019년)에는 모주(mother plant)와 지하경으로부터 새롭게 발생한 영양개체(ramets)의 신초를 구분하여 개화 및 결실 특성을 조사하였다(Tables 4 and 5). 만주감초 모주의 초장은 T1구간 144.4cm로 가장 컸으며, T2(119.3cm), T3(108.9cm), T4(94.7cm) 구간으로 갈수록 온도 상승에 따라 감소하였다. 줄기수는 온도 구간 간 큰 차이는 없었으며 평균 7.8개 수준을 유지하였다. 줄기당 화수는 3.4–5.2개 범위 였으며, T2구간에서 가장 많았다. 총 개화수는 T1(38.6개), T2(40.6개)에서 비교적 높았으나, T3(25.4개), T4(22.5개)에서는 감소하는 경향을 보였다. 결실수는 온도 증가에 따라 뚜렷한 감소세를 보였으며, 이에 따라 화당 결실률은 T1(84.0%), T2(69.9%), T3(49.8%), T4(39.4%)로 고온에서 지속적인 감소세를 나타냈다.
Table 4.
Growth and flowering characteristics of Three-year-old G. uralensis under temperature variations
|
Temperature range sectionz |
Plant height (cm) |
No. of stems (stems/plant) | Flowers per stem |
Total flowers (flowers/plant) |
Flowering rate (%) | |
| Mother shoot | T1 | 144.4 ± 20.3 a | 10.9 ± 4.2 | 3.5 ± 1.6 | 38.6 ± 24.8 | 100.0 |
| T2 | 119.3 ± 30.1 b | 7.3 ± 5.1 | 5.2 ± 3.6 | 40.6 ± 39.1 | 100.0 | |
| T3 | 108.9 ± 29.7 bc | 7.3 ± 4.5 | 4.2 ± 4.4 | 25.4 ± 32.2 | 88.0 | |
| T4 | 94.7 ± 35.8 c | 6.4 ± 6.5 | 3.4 ± 4.4 | 22.5 ± 32.8 | 85.7 | |
| Average | 116.8 ± 33.0 | 7.8 ± 5.2 | 4.3 ± 3.7 | 32.5 ± 33.7 | 93.4 | |
| Significancey | *** | ns | ns | ns | ||
| New shoot 1 | T1 | 143.7 ± 25.0 a | 9.8 ± 5.1 a | 3.6 ± 2.1 b | 32.9 ± 22.1 | 100.0 |
| T2 | 117.9 ± 35.3 b | 4.8 ± 3.2 b | 6.1 ± 4.1 b | 30.3 ± 24.6 | 95.2 | |
| T3 | 116.3 ± 26.4 b | 4.3 ± 2.5 b | 11.0 ± 9.4 a | 50.0 ± 46.6 | 84.2 | |
| T4 | 105.2 ± 29.7 b | 5.3 ± 3.8 b | 4.7 ± 3.6 b | 30.8 ± 34.6 | 84.6 | |
| Average | 120.7 ± 32.0 | 5.9 ± 4.2 | 6.7 ± 6.3 | 36.5 ± 34.0 | 91.0 | |
| Significancey | ** | *** | ** | ns |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
Table 5.
Fruit set characteristics of Three-year-old G. uralensis (mother shoot and new shoot) under temperature variations
| Temperature range sectionz | Fruits per stem | Fruits per plant |
Fruit set per flower (%) |
Fruit set rate by section (%) | |
| Mother shoot | T1 | 3.0 ± 1.4 a | 32.4 ± 22.1 a | 84.0 | 100.0 |
| T2 | 3.7 ± 2.7 a | 28.3 ± 25.9 a | 69.9 | 100.0 | |
| T3 | 2.4 ± 3.2 ab | 12.6 ± 19.1 b | 49.8 | 84.0 | |
| T4 | 1.0 ± 1.2 b | 8.9 ± 17.0 b | 39.4 | 78.6 | |
| Average | 2.7 ± 2.6 | 21.0 ± 23.5 | 60.7 | 91.0 | |
| Significancey | * | ** | |||
| New shoot 1 | T1 | 2.6 ± 1.8 ab | 24.2 ± 17.7 a | 73.5 | 100.0 |
| T2 | 3.7 ± 2.4 a | 18.6 ± 16.1 a | 61.4 | 95.2 | |
| T3 | 4.4 ± 5.3 a | 20.5 ± 26.7 a | 37.8 | 78.9 | |
| T4 | 0.7 ± 0.7 b | 4.5 ± 5.4 b | 14.5 | 69.2 | |
| Average | 3.1 ± 3.4 | 17.7 ± 19.7 | 46.8 | 85.9 | |
| Significancey | * | * |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
신초1의 경우, 초장이 T1 구간에서 143.7cm로 가장 컸으며, T2–T4 구간은 유사하여 모주와 달리 온도 상승에 따른 초장 감소가 덜 분명하게 나타났다. 개화수는 T3에서 50.0개로 가장 많았고, 줄기당 화수 및 총 개화수 모두에서 대체로 모주보다 높은 수치를 보였다. 반면 결실률은 T1 구간에서 73.5%, T2 61.4%, T3 37.8%, T4 14.5%로, 모주와 유사하게 고온 구간에서 낮아지는 경향을 나타냈다. 또한 T1–T4 구간 전체의 평균 개화수는 모주가 32.5개, 신초1이 36.5개로 신초가 다소 많았으나, 결실수는 모주 21.0개, 신초1 17.7개로 오히려 신초에서 결실이 더 적게 이루어졌다.
온도구간별 3년생 원감 품종의 생육, 개화 및 결실 특성은 Tables 6, 7에 제시된 바와 같다. 모주의 초장은 T1 구간(156.1cm)에서 가장 길었으며, T4 구간(123.6cm)까지 온도 상승에 따라 유의하게 감소하였다. 줄기당 화수는 T2에서 가장 많았으며(10.8개), T4 구간에서는 6.2개로 가장 적었다. 줄기당 화수와 개화수는 모두 T1, T2에서 높았고, 온도 상승에 따라 감소하는 경향이 뚜렷하게 나타났다. 모주의 평균 결실률은 83.0%로, 만주감초 모주의 60.7% 대비 현저히 높았으며, T4 구간에서도 70.6%로 높은 수준을 유지하였다.
Table 6.
Growth and flowering characteristics of Three-year-old wongam (mother shoot and new shoot) under temperature variations
| Temperature range sectionz |
Plant height (cm) |
No. of stems (stems/plant) | Flowers per stem |
Total flowers (flowers/plant) |
Flowering rate (%) | |
| Mother shoot | T1 | 156.1 ± 11.0 a | 9.9 ± 6.5 a | 8.1 ± 3.2 b | 71.7 ± 34.9 ab | 100.0 |
| T2 | 140.0 ± 13.1 b | 7.7 ± 4.3 a | 10.8 ± 3.0 a | 79.2 ± 38.1 a | 100.0 | |
| T3 | 135.8 ± 17.2 b | 4.3 ± 4.9 b | 7.6 ± 3.8 b | 41.8 ± 78.2 bc | 93.8 | |
| T4 | 123.6 ± 16.2 c | 2.6 ± 1.6 b | 6.2 ± 3.9 b | 12.6 ± 9.8 c | 92.9 | |
| Average | 139.2 ± 17.9 | 6.3 ± 5.3 | 8.5 ± 3.8 | 54.7 ± 53.0 | 96.7 | |
| Significancey | *** | *** | ** | *** | ||
| New shoot 1 | T1 | 154.3 ± 10.8 a | 9.1 ± 9.0 | 5.3 ± 2.0 b | 42.9 ± 37.7 b | 100.0 |
| T2 | 131.3 ± 14.5 b | 5.2 ± 4.5 | 9.2 ± 5.1 a | 44.5 ± 40.6 b | 100.0 | |
| T3 | 127.6 ± 22.4 b | 10.1 ± 6.9 | 11.1 ± 4.0 a | 107.2 ± 70.5 a | 100.0 | |
| T4 | 133.5 ± 11.5 b | 6.9 ± 5.2 | 9.4 ± 3.3 a | 61.4 ± 47.5 b | 100.0 | |
| Average | 137.1 ± 18.5 | 7.9 ± 6.8 | 8.7 ± 4.2 | 64.0 ± 56.0 | 100.0 | |
| Significancey | *** | ns | ** | ** |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
Table 7.
Fruit set characteristics of Three-year-old wongam licorice (mother shoot and new shoots) under temperature variations
| Temperature range sectionz | Fruits per stem | Fruits per plant |
Fruit set per flower (%) |
Fruit set rate by section (%) | |
| Mother shoot | T1 | 7.0 ± 2.6 b | 63.5 ± 32.8 ab | 88.5 | 100.0 |
| T2 | 9.8 ± 2.6 a | 72.8 ± 36.4 a | 91.9 | 100.0 | |
| T3 | 5.8 ± 3.3 bc | 33.9 ± 68.8 bc | 81.2 | 93.8 | |
| T4 | 4.0 ± 2.8 c | 8.9 ± 7.4c | 70.6 | 92.9 | |
| Average | 7.0 ± 3.5 | 48.1 ± 48.7 | 83.0 | 96.7 | |
| Significancey | *** | *** | |||
| New shoot 1 | T1 | 4.2 ± 2.1 | 32.3 ± 31.6 b | 75.2 | 100.0 |
| T2 | 6.6 ± 5.0 | 36.1 ± 36.1 ab | 81.0 | 92.3 | |
| T3 | 6.7 ± 3.6 | 56.7 ± 31.0 a | 52.9 | 100.0 | |
| T4 | 4.1 ± 3.2 | 20.7 ± 12.3 b | 33.7 | 100.0 | |
| Average | 5.4 ± 3.7 | 36.7 ± 31.4 | 60.7 | 98.1 | |
| Significancey | ns | * |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
신초의 경우, T2에서 T4까지 온도 상승에 따른 초장 감소가 없었으며, 개화수는 T3 구간에서 107.2개로 가장 많았다. 평균 결실률은 60.7%로, 모주(83.0%) 보다는 낮았지만, 만주감초 신초의 평균 결실률(46.8%)보다는 높았다. 특히 T4에서도 33.7%의 결실률을 유지하여, 고온 환경에서도 일정한 수준 이상의 생식 성장이 지속되었음을 시사한다. 종합하면, 원감 품종은 개화수 측면에서는 신초가 모주보다 우세하였고, 결실수는 모주에서 더 높게 나타났다. 이는 전체적인 경향에서 만주감초와 유사하지만, 고온(T4) 구간에서의 결실률 감소폭이 더 작아, 고온 장해에 상대적으로 내성이 강한 생식 특성을 보인 것으로 판단된다.
만주감초 4년생의 경우 모주 및 신초1의 초장은 온도가 높아질수록 점진적으로 감소하여 3년차와 유사한 생육 패턴을 보였다. 그러나 신초2는 T2 및 T3 구간에서 유의적으로 가장 높은 초장을 기록하여, 다른 생장 형태를 보였다(Table 8). 총화수는 모주, 신초1, 신초2 모두 T3 구간에서 가장 많았으며, 이는 생육에 최적인 온도(T1–T2)보다 개화에 적합한 온도가 상대적으로 높음을 시사한다.
Table 8.
Growth and flowering characteristics of Four-year-old G. uralensis (mother shoot and new shoots) under temperature variations
| Temperature range sectionz |
Plant height (cm) |
No. of stems (stems/plant) | Flowers per stem |
Total flowers (flowers/plant) |
Flowering rate (%) | |
| Mother shoot | T1 | 132.0 ± 24.3 a | 7.2 ± 4.0 | 4.0 ± 4.9 | 23.2 ± 17.2 b | 90.0 |
| T2 | 128.6 ± 37.8 a | 5.7 ± 5.1 | 6.4 ± 4.0 | 33.7 ± 38.1 ab | 94.3 | |
| T3 | 113.8 ± 32.9 ab | 6.1 ± 5.0 | 10.8 ± 19.3 | 49.8 ± 45.4 a | 97.2 | |
| T4 | 98.7 ± 31.1 b | 8.7 ± 10.4 | 3.9 ± 2.1 | 25.3 ± 17.0 b | 100.0 | |
| Average | 120.0 ± 34.5 | 6.5 ± 5.9 | 7.1 ± 11.9 | 36.0 ± 36.9 | 95.4 | |
| Significanty | ** | ns | ns | * | ||
| New shoot 1 | T1 | 124.2 ± 26.4 a | 10.7 ± 7.0 a | 2.5 ± 1.5 b | 27.1 ± 19.8 | 90.0 |
| T2 | 123.0 ± 32.7 a | 7.3 ± 5.1 b | 4.6 ± 2.6 a | 28.9 ± 19.6 | 97.4 | |
| T3 | 118.3 ± 38.7 a | 6.2 ± 4.6 b | 5.2 ± 2.8 a | 32.7 ± 33.6 | 97.5 | |
| T4 | 87.6 ± 27.0 b | 4.7 ± 2.8 b | 4.3 ± 3.0 a | 14.9 ± 9.9 | 100.0 | |
| Average | 116.6 ± 35.0 | 7.1 ± 5.3 | 4.4 ± 2.7 | 27.9 ± 25.0 | 96.2 | |
| Significanty | ** | ** | ** | ns | ||
| New shoot 2 | T1 | 107.8 ± 31.7 b | 2.5 ± 2.0 b | 3.7 ± 4.8 b | 7.2 ± 8.0 c | 50.0 |
| T2 | 147.6 ± 35.6 a | 3.6 ± 2.8 ab | 8.1 ± 4.2 a | 26.1 ± 22.3 ab | 92.3 | |
| T3 | 142.2 ± 29.8 a | 5.1 ± 3.2 a | 9.3 ± 4.7 a | 40.4 ± 21.0 a | 100.0 | |
| T4 | 114.1 ± 21.4 b | 2.2 ± 1.6 b | 10.0 ± 4.9 a | 18.5 ± 12.0 bc | 100.0 | |
| Average | 134.9 ± 34.4 | 3.5 ± 2.8 | 8.4 ± 4.8 | 25.9 ± 21.2 | 85.6 | |
| Significanty | ** | * | * | ** |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
결실 특성은 구간별로 차이를 보였으나 전반적으로 고온 구간(T4)으로 갈수록 결실수와 결실률이 감소하는 경향을 나타냈다. 모주의 경우 T3에서 줄기당 결실수는 가장 많았으나, 화당 결실률은 26.9%로 낮았고, 신초1과 신초2 역시 고온 구간(T4)에서 결실률이 각각 27.3%, 7.1%로 급감하였다(Table 9). 특히 신초2는 해당 구간에서 전체적으로 가장 낮은 구간 결실률(59.4%)을 기록하였다. 이는 고온 스트레스가 생식생장, 특히 결실 과정에 부정적 영향을 미친 것으로 판단된다. 모주의 줄기당 결실수 평균은 3년차(2.7개)보다 4년차(3.1개)에서 약간 증가하였으나, 화당 결실률은 3년차 60.7%에서 4년차 48.3%로 감소하여, 연차에 따른 결실 품질 저하 가능성을 시사하였다. 한편, 신초1과 신초2의 결실수 및 결실률은 4년차에 감소하였으나, 개체 전체 결실수는 연차별로 증가하였기 때문에, 이는 생식기능 저하보다는 신초 수 증가에 따른 자원 분산의 결과로 해석된다.
Table 9.
Fruit set characteristics of Four-year-old G. uralensis (mother shoot and new shoots) under temperature variations
| Temperature range sectionz | Fruits per stem | Fruits per plant |
Fruit set per flower (%) |
Fruit set rate by section (%) | |
| Mother shoot | T1 | 2.1 ± 1.9 | 15.6 ± 13.9 | 67.2 | 80.0 |
| T2 | 3.3 ± 2.7 | 20.9 ± 31.8 | 61.8 | 77.1 | |
| T3 | 4.1 ± 10.8 | 13.4 ± 14.7 | 26.9 | 83.3 | |
| T4 | 1.5 ± 1.4 | 9.5 ± 10.5 | 37.4 | 80.0 | |
| Average | 3.1 ± 6.5 | 15.7 ± 21.6 | 48.3 | 80.1 | |
| Significanty | ns | ns | |||
| New shoot 1 | T1 | 1.3 ± 1.4 | 13.8 ± 17.3 | 50.9 | 80.0 |
| T2 | 1.7 ± 1.8 | 9.7 ± 9.1 | 33.4 | 89.7 | |
| T3 | 2.0 ± 2.1 | 10.8 ± 12.4 | 33.2 | 87.5 | |
| T4 | 0.9 ± 1.9 | 4.1 ± 7.1 | 27.3 | 43.8 | |
| Average | 1.6 ± 1.9 | 10.0 ± 12.0 | 36.2 | 75.2 | |
| Significanty | ns | ns | |||
| New shoot 2 | T1 | 1.2 ± 2.4 ab | 1.7 ± 2.7 a | 23.3 | 33.3 |
| T2 | 2.8 ± 2.5 a | 12.2 ± 15.3 a | 46.6 | 80.8 | |
| T3 | 2.2 ± 1.9 a | 12.1 ± 12.5 a | 30.0 | 92.9 | |
| T4 | 0.3 ± 0.8 b | 1.3 ± 3.8 a | 7.1 | 30.8 | |
| Average | 2.0 ± 2.3 | 8.7 ± 12.8 | 26.7 | 59.4 | |
| Significanty | ** | * |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
4년차 원감 품종의 초장은 모주보다 신초가 우세하였고, 개화 및 결실 능력은 신초1이 모주를 상회하였으며, 신초2는 상대적으로 낮은 결과를 보였다(Tables 10 and 11). 전체적으로 볼 때, 초장이나 개화수의 경우는 감초종이나 연령, 줄기의 유형에 따라 다소 차이를 보였으나 결실률의 경우는 모든 연령대와 줄기에서 온도가 상승할수록 뚜렷이 감소하였다.
Table 10.
Growth and flowering characteristics of Four-year-old wongam (mother shoot and new shoots) under temperature variations
| Temperature range sectionz |
Plant height (cm) |
No. of stems (stems/plant) | Flowers per stem |
Total flowers (flowers/plant) |
Flowering rate (%) | |
| Mother shoot | T1 | 124.4 ± 45.5 | 4.8 ± 4.0 | 4.4 ± 3.2 | 21.0 ± 24.4 | 100.0 |
| T2 | 135.2 ± 23.8 | 3.1 ± 2.9 | 12.0 ± 21.3 | 29.7 ± 31.8 | 93.5 | |
| T3 | 134.3 ± 25.5 | 3.1 ± 3.6 | 7.4 ± 2.7 | 25.3 ± 39.2 | 96.2 | |
| T4 | 127.0 ± 20.8 | 3.3 ± 2.1 | 6.9 ± 2.6 | 25.5 ± 19.6 | 100.0 | |
| Average | 132.4 ± 28.7 | 3.4 ± 3.3 | 8.7 ± 13.9 | 26.4 ± 32.4 | 97.4 | |
| Significanty | ns | ns | ns | ns | ||
| New shoot 1 | T1 | 140.3 ± 27.6 | 12.5 ± 8.1 a | 4.3 ± 2.9 b | 50.3 ± 37.2 | 91.7 |
| T2 | 137.8 ± 27.0 | 7.9 ± 5.1 b | 7.9 ± 2.7 a | 61.9 ± 48.2 | 100.0 | |
| T3 | 142.1 ± 21.3 | 7.6 ± 4.4 b | 7.8 ± 2.4 a | 59.6 ± 37.1 | 100.0 | |
| T4 | 137.9 ± 27.9 | 6.6 ± 3.6 b | 8.6 ± 3.0 a | 53.6 ± 27.9 | 100.0 | |
| Average | 139.7 ± 25.2 | 8.5 ± 5.8 | 7.3 ± 3.0 | 57.9 ± 40.3 | 97.9 | |
| Significanty | ns | ** | *** | ns | ||
| New shoot 2 | T1 | 151.6 ± 17.8 ab | 4.7 ± 2.7 a | 5.5 ± 1.9 | 23.6 ± 17.7 | 100.0 |
| T2 | 127.0 ± 19.6 c | 2.1 ± 2.8 c | 7.5 ± 4.5 | 15.3 ± 20.3 | 85.0 | |
| T3 | 135.6 ± 29.6 bc | 2.5 ± 1.9 bc | 7.7 ± 3.3 | 15.9 ± 10.2 | 100.0 | |
| T4 | 160.7 ± 10.1 a | 4.3 ± 2.4 ab | 7.4 ± 2.2 | 30.1 ± 19.2 | 100.0 | |
| Average | 139.9 ± 24.6 | 3.1 ± 2.7 | 7.0 ± 3.4 | 19.3 ± 17.4 | 96.3 | |
| Significanty | ** | ** | ns | ns |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
Table 11.
Fruit set characteristics of Four-year-old wongam (mother shoot and new shoots) under temperature variations
| Temperature range sectionz | Fruits per stem | Fruits per plant |
Fruit set per flower (%) |
Fruit set rate by section (%) | |
| Mother shoot | T1 | 2.9 ± 2.0 | 14.2 ± 17.3 | 67.4 | 92.3 |
| T2 | 5.1 ± 7.7 | 14.4 ± 19.9 | 48.5 | 77.4 | |
| T3 | 2.4 ± 2.2 | 10.3 ± 23.5 | 40.7 | 80.8 | |
| T4 | 1.3 ± 1.9 | 5.2 ± 7.3 | 20.3 | 50.0 | |
| Average | 3.5 ± 5.3 | 12.2 ± 20.1 | 44.2 | 75.1 | |
| Significanty | ns | ns | |||
| New shoot 1 | T1 | 2.9 ± 2.2 | 35.5 ± 29.2 a | 70.9 | 91.7 |
| T2 | 3.2 ± 2.3 | 23.5 ± 18.5 b | 37.9 | 90.9 | |
| T3 | 2.1 ± 1.5 | 14.9 ± 11.7 b | 25.6 | 90.2 | |
| T4 | 3.3 ± 2.0 | 18.6 ± 11.2 b | 34.7 | 100.0 | |
| Average | 2.8 ± 2.1 | 22.3 ± 19.8 | 42.2 | 93.2 | |
| Significanty | ns | *** | |||
| New shoot 2 | T1 | 1.8 ± 2.1 | 12.0 ± 16.6 a | 50.9 | 75.0 |
| T2 | 2.8 ± 4.1 | 4.7 ± 6.9 a | 30.4 | 75.0 | |
| T3 | 2.1 ± 3.5 | 3.5 ± 4.4 a | 22.0 | 50.0 | |
| T4 | 3.5 ± 3.6 | 11.4 ± 8.0 a | 37.9 | 85.7 | |
| Average | 2.4 ± 3.4 | 7.0 ± 10.5 | 35.3 | 71.4 | |
| Significanty | ns | * |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
연생별 개화 및 결실수는 Table 12와 같은데, 만주감초의 주당 평균 개화수는 2년생 2.2개에서 3년생 62.0개, 4년생 65.8개로 증가하였으며, 결실수는 연차별로 각각 0.2개, 35.2개, 25.6개로 3년차에 종자 수확 기대치가 최대가 되었다. 반면 원감 품종의 경우 주당 평균 개화수가 2년생에 56.2개, 3년생 104.2개, 4년생 79.7개였으며, 결실수는 각각 24.6개, 75.9개, 31.9개로 3년차 이후 생산성 증가폭이 크게 둔화되었다. 또한 원감 품종은 만주 감초보다 빠른 2년차부터 조기 결실이 가능하였으나 4년차 총생산량은 만주감초와 유사한 수준으로 떨어졌다. 결론적으로 만주감초의 종자수확 최적기는 3–4년차로, 원감 품종은 2년차부터 수확 가능성이 있지만, 생산성 측면에서는 3년차가 적기로 판단된다.
Table 12.
Average number of flowers and fruits per plant in G. uralensis and wongam by year of growth
| Category | Flower and fruit counts (per plant) by plant age | |||
| Two-year-old | Three-year-old | Four-year-old | ||
| No. of flowers | G. uralensis | 2.2 ± 6.9 b | 62.0 ± 49.9 b | 65.8 ± 44.5 |
| Wongam | 56.2 ± 61.4 a | 104.2 ± 64.1 a | 79.7 ± 51.5 | |
| Significantz | *** | *** | ns | |
| No. of fruits | G. uralensis | 0.2 ± 1.3 b | 35.2 ± 33.8 b | 25.6 ± 23.0 |
| Wongam | 24.6 ± 29.5 a | 75.9 ± 52.5 a | 31.9 ± 26.6 | |
| Significantz | *** | *** | ns | |
3–4년생 감초를 대상으로 생식 생장이 환경 스트레스, 특히 고온에 어떤 반응을 보이는지 확인하기 위하여 개화수 및 결실수와 생육지표(초장, 경수) 및 월별 기온(5–10월), 생육기 평균기온 간의 상관관계를 분석하였다(Tables 13 and 14). 초장과 개화수는 대체로 양의 상관관계가 많았다(65%). 가령 3년생 만주감초 모주의 경우 개화수와 초장 간 상관계수는 0.809, 결실수는 0.919로 나타났으며, 유사하게 4년생 원감 신초2에서도 각각 0.972 및 0.896의 높은 양의 상관계수를 보였다. 상관성이 낮은 경우는 전체의 약 30% 정도였으며, 예외적으로 음의 상관관계인 경우도 5% 정도 있었다. 이러한 결과는 감초에서 생식기관의 형성이 전체적인 생장 상태, 특히 지상부 생장과 밀접한 관련이 있음을 시사한다.
Table 13.
Correlation between Flowering/Fruiting and Growth/Temperature in Three-year-old licorice
Table 14.
Correlation between Flowering/Fruiting and Growth/Temperature in Four-year-old licorice
한편, 고온은 생식 생장 전반에 부정적인 영향을 미쳤으나, 특히 결실 단계에서 더욱 뚜렷한 억제 효과를 보였다. 대부분의 품종 및 조사구에서 월별 기온 및 생육기 평균기온과 결실수 간에는 강한 음의 상관관계가 관찰되었으며, 그 크기는 개화수에 비해 현저히 높았다. 예를 들어 3년생 만주감초 모주의 경우, 6월 평균기온과 결실수 간 상관계수는 ‒0.986, 7월은 ‒0.978, 생육기 평균기온에 대해서는 ‒0.980으로 나타나, 고온이 배 발달이나 수정 안정성 등에 부정적 영향을 주었을 것으로 추정된다(Pokharel et al. 2021). 반면 개화수의 경우 음의 상관을 보인 경우도 많았으나, 그 수치는 결실수에 비해 낮은 경향을 나타냈다. 이와 같은 결과는 감초의 생식 생장 최적화를 위해 생육기 온도 조절이 중요함을 보여주며, 특히 개화기 이후의 고온 조건은 결실 실패의 주요 요인이 될 수 있음을 시사한다.
또한 식물체 내 부위별로도 반응 양상에 차이가 있었다. 결실수의 경우 일반적으로 연령이 높은 모주와 신초1에 비해 신초2는 고온에 덜 민감한 반응을 나타냈다. 예를 들어 만주감초와 원감 품종의 4년생 모주, 신초1은 생육기 평균온도와의 상관계수가 대부분 –0.7에서 –0.9 사이로 강한 음의 상관관계를 나타냈다. 반면, 신초2의 경우, ‒0.1 수준으로 온도와의 관련성이 크게 낮아짐을 확인할 수 있다. 이러한 점은 모주가 고온 스트레스에 직접적으로 노출되더라도, 지하경의 발달과 신초의 지속적인 발생을 통해 생식 능력과 생장 기반을 유지하려는 생리적 보상 전략을 취하고 있을 가능성을 시사한다.
5년생 개체의 지하부 생육 특성은 Tables 15, 16에 요약된 바와 같이 두 품종 간 명확한 차이를 보였다. 만주감초는 온도가 상승할수록 주근장, 주근경, 생근중의 감소세가 뚜렷하였다. 반면 지하경은 온도 상승에 따라 지하경장, 지하경경, 지하경수의 감소세가 뚜렷하지 않고, 지하경중 감소폭도 주근 대비 작았다. 이는 고온 조건에서 주근 발달이 억제되더라도, 지하경을 통한 일정 수준의 영양번식 자원 확보가 가능함을 시사하며, 특히 T2–T3 구간까지 지하경 관련 지표들이 상대적으로 안정적인 수준을 유지한 점은 이러한 전략을 뒷받침한다. 이는 온도 구간별로 생육 반응이 덜한 지하경을 기반으로 신초가 발생하기 때문에, 모주(주근 기반)보다 신초의 생육이나 개화수가 온도 변화에 둔감한 생육 반응을 보이는 원인을 설명해준다. 반면 고온 적응성이 높은 원감 품종은 만주감초와 다소 상이한 생장 특성을 나타냈다(Table 16). 원감은 대부분의 지표에서 통계적으로 유의한 차이를 보이지는 않았으나, 주근 관련 지표는 구간별로 큰 변동 없이 일정한 수준을 유지하다가 T4 구간에서 감소하였다. 지하경도 대체로 유사한 특성을 보였다.
Table 15.
Underground growth characteristics of Five-year-old G. uralensisz
|
Temperature range sectionz | Main root | Rhizome | |||||||
|
Length (cm) |
Diameter (mm) | Number |
Fresh weight (g) |
Length (cm) |
Diameter (mm) | Number |
Fresh weight (g) | ||
| T1 | 73.4 ± 19.3 a | 48.9 ± 16.7 a | 5.5 ± 2.8 | 695.4 ± 382.4 a | 153.4 ± 28.0 | 15.7 ± 3.8 a | 12.6 ± 6.0 | 668.9 ± 347.3 | |
| T2 | 62.9 ± 15.2 a | 38.9 ± 12.9 ab | 5.8 ± 2.3 | 641.8 ± 466.5 ab | 133.1 ± 60.9 | 12.5 ± 4.1 ab | 10.4 ± 5.7 | 553.2 ± 501.4 | |
| T3 | 47.5 ± 11.7 b | 35.3 ± 13.3 b | 5.9 ± 2.0 | 370.9 ± 325.6 bc | 135.8 ± 45.4 | 11.4 ± 4.8 b | 10.8 ± 7.0 | 505.3 ± 378.3 | |
| T4 | 42.4 ± 8.6 b | 30.4 ± 11.4 b | 4.2 ± 1.8 | 268.9 ± 160.6 c | 144.3 ± 82.3 | 11.0 ± 2.8 b | 12.8 ± 6.7 | 293.3 ± 214.3 | |
| Average | 57.4 ± 18.6 | 38.9 ± 15.0 | 5.5 ± 2.3 | 508.2 ± 393.5 | 141.1 ± 53.0 | 12.8 ± 4.4 | 11.5 ± 6.3 | 524.8 ± 395.3 | |
| Significancey | *** | * | ns | * | ns | * | ns | ns | |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
Table 16.
Underground growth characteristics of Five-year-old wongam
| Temperature range sectionz | Main root | Rhizome | |||||||
|
Length (cm) |
Diameter (mm) | Number |
Fresh weight (g) |
Length (cm) |
Diameter (mm) | Number |
Fresh weight (g) | ||
| T1 | 61.1 ± 37.3 | 34.4 ± 6.9 | 3.5 ± 1.8 | 413.6 ± 176.3 | 148.5 ± 95.1 | 19.1 ± 4.5 | 4.5 ± 3.7 | 1430.0 ± 2078.1 | |
| T2 | 55.4 ± 16.5 | 38.3 ± 10.3 | 4.4 ± 2.0 | 487.3 ± 308.7 | 169.4 ± 65.4 | 19.4 ± 6.6 | 7.4 ± 4.3 | 1530.0 ± 1670.2 | |
| T3 | 58.6 ± 6.8 | 38.9 ± 11.7 | 5.8 ± 1.5 | 682.0 ± 510.0 | 140.5 ± 49.1 | 19.3 ± 3.6 | 6.2 ± 3.3 | 1055.0 ± 512.7 | |
| T4 | 41.3 ± 16.0 | 27.1 ± 9.5 | 3.2 ± 1.1 | 278.0 ± 110.5 | 111.0 ± 21.3 | 16.9 ± 4.1 | 6.2 ± 2.7 | 777.0 ± 359.6 | |
| Average | 55.7 ± 25.1 | 35.2 ± 9.8 | 4.1 ± 1.9 | 458.8 ± 300.1 | 147.2 ± 71.3 | 18.8 ± 4.9 | 6.0 ± 3.7 | 1288.7 ± 1546.9 | |
| Significancey | ns | ns | ns | ns | ns | ns | ns | ns | |
zT1, T2, T3, and T4 represent individual temperature zones in the temperature-gradient tunnel shown in Fig. 1.
지하경 수량을 포함한 전체 지하부 수량을 분석한 Table 17의 결과에서도 이러한 종간 차이가 더욱 뚜렷하게 확인되었다. 만주감초는 주근과 지하경의 총 수량이 30,731g으로, 주근:지하경 비율이 평균 1:1.7이었다. 반면 원감 품종은 총 수량 49,966g 중 약 91%가 지하경에서 기인하였으며, 주근:지하경 비율은 평균 1:10.1로 매우 높은 지하경 비중을 보였다. 또한 지하부의 생식자원 배분율(RA)도 만주감초는 61.9% 였으나 원감 품종은 90.5%로 매우 높게 나타났다. 이는 원감 품종이 후기 생장 단계에서 지하경 중심으로 에너지와 자원을 재분배함으로써, 지하경 기반의 영양번식과 생태적 확산 전략을 강화하고 있음을 시사한다.
Table 17.
Main root and rhizome yield and resource allocation ratio by temperature section in Five-year-old licorice (unit: g)
| Category | Main root | Rhizome |
Total biomass (root + rhizome) |
Root-to-rhizome ratio |
RAz (%) | |
| G. uralensis | T1 | 3,886.7 ± 197.3 a | 4,326.7 ± 670.2 b | 8,213 ± 862 a | 1:1.1 | 52.7 |
| T2 | 3,415.0 ± 551.2 a | 6,265.0 ± 420.9 a | 9,680 ± 217 a | 1:1.8 | 64.7 | |
| T3 | 2,803.3 ± 1,042.0 ab | 5,935.0 ± 849.6 a | 8,738 ± 1821 a | 1:2.1 | 67.9 | |
| T4 | 1,590.0 ± 832.9 b | 2,510.0 ± 748.9 c | 4,100 ± 1576 b | 1:1.6 | 61.2 | |
| Total | 11,695 | 19,037 | 30,731 | 61.9 | ||
| significancey | * | *** | ** | - | ||
| Wongam | T1 | 1,516.7 ± 525.0 a | 12475.0 ± 3319.2 a | 13,992 ± 3234 a | 1:8.2 | 89.2 |
| T2 | 1,624.2 ± 308.6 a | 14956.7 ± 3109.9 a | 16,581 ± 3394 a | 1:9.2 | 90.2 | |
| T3 | 1,136.7 ± 389.7 ab | 12061.7 ± 273.9 a | 13,198 ± 126 a | 1:10.6 | 91.4 | |
| T4 | 463.3 ± 144.3 b | 5731.7 ± 387.5 b | 6,195 ± 273 b | 1:12.4 | 92.5 | |
| Total | 4,741 | 45,225 | 49,966 | 90.5 | ||
| significancey | * | ** | ** | - | ||
|
Yield Change (%) of Wongam (vs. G. uralensis) | ‒59.5 | +137.6 | +62.6 | - | ||
또한 Table 12의 지상부 개화 및 결실 비교 결과와 결부하여 해석해 볼 때, 두 종이 상이한 번식 전략을 취하고 있음이 보다 뚜렷해진다. 5년간의 생장 과정에서 원감 품종은 2년차부터 생식 생장에서 현저히 우위를 보였다. 그러나 4년차에는 개화수와 결실수 모두에서 만주감초와 유사한 수준으로 떨어졌다. 이와 같은 생장 연차에 따른 개화 및 결실수 변화와 지하경 생체량의 분포 차이를 종합적으로 고려할 때, 만주감초는 지하경 생장보다 지상부 생식 기관 발달을 점진적으로 강화하여, 후기까지도 개화 및 결실수를 증가시키는 양상을 보였다. 이는 생장 전반에 걸쳐 유성번식을 유지하려는 경향으로 해석된다. 반면 원감 품종은 생장 초기부터 만주감초보다 우세한 지상부 생장을 바탕으로 조기에 유성생식에 집중한 이후, 후기에는 지하경 생체량을 증대시키며 자원 배분 전략을 전환하는 양상을 보였다. 결과적으로 두 품종은 생장 시기별로 번식 자원 분배 방식에 차이를 보이며, 유성번식과 영양번식 사이에서 상이한 전략적 균형을 유지하는 것으로 판단된다.
고 찰
본 연구는 감초 두 종(G. uralensis 및 G. korshinskyi)을 대상으로 장기간 온도구배 조건에서 유성번식 및 영양번식 특성을 비교하여, 품종 간 자원 배분 전략, 지하부 구조의 발달, 고온 반응성, 그리고 생리적 적응성에서의 차이를 분석하였다. 이를 통해 감초의 재배 목적별 종 선택 및 관리 전략 수립을 위한 생리·생태학적 기초자료를 제시하였다.
첫째, 감초 두 종 간 번식을 위한 자원 배분 전략에는 뚜렷한 차이가 확인되었다. 일반적으로 꽃눈 분화에 영향을 미치는 식물 호르몬은 GA, ABA, ZR, IAA 등이 알려져 있는데, 이중 IAA와 ZR은 지상부 생장에도 중요한 역할을 한다(Blázquez and Weigel 1999). Yan et al.(2019)은 만주감초의 개화한 개체가 개화하지 않은 개체보다 IAA와 ZR 수치가 높았다고 하여 영양생장과 개화수와의 관련성을 시사한 바 있다. 원감(G. korshinskyi)의 경우도 빠른 영양 생장을 바탕으로 초기부터 생식에 집중하는 모습을 보였다. 반면 만주감초(G. uralensis)는 초기에는 생식을 지연하고 영양생장에 더 많은 자원을 투자한 후 후기 생식에 집중하는 차이점을 보였다. 식물의 자원배분 전략은 환경 적응성과 생육 연한에 따라 다양하게 나타날 수 있다. 다년생 식물의 경우 첫 번째 계절 이후 번식을 지연하는 경우가 있다고 하였다(Lundgren and Des Marais 2020). Lupinus 속에서도 1년생과 다년생 간에 생식과 영양기관 유지에 대한 에너지 배분 전략이 다르게 나타난다고 보고된 바 있다(Pitelka 1977; Lundgren and Des Marais 2020). 종자 생산과 영양번식은 종종 상충관계에 놓인다. Butomus umbellatus와 Sagittaria latifolia의 경우 종자자원 투자가 증가할 경우 클론 번식체의 생산을 감소시킨다고 보고되었다(Thompson and Eckert 2004; Van Drunen and Dorken 2012; Li et al. 2018). 감초 실험에서 확인된 두 품종 간 자원 배분 차이 또한, 기존 선행 연구에서 보고된 식물들의 사례처럼 유성생식과 영양생장이 환경에 따라 경쟁적으로 조절된다는 점에서 어느 정도 공통된 양상을 보인다. 이는 종자 생산 등 재배 목적에 따라 품종 선택 및 생육 환경 관리를 달리할 필요가 있음을 시사한다.
둘째, 지하부 생체량 구성에서도 두 종은 상이한 전략을 보였다. 5년차 조사에서 만주감초는 뿌리와 지하경의 지속적인 비대를 나타낸 반면, 원감 품종은 주로 지하경 비대에 집중되어 전체 지하부의 91%를 차지하였다. 이는 만주감초가 후기 유성생식 기반의 전략을 취하고 있는 반면, 원감 품종은 높은 생식자원 배분률(RA)에서도 확인되듯이 지하경을 통한 영양번식 확산 전략을 택하고 있음을 보여준다. Li et al.(2018)은 수생식물인 Vallisneria spinulosa의 경우 수심에 따라 유성생식이 불리해지면 괴경 생산에 더 많은 자원을 투자한다고 하였으며, 생식자원 배분률(RA) 변화는 환경 변화 속에서 식물의 적응도를 유지하는데 중요한 역할을 한다고 하였다. 또한 Carex Brevicuspis는 비옥한 토양에서 유성생식보다 무성생식에 더 많은 자원을 투자한다고 보고되었다(Chen et al. 2015; Li et al. 2018). 이러한 경향성은 강우량이 많고 토양비옥도가 높은 한국의 재배 환경에 적응성이 높은 원감 품종(G. korshinskyi)이 지하경 수량이 더 많은 것과도 유사성이 있다. 이러한 두 종간의 지하부 생체량 구성 차이는 장기재배에 따른 지하경 생산을 고려할 때 중요한 참고 정보가 될 수 있을 것이다.
셋째, 고온 환경은 두 종 모두의 생식 생장에 부정적인 영향을 미쳤으나, 그 민감도에는 차이가 있었다. 고온 환경에서는 식물체가 수분 손실을 줄이기 위해 기공을 닫는데 이를 위해 ABA를 증가시키는 경향이 있다(Chen et al. 2015). Yan et al.(2019)은 만주 감초의 꽃차례가 탈리된 개체에서 GA와 ABA가 높게 나타났다고 하였는데, 이는 고온구간에서 결실률이 특히 떨어지는 이유를 설명해 준다. 감초 종별로 볼 때, 만주 감초는 고온 구간에서 개화가 현저히 줄어들고 결실도 거의 발생하지 않았으나, 원감 품종은 비교적 높은 개화율과 결실률을 유지하였다. 이는 원감 품종이 고온 조건에서 일정 수준 이상의 생식 생장을 유지할 수 있는 생리적 내성을 갖고 있음을 의미한다. 많은 식물들에게 있어서 생식기의 꽃가루의 생존력, 수정 및 배아 발달은 특정 한계 온도를 넘어서는 열에 민감한 것으로 알려져 있다(Thompson and Eckert 2004; Van Drunen and Dorken 2012; Li et al. 2018). 예를 들어 벼(Oriza sativa)의 경우 개화기가 고온에 가장 취약하고, 이삭단계에서는 수량과 품질을 심각하게 감소시킬 수 있다고 보고된 바 있다(Prasad et al. 2006; Jagadish et al. 2007; Resentini et al. 2023). 또한 품종별로도 열스트레스 내성이 달라 수정 능력이 차이가 있음이 보고 되었다(Shi et al. 2022; Resentini et al. 2023). 특히 환경 적응성이 큰 표현형 가소성을 보이는 작물의 경우 수량 안정성 확보에 유리하다고 하였다(Resentini et al. 2023). 이는 감초종별로 온도관리를 달리할 필요가 있으며, 특히 G. korshinskyi와 같은 감초종의 경우 기후변화에 따른 고온 환경에 유리한 적응 품종으로 활용될 수 있음을 시사한다.
넷째, 신초는 모주에 비해 고온 환경에서 상대적으로 안정적인 영양 및 생식 생장을 보였으며, 이는 생리적 보상 또는 회피 메커니즘일 가능성이 있다. 불량 환경에 대한 회피 메커니즘은 다양한 식물에서 관찰된다. 여러번 꽃을 피우는 카놀라(Brassica napus L.)에 2주간 열스트레 처리한 결과 기형 수술이 증가하고 꽃이 왜소해져 꼬투리(silique) 수량이 90% 이상 감소하였으나 다음 결실에서는 오히려 꼬투리 수량이 11–18% 증가하였다고 보고된 바 있다(Pokharel et al. 2021). McGregor(1981)도 카놀라의 열 스트레스가 해소된 후 새 꽃차례에 추가 가지나 꽃이 피거나 꽃줄기당 더 많은 종자가 생겨 상당한 회복이 이루어질 가능성을 언급했다. 다년생인 감초는 지하경을 통해 영양개체(ramets) 간 서로 연결되어 있어 양분과 신호전달이 가능한 클론 집단이다. 이러한 지하부 구조체계는 신초수와 뿌리 밀도 조절 및 유성생식 등에 대한 양분 배분에 클론 집단 차원에서 관여하기 용이하며, 환경 적응을 위한 수단으로 활용될 수 있다. 이러한 현상은 생리적 통합으로도 알려져 있다(Ashmun et al. 1982; Alpert 1996; Guo et al. 2020). Guo et al.(2020)은 지하경을 통해 번식하는 다년생 초본식물인 가시벼풀(Leymus chinensis) 개체군에서도 연결된 영양개체(ramets)를 통해 화서 바이오매스, 종자수, 생식 할당 등의 조절 가능성을 언급한 바 있다. 또한 클론 식물의 자원이동은 대부분 acropetal 이동(발달적으로 오래된 ramets에서 어린 ramets으로 이동)이 basipetal 이동(어린 ramets에서 늙은 ramets로 이동) 보다 더 일반적이라고 알려져 있다(Alpert 1996; de Kroon et al. 1996; D'hertefeldt and Jónsdóttir 1999; Guo et al. 2020). 이러한 생리적 통합 기능은 폭넓은 환경 스펙트럼 속에서 감초의 신초가 모주에 비해 생육 및 번식에 더 안정적일 수 있는 이유를 설명해 준다. 한편 이러한 적응 전략에도 불구하고, 실제 결실률이 신초에서 더 낮게 나타난 점은 조직의 생리적 미성숙의 한계로 해석해 볼 수 있다.
종합적으로 볼 때, 감초 두 종은 온도 및 생육기간에 따라 상이한 생식·영양 번식 전략을 취하고 있었으며, 이러한 차이는 종자 생산 또는 지하부 수확을 주요 재배 목적으로 할 경우 품종 선택에 중요한 기준이 될 수 있다. 원감 품종은 고온 환경에 강하고, 만주감초는 후기까지 일정한 수준의 결실을 보였다. 만주 감초는 뿌리 비대가 특징이어서 전통적인 약재 생산에 유리하고, 원감 품종은 지하부 비중이 높아, 기존의 재배 감초에서 활용이 제한되었던 지하경을 약재 자원으로 활용할 수 있는 가능성이 있어, 각 종의 특성에 따라 적정 재배 연한과 관리 전략을 설정할 필요가 있다. 본 연구는 이러한 전략 수립을 위한 생리·생태학적 기초 자료를 제공하며, 향후 감초의 자급률 향상과 기후 적응형 재배 기술 개발에도 기여할 수 있을 것이다.
