

Introduction
Materials and Methods
Experimental plants and environmental conditions
Investigation of morphophysiological characteristics
Statistical analysis
Results and Discussion
Survival and success rates
Morphophysiological analysis
Conclusions
Introduction
Vegetative propagation methods such as stem, leaf, petiole, and root cuttings are highly effective when used to preserve the genetic stability of certain cultivars, including F1 hybrids and haploids, as well as variegated forms of plants, and specific mutations. Although some variegated traits may be lost through cutting propagation methods, maintaining these traits is significantly more feasible in this way than through seed propagation (Matsuo et al. 1989). These propagation methods not only preserve the characteristics of the mother plant but also allow for rapid mass propagation over a short period of time (Tchoundjeu et al. 2004; Ranawana and Eeswara 2008; Zhang et al. 2013). This approach is widely used in farms and plant nurseries because it is easy to implement and does not have the drawbacks associated with seed propagation (Kim and Kim 2015; Kwon and Cho 2023).
Among these methods, the stem-cutting method is particularly versatile as it can be used for plants in which leaf or petiole cuttings are not feasible, exhibiting a low risk of failure due to the development of axillary buds. The efficiency and reduced cultivation period of vegetative propagation are influenced by various environmental factors, such as the culture media, growth temperature, relative humidity, light intensity, light quality, soil moisture content, and the concentration of endogenous or exogenous phytohormones. Among these factors, phytohormones play a crucial role in regulating plant growth and development, exerting significant physiological effects even in minute quantities (Sosnowski et al. 2023).
Natural phytohormones, such as auxins, cytokinins, and gibberellic acid (GA), promote plant growth and development. In contrast, abscisic acid (ABA) significantly inhibits these processes, and ethylene is known for its role in fruit ripening (Nam et al. 2024). Among these, auxins, derived from the Greek word ‘auxein’ meaning ‘to grow’, are involved in almost all aspects of plant growth and development (Enders and Strader 2015; Dong and Huang 2018; Luo et al. 2018; Basunia and Nonhebel 2019; Tivendale and Millar 2022). Notably, auxins play a pivotal role in the development of adventitious roots, a critical process for plant propagation and stress adaptation (Meng et al. 2019; Wang et al. 2021b; Oh et al. 2023; Sekhukhune and Maila 2025). Auxins are found in plants, fungi, and microbes in nature and are known to play crucial roles as key regulators of plant growth and development by integrating abiotic stress signals and controlling downstream stress responses (Spaepen and Vanderleyden 2011; Keswani et al. 2020; Jing et al. 2023; Thakor et al. 2023).
Since the discovery of auxins in the 1930s, various propagation methods utilizing auxins, such as the basal quick-dip, powder application, and the dilute soak methods, have been developed, with the first two methods still widely used today (Blythe et al. 2007). Auxins exist in natural forms such as indole acetic acid (IAA), phenylacetic acid (PAA), and 4-chloroindole-3-acetic acid (4-Cl-IAA) and in synthetic forms such as 3-indolebutyric acid (IBA), 1-naphthaleneacetic acid (NAA), and 2,4-dichlorophenoxyacetic acid (2,4-D) (Enders and Strader 2015). These forms are used not only to promote plant growth and development but also as herbicides (Mithila et al. 2011; Peterson et al. 2016).
Auxin-based rooting promoters, also known as auxinic rooting promoters, have been shown to be effective when used for stem cuttings in various species, such as Apocynum lancifolium (Seo et al. 2023), Echeveria spp. (Cabahug et al. 2016), Philodendron hederaceum var. oxycardium (Lee et al. 2024), Sageretia thea (Son et al. 2022), and Sedum takesimense (Oh and Lee 2022). Previous studies have investigated the effects of the dipping time on root formation (Wang et al. 2021a; Bak et al. 2023), with some focusing on different types of auxins and their varying rates of efficacy in promoting rooting (Kwon and Cho 2023; Lee et al. 2024). Others have explored foliar application methods for propagation and growth promotion (Blythe et al. 2004; Mir et al. 2020; Bowden et al. 2022; Jung et al. 2023). Given these factors, careful selection of auxin types, concentrations, and application methods is crucial, as they can significantly influence various aspects of plant growth and development depending on the species or cultivar (Lee et al. 2024). This highlights the need for diverse experimental approaches to achieve optimal results.
Most cutting experiments on the genus Hedera have focused on H. helix (Poulsen and Andersen 1980; Richardson and Humphries 1982). The physiological characteristics of H. algeriensis, commonly known as Algerian ivy, are largely unknown, with relevant studies being limited primarily to some morphological and taxonomic research (Ackerfield and Wen 2002; Valcárcel et al. 2003; Valcárcel and Vargas 2010). The variegated H. algeriensis cultivar, specifically ‘Gloire de Marengo’, which possesses a tricolor pattern, is distributed globally and is noted for its aesthetically appealing appearance and high demand. However, the optimal auxin concentration for the cuttings of this cultivar has not yet been determined. Thus, exploring auxinic rooting promoters is essential to enhance the propagation efficiency and morphophysiological characteristics of this cultivar.
Ornamental foliage plants such as ‘Gloire de Marengo’ are highly valued for their ability to enhance both indoor and outdoor environments, including homes, offices, gardens, and public spaces (Bermejo and Sparke 2018; Lee et al. 2021; Persiani 2021; Francini et al. 2022). Beyond their aesthetic appeal, these plants contribute to improving air quality levels by reducing fine dust and other airborne pollutants (Jeong et al. 2020, 2021), and they offer significant benefits to the physical and mental health of humans by creating more pleasant and stress-reducing environments (Deng and Deng 2018; Tao et al. 2020). The increasing demand for ornamental foliage plants such as ‘Gloire de Marengo’ underscores the importance of efficient propagation methods. Given the importance of preserving the distinct variegated traits of this cultivar, the use of auxinic rooting promoters in stem-cutting propagation is particularly advantageous. This approach not only overcomes the limitations associated with seed propagation, which can usually lead to a loss of variegation, but also enables the rapid, mass production necessary to meet market demands.
Therefore, in this study, H. algeriensis cv. Gloire de Marengo, a variegated ornamental foliage plant, serves as the experimental plant to investigate the effects of auxinic rooting promoters on the propagation efficiency and morphophysiological characteristics of its stem cuttings.
Materials and Methods
Experimental plants and environmental conditions
This study was conducted over four months, from February to June 2024, in the experimental greenhouse of the Department of Environmental Horticulture at Sahmyook University, located in Nowon-gu, Seoul, South Korea. The experimental greenhouse had a 25% shading level and was open on all sides.
The auxinic rooting promoters used in this study were 3-indolebutyric acid (IBA) and 1-naphthylacetic acid (NAA) (Sigma-Aldrich, USA). Nine concentrations were designed: control (T1), IBA at 250, 500, 750, and 1000 mg·L-1 (T2, T3, T4, and T5, respectively), and NAA at 250, 500, 750, and 1000 mg·L-1 (T6, T7, T8, and T9, respectively), based on the methods of Lee et al. (2024), with modifications to types and concentrations of auxinic rooting promoters in this study. All auxins used in this study were first dissolved in a small amount of 95% ethanol and then diluted with purified water until the final ethanol concentration reached 20% (Table 1). The control group consisted of 20% ethanol without any auxinic rooting promoter.
Table 1.
Concentrations and molarity values of the auxinic rooting promoters used in this study
| Treatment symbolsz | Auxinic rooting promoters | Concentrations (mg·L-1) | Molarity (mM) |
| T1 | Control | 0 | 0 |
| T2 | 3-indolebutyric acid (IBA) | 250 | 1.23 |
| T3 | 500 | 2.46 | |
| T4 | 750 | 3.69 | |
| T5 | 1000 | 4.92 | |
| T6 | 1-naphthylacetic acid (NAA) | 250 | 1.34 |
| T7 | 500 | 2.69 | |
| T8 | 750 | 4.02 | |
| T9 | 1000 | 5.37 |
zThese treatments were based on methods of Lee et al. (2024), with modifications to types and concentrations of auxinic rooting promoters.
Rectangular pots measuring 48.5 × 33.0 × 8.0 cm (length × width × height) were used. The culture medium consisted of a 1:1:1 (v/v/v) mixture of a fertilized horticultural substrate (Hanareumsangto, Shinsung Mineral, South Korea), vermiculite (Verminuri, GFC, South Korea), and perlite (New Pearl Shine No. 1, GFC, South Korea). Stem cuttings were prepared from one-year-old ‘Gloire de Marengo’ plants, with each cutting consisting of a single 3 cm node, one axillary bud, and a single leaf to ensure uniformity in the experimental conditions. The dipping method involved immersing the bases of the stem cuttings to a depth of 5 mm for one minute. Watering was carried out twice a week, and liquid fertilizer with an N-P-K ratio of 7-10-6 (High Grade S, HYPONeX, Japan) was applied at a concentration of 1000 ppm once a month. During the study, the average temperature and relative humidity were 21.4 ± 3.9°C and 60.3 ± 12.5%, respectively.
Investigation of morphophysiological characteristics
To investigate the effects of auxinic rooting promoters on the propagation efficiency and morphophysiological characteristics of ‘Gloire de Marengo’ stem cuttings, various parameters were measured, in this case the survival rate, shooting and rooting success rates; shoot length and width, stem diameter; root length; ground cover; number of leaves, the leaf length, width, and area; the fresh weights of the shoots and roots; the dry weights of the shoots and roots; the total fresh and dry weights; the moisture content of the shoots and roots; the compactness index; the Dickson quality index (DQI); the normalized difference vegetation index (NDVI); the modified chlorophyll absorption ratio index (MCARI); the maximum quantum yield (Fv/Fm); and the photosynthetic performance index (PIABS).
Plant sizes were measured following the methods described in Park et al. (2023a). To ensure accurate measurements, the shoot length was measured from the ground to the terminal bud of the plant after the shoot was completely straightened, i.e., contained no bends. The shoot width was measured as well after the shoot was completely straightened at the widest part when viewed from the front. Root length was determined by measuring the longest root. Dry weight was obtained after drying the samples at 85°C for 12 hours using a hot-air drying oven (HK-DO135F, Hankuk S&I, South Korea) and then weighing them.
To evaluate the moisture content and quality of the plants, the moisture content (Lee and Nam 2024) (Eq. 1), compactness index (Kim et al. 2016) (Eq. 2), and DQI (Dickson et al. 1960) (Eq. 3) were analyzed.
(FW, fresh weight; DW, dry weight; SDW, shoot dry weight; SL, shoot length; SD, stem diameter; RDW, root dry weight; TDW, total dry weight)
Remote sensing vegetation indices were measured using a portable device specialized for measuring vegetation indices (PolyPen RP 410, Photon Systems Instruments, Czech Republic). The relevant equations for the NDVI (Rouse et al. 1973) (Eq. 4) and MCARI (Daughtry et al. 2000) (Eq. 5) are given below. These indices were used to evaluate the physiological status of ‘Gloire de Marengo’ as influenced by the auxinic rooting promoters.
A chlorophyll fluorescence analysis was conducted using a detachable dark-adapted leaf clip following the manufacturer’s guidelines. Newly expanded leaves were randomly selected from the stem cuttings, dark-adapted for 15 minutes (PSI 2024), and measured using a portable chlorophyll fluorescence meter (FluorPen FP 110/D, Photon Systems Instruments, Czech Republic). Two chlorophyll fluorescence parameters were evaluated to assess changes in the physiological status of photosystem II (PSII). The descriptions and equations used are based on work by Stirbet and Govindjee (2011) and on the PSI (2024) manufacturer’s guidelines, as described below.
The maximum quantum yield, represented by Fv/Fm (Eq. 6), and the photosynthetic performance index, PIABS (Eq. 7), which together indicate the overall vitality and efficiency of PSII, were investigated. Chlorophyll fluorescence measurements were specifically obtained from the dark green areas of the tricolor leaf patterns while avoiding the light green and white patterns as well as the leaf veins.
Statistical analysis
The experimental results were analyzed using SAS 9.4 (SAS Institute, USA) by means of a one-way analysis of variance (ANOVA). Duncan’s multiple range test was applied for mean comparisons at a significance level of p < 0.05. The experimental design was a completely randomized design (CRD), with seven stem cuttings per replication and a total of five replications per treatment.
Results and Discussion
Survival and success rates
In the nursery industry, rooting promoters are applied to stem cuttings to induce the development of adventitious roots across a wide range of plant species and cultivars, with the aim of accelerating root growth and development in cuttings (Blythe et al. 2007; Crawford et al. 2016). Owing to their greater stability against heat and light degradation, both IBA and NAA are more effective compared to IAA in promoting adventitious root development (Chadwick and Kiplinger 1939). Therefore, these two auxins were used as auxinic rooting promoters in the present study. Recently, experiments involving the use of the basal quick-dip method and a foliar application, which are simple and promising propagation methods, have been conducted on plants of the genus Hedera (Blythe et al. 2004; Abu-Zahra et al. 2013). However, these studies are relatively less sophisticated and tend to provide limited information. In this study, we aim to address these gaps by systematically evaluating the effects of auxinic rooting promoters on the propagation efficiency and morphophysiological characteristics of H. algeriensis cv. Gloire de Marengo stem cuttings.
Photographic documentation from this study showing the growth evaluation of ‘Gloire de Marengo’ stem cuttings as influenced by different concentrations of auxinic rooting promoters is shown in Fig. 1. The results of this study showed that neither the type nor the concentration of auxins significantly influenced the survival rate of ‘Gloire de Marengo’ stem cuttings (Fig. 2A). Similarly, there were no significant differences in shooting success rates among the treatments (Fig. 2B). In contrast, the rooting success rate was lowest in the control (T1) group at 88.5%, whereas treatments T3–T5 (3-indolebutyric acid, IBA 500–1000 mg·L-1) and T7–T8 (1-naphthylacetic acid, NAA 500–750 mg·L-1) achieved a high rooting success rate of 100%, indicating that the use of auxinic rooting promoters was effective in promoting rooting in ‘Gloire de Marengo’ stem cuttings (Fig. 2C).

Fig. 1.
Growth outcomes of Hedera algeriensis cv. Gloire de Marengo stem cuttings as influenced by auxinic rooting promoters at various concentrations after four months of growth. Control (T1) uses purified water with ethanol; T2, T3, T4, and T5 are treated with 3-indolebutyric acid (IBA) at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively; and T6, T7, T8, and T9 are treated with 1-naphthylacetic acid (NAA) at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively.
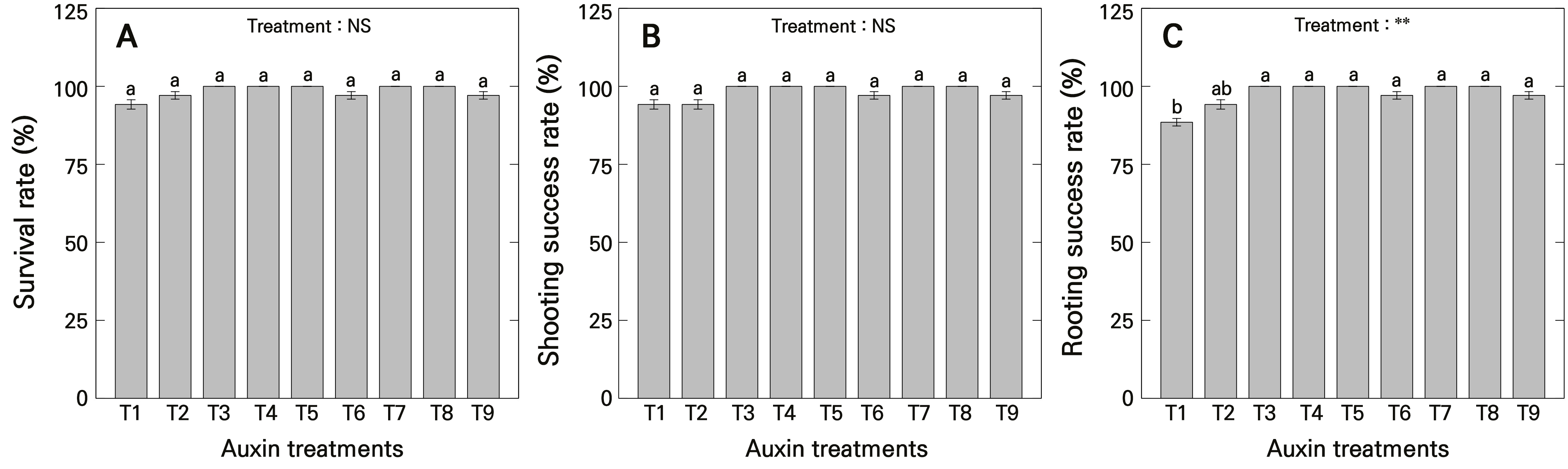
Fig. 2.
Survival rate and success rates of H. algeriensis cv. Gloire de Marengo stem cuttings as influenced by auxinic rooting promoters at various concentrations after four months of growth. A) survival rate; B) shooting success rate; and C) rooting success rate. T1 uses purified water with ethanol; T2, T3, T4, and T5 are treated with IBA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively; and T6, T7, T8, and T9 are treated with NAA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively. Means were derived from five replications (n = 5), each consisting of seven stem cuttings. Vertical bars indicate the mean ± standard error (SE), and asterisks (**) indicate significance at p < 0.01. Different lowercase letters indicate significant differences at p < 0.05, while the same lowercase letters indicate no significant difference at p > 0.05 based on Duncan’s multiple range test (DMRT). NS indicates no significant difference.
It is well known that IBA and NAA promote the development of adventitious roots in cuttings (Sourati et al. 2022). In particular, IBA, an auxin precursor, regulates IAA levels through β-oxidation (Aryal et al. 2019). High IBA concentrations can stimulate cell division by activating mitotic genes that are generally regulated by auxins (Qu et al. 2019). However, to optimize rooting rates, the concentration of auxins should be adjusted according to the treatment method and species characteristics (Sourati et al. 2022). While the combination of IBA and NAA could also be considered for potentially enhancing rooting success, it is essential initially to gain a thorough understanding of their independent effects to determine the optimal conditions for each auxin.
In a previous study by El Malahi et al. (2024), Damask rose (Rosa × damascena), a floricultural crop, showed the highest rooting and survival rates at an IBA concentration of 1000 ppm among the tested levels (i.e., 0, 1000, and 10000 ppm). In contrast, Oh and Lee (2022) found that Sedum takesimense had the highest rooting and survival rates at IBA 250–500 mg·L-1 and NAA 500 mg·L-1. In the case of Hedera sp., Abu-Zahra et al. (2013) reported that the rooting rate was highest at an NAA concentration of 3000 ppm. On the other hand, Eucalyptus benthamii showed a positive response to rooting at a high concentration of IBA 2000 mg·L-1 (Brondani et al. 2012). However, Kwon and Cho (2023) reported no differences in the survival and rooting rates of Pseudolysimachion ovatum across different concentrations of auxinic rooting promoters, suggesting that the response to auxinic rooting promoters varies by species and cultivar.
Consequently, the ‘Gloire de Marengo’ cultivar demonstrated successful propagation via stem cuttings; however, compared to the control, the application of auxinic rooting promoters has been found to improve the rooting success rate significantly.
Morphophysiological analysis
In this study, we observed that among the parameters related to plant size, shoot length was longest in the T3 and T7 treatments, measuring 33.71 and 32.44 cm, respectively, while shoot width was widest in the T4 treatment (IBA 750 mg·L-1) at 16.41 cm (Table 2). Conversely, both shoot length and shoot width were shortest in the T1 treatment, measuring 19.63 and 12.40 cm, respectively. The stem diameter was thickest in the T4 and T8 treatments (at 0.32 cm) and thinnest in the T1 treatment (at 0.23 cm). Meanwhile, root length was longest in the T4, T5 (IBA 1000 mg·L-1), and T7 treatments, measuring 16.00, 15.94, and 15.71 cm, respectively. The ground cover was widest in the T4 and T5 treatments, measuring 270.4 and 273.7 cm2, respectively. Previous studies have shown that S. takesimense exhibits greater root length at IBA and NAA concentrations of 500 mg·L-1 (Oh and Lee 2022) and that P. ovatum exhibits greater root length compared to a control treatment regardless of the type and concentration of auxin used (Kwon and Cho 2023). Similarly, Abu-Zahra’s study (2013) reported that Hedera sp. achieved the longest root lengths at NAA concentrations of 3000–4000 ppm. These findings highlight the fact that both the type and concentration of auxin are critical factors when seeking to optimize root development across species.
Table 2.
Plant sizes and leaf growth parameters of Hedera algeriensis cv. Gloire de Marengo stem cuttings as influenced by auxinic rooting promoters at various concentrations after four months of growth
| Treatment symbolsz | Plant sizes (cm) |
Ground cover (cm2) | Number of leaves | Leaf sizes (cm) | Leaf area (cm2) | ||||
| Shoot length |
Shoot width | Stem diameter |
Root length | Length | Width | ||||
| T1 | 19.63 dy | 12.40 c | 0.23 b | 8.08 c | 160.6 c | 4.1 c | 7.08 d | 6.69 c | 47.4 d |
| T2 | 24.31 bcd | 13.18 abc | 0.30 ab | 12.35 b | 190.1 abc | 5.5 ab | 7.19 cd | 7.25 abc | 52.4 cd |
| T3 | 33.71 a | 15.95 ab | 0.31 ab | 16.00 a | 262.3 ab | 6.5 ab | 7.75 bcd | 7.34 abc | 56.7 bcd |
| T4 | 29.38 abc | 16.41 a | 0.32 a | 15.94 a | 270.4 a | 6.2 ab | 7.98 bc | 7.57 ab | 60.2 abc |
| T5 | 23.84 bcd | 16.18 ab | 0.31 ab | 14.65 ab | 273.7 a | 5.5 ab | 8.87 a | 7.60 ab | 67.8 a |
| T6 | 23.24 cd | 13.15 abc | 0.28 ab | 14.48 ab | 179.0 bc | 5.1 bc | 7.34 cd | 7.39 abc | 54.6 cd |
| T7 | 32.44 a | 15.08 abc | 0.30 ab | 15.71 a | 228.6 abc | 7.1 a | 8.30 ab | 7.94 a | 65.8 ab |
| T8 | 30.37 ab | 13.72 abc | 0.32 a | 14.93 ab | 190.5 abc | 6.8 ab | 7.62 bcd | 7.09 abc | 54.3 cd |
| T9 | 28.37 abc | 13.01 bc | 0.28 ab | 15.35 ab | 171.2 c | 6.7 ab | 7.21 cd | 6.98 bc | 50.6 cd |
| Significancex | *** | * | * | *** | * | ** | *** | * | *** |
zControl (T1) uses purified water with ethanol; T2, T3, T4, and T5 are treated with 3-indolebutyric acid (IBA) at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively; and T6, T7, T8, and T9 are treated with 1-naphthylacetic acid (NAA) at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively.
Among the parameters related to the leaf size, the number of leaves was highest in the T7 treatment at 7.1 leaves and was lowest in the T1 treatment at 4.1 leaves. Meanwhile, leaf length and width were highest in the T5 and T7 treatments, measuring 8.87 and 7.94 cm, respectively, while the T1 treatment had the lowest values at 7.08 and 6.69 cm, respectively. Leaf area was greatest in the T5 treatment at 67.8 cm2.
Thus, the use of auxinic rooting promoters showed significant positive effects on parameters related to plant size. Although the optimal type and concentration of auxin varied for different parts of ‘Gloire de Marengo’ stem cuttings, the influence was positive with regard to the overall increase in the plant size parameters.
In previous studies by Wang et al. (2019, 2024), auxins were suggested to be involved in the photosynthetic efficiency of plants and to play a crucial role in biomass accumulation. However, this may be a complex outcome arising from the overall growth rate improvement owing to the promotion of root development by auxins, necessitating sufficient consideration of various growth factors (Tivendale and Millar 2022). The results regarding biomass revealed that the shoot fresh weight ranged from 7.73 to 8.13 g in the T3–T5 treatments and from 8.05 to 9.04 g in the T7–T9 treatments, indicating a high shoot fresh weight at concentrations of 500–1000 mg·L-1 regardless of the types of auxins used (Fig. 3A). However, root fresh weight was highest in the T7 treatment at 1.50 g. Shoot dry weight was heaviest in T3 and T7 treatments, measuring 1.55 and 1.61 g, respectively (Fig. 3B). Meanwhile, root dry weight was lowest in the T1 treatment (at 0.12 g), with no significant differences observed between the T2–T9 treatments. The total fresh weight followed a pattern similar to that of the shoot fresh weight, being heavier in the T3–T5 and T7–T9 treatments (Fig. 3C). The total dry weight was heaviest in T3 and T7 treatments, measuring 1.87 and 1.95 g, respectively, while the T1 treatment had the lowest total dry weight at 0.83 g. Regarding the moisture content, there were no statistically significant differences in the moisture contents of the shoots and roots of ‘Gloire de Marengo’ stem cuttings treated with various types and concentrations of auxinic rooting promoters (Fig. 3D). Consequently, the biomass results, similar to those related to the plant size, exhibited complex outcomes based on the type and concentration of auxinic rooting promoters, with the T3 and T7 treatments showing relatively high biomass outcomes.
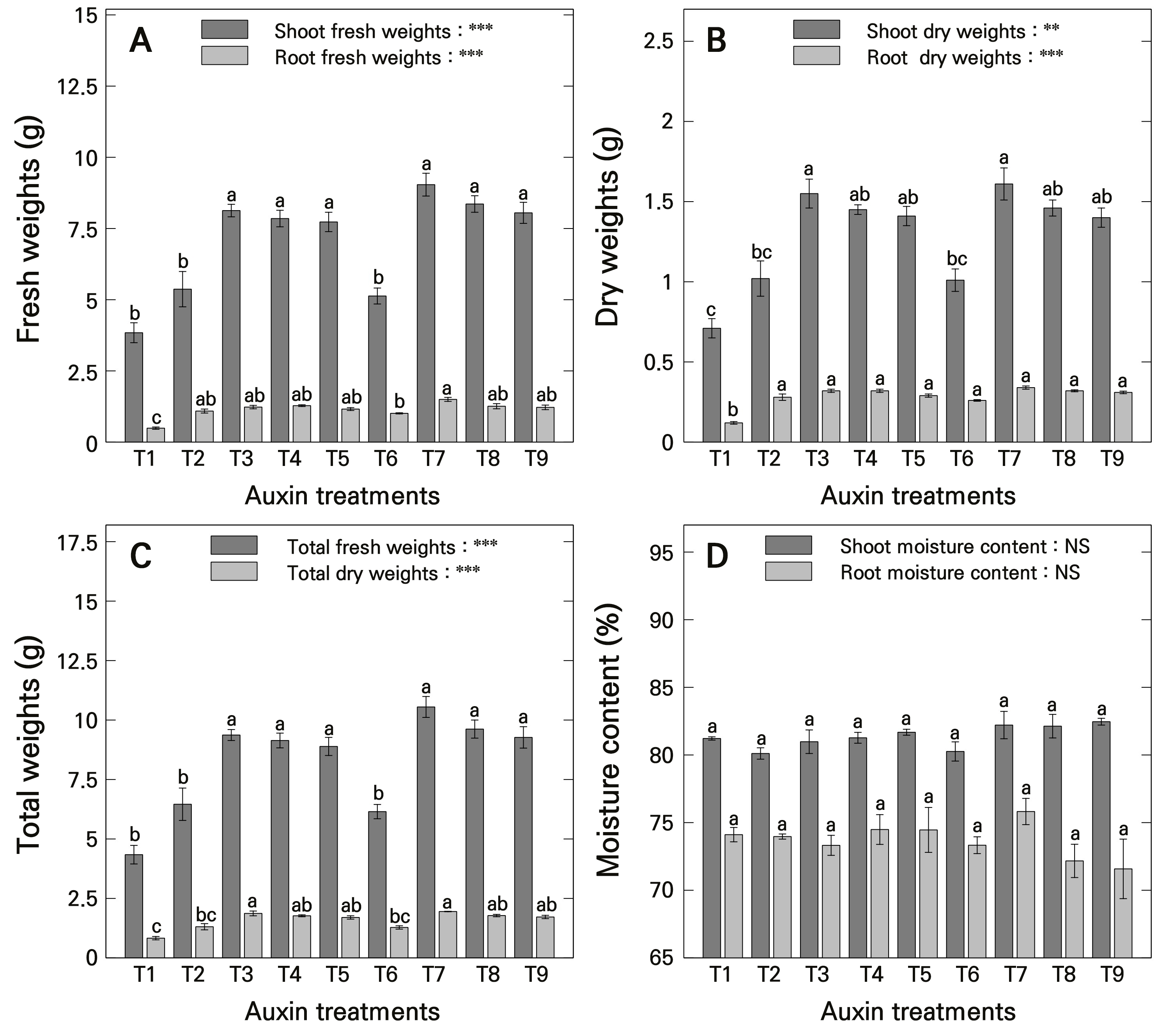
Fig. 3.
Plant biomass and moisture content of H. algeriensis cv. Gloire de Marengo stem cuttings as influenced by auxinic rooting promoters at various concentrations after four months of growth. A) fresh weights; B) dry weights; C) total weights; and D) moisture content. T1 uses purified water with ethanol; T2, T3, T4, and T5 are treated with IBA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively; and T6, T7, T8, and T9 are treated with NAA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively. Means were derived from five replications (n = 5), each consisting of seven stem cuttings. Vertical bars indicate the mean ± SE, and asterisks (** and ***) indicate significance at p < 0.01 and 0.001, respectively. Different lowercase letters indicate significant differences at p < 0.05, while the same lowercase letters indicate no significant difference at p > 0.05, based on DMRT. NS indicates no significant difference.
The compactness index of ‘Gloire de Marengo’ stem cuttings was highest in the T3 and T7 treatments, with values of 52.18 and 51.56, respectively (Fig. 4A). The DQI was lowest in the T1 treatment at 0.158, whereas it ranged from 0.361 to 0.450 across the other treatments, showing no significant differences regardless of the types and concentrations of auxinic rooting promoters used (Fig. 4B). Therefore, with regard to shoot growth, it was evident that the T3 and T7 treatments exhibited a high biomass density relative to the shoot length and shoot dry weight. From a broader perspective, all treatments except for the T1 treatment demonstrated relatively superior growth conditions for stem cuttings, indicating improved growth performance overall.
Among the methods used to assess plant physiological status, remote sensing vegetation indices and chlorophyll fluorescence analyses are widely utilized as non-destructive (or non-invasive) methods (Jang et al. 2023). These methods have been applied extensively in studies investigating plant physiological conditions (Kawakatsu and Fukuda 2023; Nam et al. 2023; Zheng et al. 2023; Lin et al. 2024). Such techniques are particularly effective for providing real-time data on plant health and stress responses, making them indispensable in studies focused on optimizing cutting propagation techniques (Kwon et al. 2023; Lee et al. 2024), enhancing growth and development (Guo et al. 2023; Shin et al. 2023; Kim et al. 2024b), and improving overall plant qualities (Ahn et al. 2024; Kim et al. 2024a, 2024c; Park et al. 2024).
One of the remote sensing vegetation indices, NDVI, was found to be high under the T3, T7, and T8 treatments, measuring 0.696, 0.702, and 0.694, respectively (Fig. 4C). Conversely, the NDVI values were lower in the T1, T5, and T6 treatments, with corresponding values of 0.620, 0.627, and 0.634. These results suggest that the use of auxinic rooting promoters positively influences the vegetation index of plants produced from stem cuttings and that excessively low or high auxin concentrations result in lower vegetation indices. This is in line with a previous study by Oh and Lee (2022), which reported that S. takesimense exhibited the lowest NDVI in the T1 treatment but showed higher NDVI values at IBA concentrations of 250–500 mg·L-1 and NAA concentration of 500 mg·L-1. Notably, NDVI values significantly decreased at a NAA concentration of 1000 mg·L-1, underscoring the importance of determining the optimal concentrations of exogenous auxins.
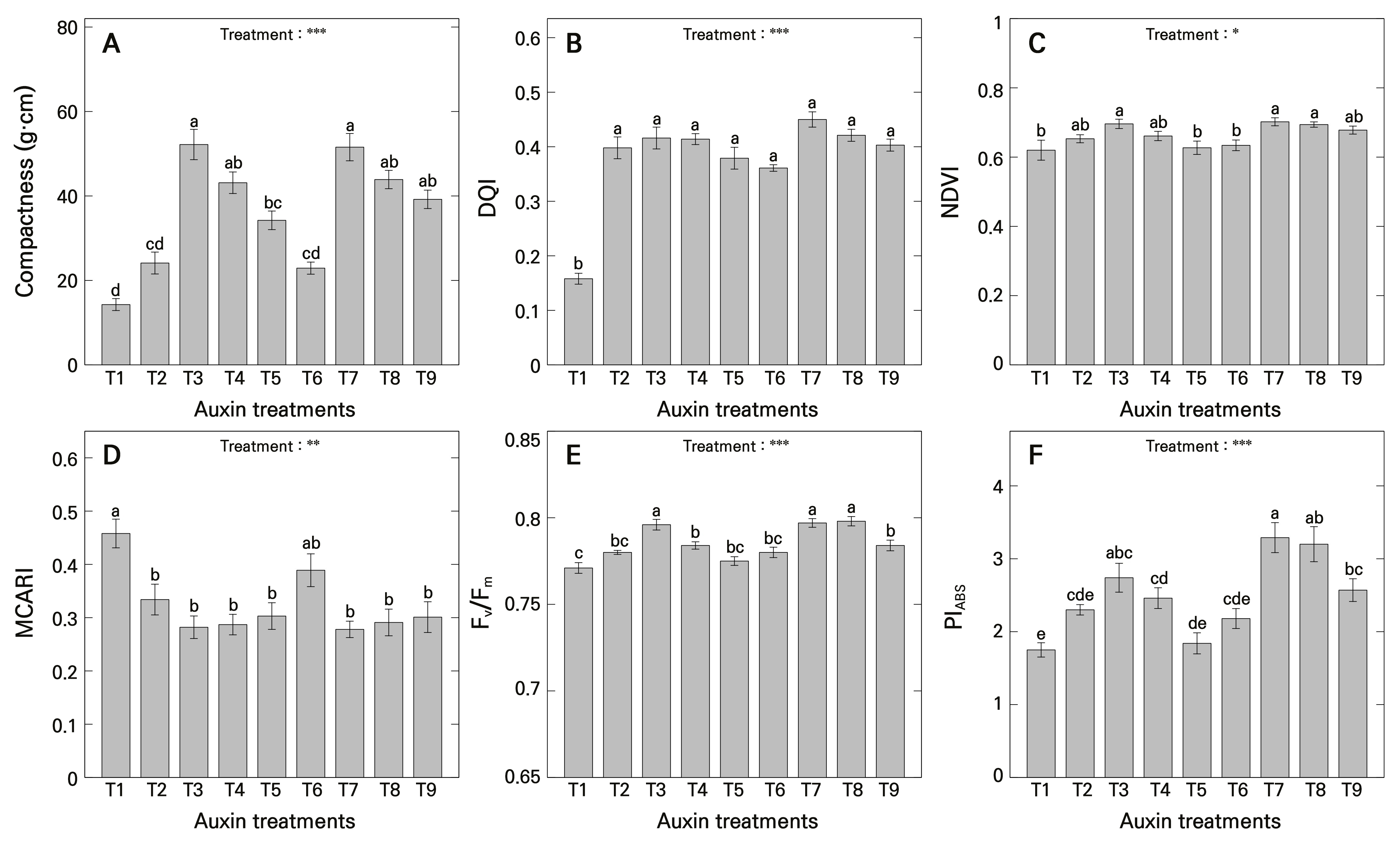
Fig. 4.
Plant growth quality indices, remote sensing vegetation indices, and chlorophyll fluorescence parameters of H. algeriensis cv. Gloire de Marengo stem cuttings as influenced by auxinic rooting promoters at various concentrations after four months of growth. A) compactness; B) Dickson quality index (DQI); C) normalized difference vegetation index (NDVI); D) modified chlorophyll absorption ratio index (MCARI); E) maximum quantum yield (Fv/Fm); and F) photosynthetic performance index (PIABS). T1 uses purified water with ethanol; T2, T3, T4, and T5 are treated with IBA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively; and T6, T7, T8, and T9 are treated with NAA at concentrations of 250, 500, 750, and 1000 mg·L-1, respectively. Means were derived from five replications (n = 5), each consisting of seven stem cuttings. Vertical bars indicate the mean ± SE, and asterisks (*, **, and ***) indicate significance at p < 0.05, 0.01, and 0.001, respectively. Different lowercase letters indicate significant differences at p < 0.05 based on DMRT.
MCARI was highest in the T1 treatment at 0.458 (Fig. 4D). MCARI is determined by the reflectance of the ground and the chlorophyll content of leaves and is generally inversely related to chlorophyll content (Nagler et al. 2000). Consequently, it was inferred that the T1 treatment had relatively lower chlorophyll content than the other treatments, except for the T6 treatment. Plants treated with exogenous auxins may exhibit either an increase or decrease in chlorophyll content or MCARI values, depending on the species. Previous studies have reported that auxins inhibit chlorophyll accumulation in Arabidopsis (Luo et al. 2023), whereas in crops such as wheat and soybeans, auxins positively influence chlorophyll content (Aldesuquy 2000; Gadallah 2000; Sosnowski et al. 2023). Considering the findings of this study and the results reported by Lee et al. (2024), the growth promotion observed in the auxin-treated stem cuttings is presumed to have a positive effect on chlorophyll content. As previously mentioned, initial chlorophyll content or MCARI values may vary depending on the treatment, but in the long term, chlorophyll content is expected to increase while MCARI values decrease.
In light of these findings, it appears that the use of auxinic rooting promoters positively influences the chlorophyll content in the stem cuttings of ‘Gloire de Marengo’ cultivar. However, further studies involving long-term and periodic monitoring of changes in chlorophyll content and MCARI values are necessary.
In the analysis of chlorophyll fluorescence responses, the maximum quantum yield of photosystem II (PSII) (Fv/Fm) was high under the T3, T7, and T8 treatments, measuring 0.796, 0.797, and 0.798, respectively (Fig. 4E). According to previous studies, an Fv/Fm range of 0.780–0.840 indicates that higher plants are not under stress (Björkman and Demmig 1987; Yoo et al. 2012; Asadi-Sanam et al. 2015; Park et al. 2023b). This Fv/Fm value is a valuable tool for assessing the stress status of cuttings, as cuttings with poor rooting may exhibit lower Fv/Fm values due to water stress. It serves as an effective tool for estimating the rooting potential of cuttings, allowing for a more accurate assessment of their physiological performance and likelihood of successful root development without physically interfering with the plants. In the present study, treatments T1 and T5 exhibited Fv/Fm values of 0.771 and 0.775, respectively, slightly below the normal range. McAdam et al. (2017) highlighted the significant potential of auxins as key developmental determinants of maximum leaf gas exchange, contributing to increased plant productivity. Therefore, the application of optimal concentrations of exogenous auxins is likely to improve the photosynthetic efficiency of the cuttings. The results of this study indicate that the use of auxinic rooting promoters increased the maximum quantum yield of ‘Gloire de Marengo’ cuttings, significantly improving the activity of the reaction centers of PSII. However, using relatively high concentrations of IBA, as observed in the T5 treatment, negatively influenced the maximum quantum yield, consistent with findings by Oh and Lee (2022).
The performance index (PIABS), a chlorophyll fluorescence parameter that indicates photosynthetic performance (Dai et al. 2019), is highly useful for assessing the overall physiological status of plants (Lee 2023). In agreement with the results observed for biomass, plant growth quality indices, and remote sensing vegetation indices, PIABS was highest in the T7 treatment at 3.29, whereas it was lowest in the T1 treatment at 1.75 (Fig. 4F). These findings suggest that the T1 treatment, with its relatively slow development of organs such as roots, likely experienced stress (Park et al. 2017). Delayed development of organs such as roots can lead to abiotic forms of stress, such as water stress, which in turn reduces photosynthesis, inactivates photosynthetic regulatory enzymes, and causes photoinhibition damage, thereby compromising the physiological health of plants (Medrano et al. 1997). The highest PIABS in the T7 treatment, which was found to have the best plant growth conditions, aligns with existing research reports showing that high PIABS values are correlated with healthy plant growth (Lee 2023; Sung et al. 2023; Lee and Nam 2024; Shin et al. 2024). Consequently, this implies that the significant promotion of growth in ‘Gloire de Marengo’ cuttings due to auxinic rooting promoters also elevated the performance index. Overall, optimal concentrations of auxinic rooting promoters were shown to provide considerable advantages in improving the physiological characteristics of ‘Gloire de Marengo’ stem cuttings.
In summary compared to the T1 treatment, the use of auxinic rooting promoters was found to be beneficial in that it improved the morphophysiological characteristics of ‘Gloire de Marengo’ stem cuttings. Specifically, by synthesizing the results of biomass, several growth quality indices, chlorophyll fluorescence parameters, and remote sensing vegetation indices, it was found that a concentration of 500 mg·L-1 NAA most effectively improved the morphophysiological characteristics of ‘Gloire de Marengo’ stem cuttings, followed by a concentration of 500 mg·L-1 IBA.
These findings provide valuable insights into the optimal concentration settings of auxinic rooting promoters and offer crucial guidance for improving the propagation efficiency and morphophysiological characteristics of ‘Gloire de Marengo’ stem cuttings. Future studies should focus on investigating the effects of the dipping time on this cultivar, as well as exploring combinations of different types of auxins and the potential benefits of foliar application methods. These approaches could further optimize propagation techniques, improve the overall quality of the propagated plants, and enhance the efficiency of mass production for ‘Gloire de Marengo.’ Additionally, expanding this research to similar species or cultivars could provide broader insights into the generalizability and application of these findings in other horticultural contexts.
Conclusions
This study was conducted to improve the propagation efficiency and morphophysiological characteristics of the variegated ornamental foliage plant H. algeriensis cv. Gloire de Marengo through stem cuttings using different types and concentrations of auxinic rooting promoters. Nine concentrations of auxinic rooting promoters were used: control; IBA at 250, 500, 750, and 1000 mg·L-1; and NAA at 250, 500, 750, and 1000 mg·L-1. In terms of the plant size, different concentrations of auxinic rooting promoters yielded varying results, with the use of IBA and NAA proving advantageous for producing larger plants than the control treatment. With regard to biomass, the shoot dry weight was highest at 500 mg·L-1 for both IBA and NAA, and the root dry weight was significantly higher regardless of the type and concentration of the auxinic rooting promoter used, compared to the control treatment. Overall, the total dry weight followed a pattern similar to that of the shoot dry weight, with both IBA 500 mg·L-1 and NAA 500 mg·L-1 being the most effective concentrations. Although the moisture content did not show significant differences among the treatments, the compactness index exhibited a trend similar to that of shoot dry weight and total dry weight, with this pattern being particularly evident in the IBA 500 mg·L-1 and NAA 500 mg·L-1 treatments. Furthermore, the DQI was higher for all treatments except for the control treatment, indicating that the use of auxinic rooting promoters could effectively improve plant quality levels during stem-cutting propagation of this cultivar. NDVI and the maximum quantum yield (Fv/Fm) were highest at IBA 500 mg·L-1 and NAA 500 and 750 mg·L-1, indicating superior physiological responses in these treatments. The MCARI and PIABS values were highest in the control and NAA 500 mg·L-1 treatments, respectively. This suggests a relatively low chlorophyll content in the control treatment and indicates that the use of NAA at a concentration of 500 mg·L-1 improves the integrity of the photosynthetic apparatus during stem-cutting propagation compared to the other treatments. In conclusion, the results of this study suggest that the most effective concentration for ‘Gloire de Marengo’ stem-cutting propagation is NAA 500 mg·L-1, followed by IBA 500 mg·L-1. Future studies should focus on investigating the effects of the dipping time on this cultivar and on exploring combinations of different types of auxins and discussing the potential benefits of foliar application methods. These approaches could further optimize propagation techniques, improve the overall quality of the propagated plants, and provide valuable insights into increasing the efficiency of the mass production of ‘Gloire de Marengo’.
