

Introduction
Materials and Methods
Plant material and vase solution
Number of bacteria and identification of microorganisms in the preservative solutions of cut gerbera flowers in a greenhouse
Antimicrobial proper treatment concentration of peracetic acid against Pseudomonas fluorescens
Quality analysis of cut gerbera flowers
Pseudomonas fluorescens antibacterial properties of peracetic acid in vase solutions
Experimental design and data analysis
Results
Discussion
Introduction
Gerbera is one of the most traded cut flower species globally (Mohsin et al. 2023; Kaya et al. 2025). It is susceptible to bending owing to its stemless structure, which is associated with a water imbalance, hormonal imbalance, and the softening of cellular tissue due to decreased water absorption. Water absorption decreases as a result of blocked vessels caused by bacterial growth in the vase solution (Steinitz 1983; Perik et al. 2012; Tonooka et al. 2021). Studies correlating the diversity of bacterial communities in vase solutions of cut roses, gerbera, and lilies have identified Pseudomonas spp. as associated species with vase life durations. Pseudomonas fluorescens is a common bacterium frequently detected in vase solutions of cut roses and gerbera (Kim et al. 1997; Balestra et al. 2005). At concentrations of 107–108 CFU·mL-1, P. fluorescens reduces the water absorption rate and relative weight of cut flowers, and the vase life is rapidly shortened due to scape bending (Li et al. 2012). Pseudomonas spp. is the primary bacterium that shortens gerbera vase life, and various disinfectants have been developed and studied to suppress it (Penningsfeld and Forchthammer 1966; de Witte et al. 2014). Various eco-friendly agents such as nanoparticle silver (Ag) and calcium oxide (CaO), melatonin (C13H16N2O2), nano-chitosan [(C6H11NO4)n], vinegar (CH3COOH), aluminum sulfate [Al2(SO4)3], calcium chloride (CaCl2), citric acid (C6H8O7), and thymol (C10H14O) have been studied extensively in relation to gerbera (de Silva et al. 2013; Salma et al. 2013; Atefepour et al. 2021; García-González et al. 2022; Timalsina et al. 2023; Meral et al. 2024).
The bactericidal effect of peracetic acid (PAA) is based on the release of free oxygen, and it is known that PAA oxidizes the bonds of the hydrogen sulfide group (-SH) and sulfur in the proteins, enzymes, and other metabolites of microorganisms, disrupting double bonds and sterilizing them (Liberti et al. 1999). Low-temperature storage for 90 days after a treatment with 0.1–0.5% PAA in pear fruit resulted in nearly half the incidence of black spot disease compared to no treatment, and antifungal effects were observed even at concentrations twice lower than sodium hypochlorite (Yang et al. 2022). In strawberries, decay caused by postharvest Botrytis cinerea infection was delayed during a PAA (100 µL·L-1) spray treatment at the immature stage, when peel color was changing from green to white before harvest (Narciso et al. 2007). In tomatoes, a head spray of a PAA (80 mg·L-1) treatment inhibited Salmonella enterica (Chang and Schneider 2012). Additionally, foliar fertilization with PAA (2.7 g·L-1) environmentally controlled nematodes for three days during host cultivation (Jagdale and Grewal 2002).
When a postharvest vase solution was treated with PAA (0.1 mL·L-1) and 3% sucrose, the vase life of lisianthus increased by 30% compared to tap water (Riva and Mazuela 2009). A treatment with PAA (50 µL·L-1) in roses maintained a higher flowering rate, greater relative fresh weight, and better water balance, extending the vase life compared to when distilled water was used (Ma and Lee 2025). Postharvest quality management studies pertaining to PAA have mainly focused on the sterilization of fruits and vegetables, and research on its role in extending vase life durations by inhibiting bacterial growth in the vase solutions and stems of cut gerbera flowers is thus far insufficient (Hong et al. 2018).
In this study, the effects of P. fluorescens on gerbera cut flower quality were examined, the optimal antibacterial vase solution concentration of PAA was determined, and the impact of PAA on extending the vase life was assessed.
Materials and Methods
Plant material and vase solution
Gerbera hybrida ‘Lemon Lime’ was transported dry to the laboratory within an hour. Gerberas were recut to 30 cm in length. They were maintained at 22.6 ± 1°C, with a relative humidity of 42.5 ± 6% and exposed to light at 10.7 µmol·m-2 s-1 (12 hours) in a growth chamber (VS-91 G09 M-1300, Visionbionex, Bucheon, Korea). Flowers were placed in a 500 mL glass bottle containing 300 mL of a vase solution (Fanourakis et al. 2013). Ten cut flowers were assigned to each treatment in a completely randomized design. The vase solutions consisted of distilled water (DW), P. fluorescens (P·f) at OD600 0.1 (1 × 108 CFU·L-1), P·f combined with peracetic acid (PAA) at 0.2 mL·L-1 (P·f + PAA), or P·f with Floralife (Smithers-Oasis, Walterboro, USA) at 10 mL·L-1 (P·f + F·L).
Number of bacteria and identification of microorganisms in the preservative solutions of cut gerbera flowers in a greenhouse
A preservative solution in which cut flowers of ‘Lemon Lime’ were stored at 6°C low-temperature storage for 24 hours after harvesting at a gerbera farm in Gwangju, Korea was prepared. The solution was dispensed onto a PDA (potato dextrose agar, Millipore, India) medium and a beef extract (MBcell, Seoul, Korea) medium at 1 mL in each case and cultured in the dark at 36°C for 48 hours (Sapkota 2022). Additionally, P. fluorescens was cultured under identical conditions in only the beef extract medium. Among the microorganisms cultured in the PDA and beef extract medium, two species showing proliferation rates and patterns most similar to P. fluorescens were selected. Colonies were collected, and pure cultures were isolated on the beef extract medium (MFDS 2022). Bacteria were identified through 16S rRNA gene sequencing and a phylogenetic tree analysis at the KCCM (Korea Microbiological Conservation Center, Seoul, Korea).
Antimicrobial proper treatment concentration of peracetic acid against Pseudomonas fluorescens
The microorganism number 12208 P. fluorescens (IFO 13922) was obtained as a freeze-dried ampoule from the KCCM. The cells were suspended in 0.5 mL of sterile physiological saline (0.85% NaCl) and incubated in beef extract plate medium at 26°C for 48 hours. After collecting only bacteria using a cell scraper, the suspension was diluted with sterile distilled water and measured using an optical plate reader (ABS Plus Microplate Reader, Molecular Devices, San Jose, USA) at 600 nm (Griffiths et al. 2011).
In the paper disc analysis, a suspension with an absorbance at 600 nm was prepared consisting of P. fluorescens and was inoculated with 100 µL of beef extract. Peracetic acid (PAA, Orion New Material, Hwaseong, Korea) was diluted to 5, 10, 15, and 20 mL·L-1 using an 85% PAA stock solution, dispensed at 20 µL onto paper discs, and incubated at 26°C for 24 hours. For the in vitro analysis, 1 mL of an OD600 (optical density at 600 nm) 1.0 suspension was prepared with sterile distilled water from P. fluorescens cultured in beef extract. To this suspension, 1 mL of distilled water or PAA at 0.05, 0.1, 0.2, and 0.3 mL·L-1 was added, and OD600 change values were measured (Widdel 2010).
Quality analysis of cut gerbera flowers
Vase life was determined when its ornamental value decreased due to scape bending, petal curling, or petal wilting (Nakamura and Matsushima 2024). Relative fresh weight (%) and vase solution uptake (mL·d-1·g-1) were measured every day and calculated every 2 d using the formula devised by Chamani and Esmaeilpour (2007).
The average flower diameter was calculated by measuring the flower diameter in both the horizontal and vertical directions using a vernier caliper. The ratio of unopened disk florets to the total flower diameter was expressed as a percentage of the head inflorescence diameter. The color of the ray florets was measured using a color spectrometer (CR-400, Minolta, Tokyo, Japan) at three random spots. The L, a, and b values were recorded. The △E, chroma C, and hue angle H values were calculated to compare flower color at harvest and at the end of vase life (McGuire 1992).
A 20-minute dark-treated head inflorescence was cut. The maximum PSII quantum yield was measured using FluorCam 10 (chlorophyll fluorescence imaging, PSI, Drasov, Drasov, Czech Republic; Lee and Kim 2018). The light conditions were set to 1880 µmol·m-2s-1, with a measurement capacity up to 4000 µmol·m-2s-1. The Fv/Fm value of the photochemical reaction was measured by automatically setting the shutter and sensitivity to the background light level using the Quenching Act 2 protocol. Scape bending was assessed by measuring the bending angle at a point 5 cm below the receptacle throughout vase life (Chen et al. 2020). Browning of the scape refers to the continuous browning length, whereas maximum scape browning refers to the maximum length of the irregular browning pattern at the point of vase life termination. For scape firmness, the maximum stress (N) was measured by cutting a 10 cm section under the receptacle into 3 cm pieces and using a 5 mm probe set to 100 mm/min on a hardness meter (FHM-5, Takemura, Tokyo, Japan).
Pseudomonas fluorescens antibacterial properties of peracetic acid in vase solutions
The OD600 value of the P. fluorescens suspension for each treatment was dispensed into each well of a 96-well microplate at 200 µL using a microplate reader. To determine the bacterial count, the vase solution was serially diluted to a range of 101–104 CFU·mL-1. One milliliter of each dilution was inoculated onto 3M PetrifilmTM (3M Company, St. Paul, MN, USA) and incubated at 35°C for 48 hours (3M Company 2014).
Experimental design and data analysis
The experiment was conducted using a completely randomized block design (n = 10). The data were analyzed by a one-way ANOVA to determine significant differences. An analysis of variance and Duncan’s new multiple range test (DMRT) were performed as post-hoc analyses using SAS software (Statistical Analysis System, version 9.4, SAS Institute Inc., Cary, NC, USA). Differences were considered statistically significant at p < 0.05.
Results
The bacterial counts in the preservative solution stored for 24 hours immediately following harvest at a commercial gerbera farm confirmed the presence of microbial contamination (Supplementary Table S1). A phylogenetic tree based on 16S rRNA sequences showed that Pseudomonas spp. were isolated (Fig. 1A and 1B). Isolated from the gerbera preservative solution, these bacteria closely clustered with P. sichuanensis and Lysobacter enzymogenes, supported by an 89% bootstrap value, indicating high genetic similarity exceeding 99% at the species level. These results indicate that bacteria associated with scape bending in gerbera are closely related to Pseudomonas spp. and that similar bacterial genera are also present in the water used in commercial cut gerbera farms.
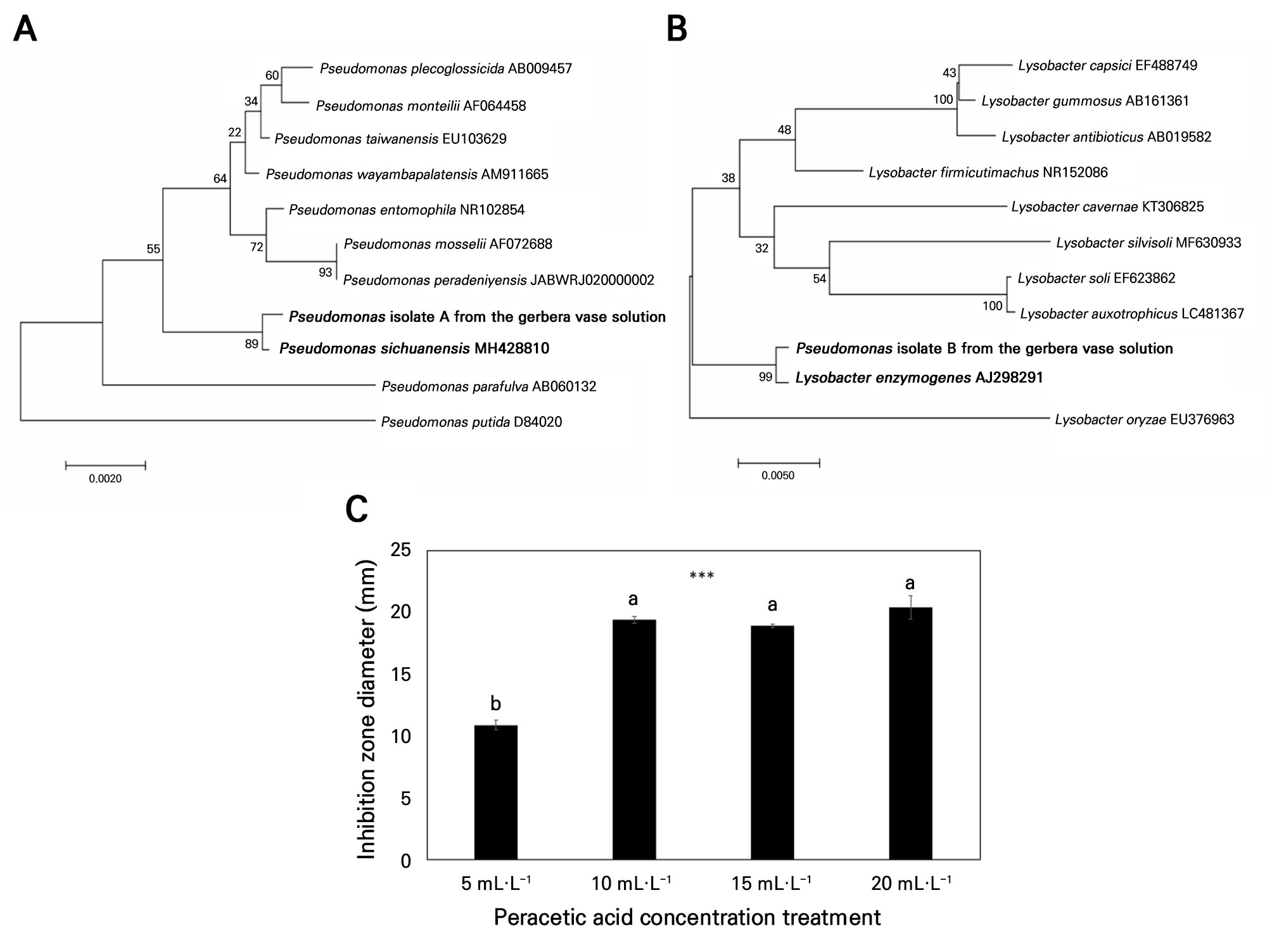
Fig. 1.
Results of phylogenetic analyses of isolates: (A) and (B) (Pseudomonas-like) from gerbera preservative based on 16S rRNA gene sequencing. The neighbor-joining tree was constructed using MEGA software. (C) Antibacterial effects of the peracetic acid at different concentrations on the Pseudomonas fluorescens inhibition zone diameter. Vertical bars denote standard errors (n = 3). Data were analyzed by means of a one-way ANOVA followed by Duncan’s multiple range test (C) (p < 0.05).
PAA exhibited clear antibacterial activity against P. fluorescens, as demonstrated by inhibition zones in the disc diffusion assay (Fig. 1C). When P. fluorescens suspensions were treated with PAA, the OD600 values of DW decreased after two hours, but all PAA treatments exhibited significantly lower OD600 values compared to DW (Table 1). These results confirmed the antibacterial effect of PAA against P. fluorescens. The minimum effective concentration was 0.05 mL·L-1, which significantly reduced OD600 compared with DW, and the treatment with 0.3 mL·L-1 PAA resulted in approximately 42% lower OD600 values compared to DW (Table 1). These results demonstrate the antibacterial activity of PAA against P·f, a major causal agent of scape bending, and confirm both its potential application as a vase-life-extending agent and the effective treatment concentration.
Table 1.
Antibacterial effects of a peracetic acid treatment on a Pseudomonas fluorescens suspension with an optical density (OD600) of 1.0, as measured after the treatments, compared to distilled water (DW)
| Peracetic acid treatment | Optical density (OD600) | ||
| After 10 seconds | After 1 hour | After 2 hours | |
| DW | 0.56 az | 0.56 a | 0.52 a |
| PAA 0.05 mL·L-1 | 0.47 b | 0.39 b | 0.34 b |
| PAA 0.1 mL·L-1 | 0.46 b | 0.37 b | 0.32 b |
| PAA 0.2 mL·L-1 | 0.45 b | 0.36 b | 0.31 b |
| PAA 0.3 mL·L-1 | 0.45 b | 0.35 b | 0.30 b |
| F-value | 4.72 | 23.02 | 20.77 |
| P-value | 0.021* | < 0.001*** | < 0.001*** |
Following treatment with PAA as a vase solution, the vase life of cut gerbera increased by 26.2% and 32.1% compared to DW when treated with P·f + PAA and P·f + F·L, respectively (Fig. 2A). The first signs of vase life termination appeared in the form of scape bending (Fig. 2B). Petal curling started on day 6, whereas petal wilting was observed last on day 13. Scape bending of termination symptoms was highest in P·f treatment, correlating with the shortest vase life. However, the addition of PAA and F·L reduced scape bending to 20% and 10%, respectively, resulting in longer vase life (Fig. 2). Therefore, consistent with previous studies, P. fluorescens was found to induce scape bending in gerbera stems.
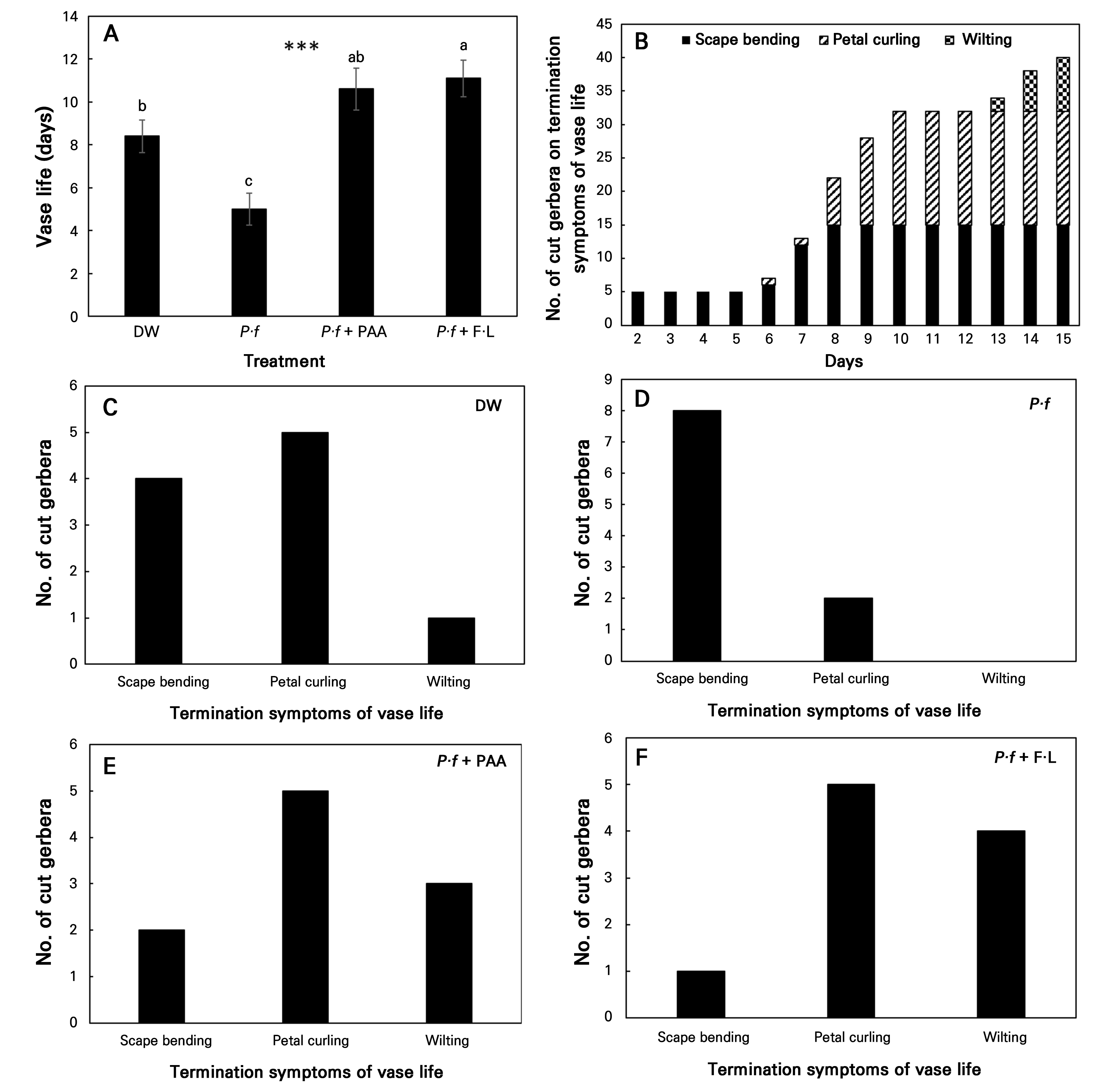
Fig. 2.
Effects of vase solution treatments on (A) vase life and (B) the number of cut flowers at the termination of vase life in cut gerbera flowers: treatments consisted of distilled water (DW), 1 × 108 CFU·L-1Pseudomonas fluorescens (P·f), Pseudomonas fluorescens with 0.2 mL·L-1 peracetic acid (P·f + PAA), and Pseudomonas fluorescens with 10 mL·L-1 Floralife (P·f + F·L). (C)–(F) show the number of cut gerbera flowers at the vase life termination point for each treatment. Vertical bars denote standard errors (n = 10). Data were analyzed by means of a one-way ANOVA followed by Duncan’s multiple range test (A) (p < 0.05).
The relative fresh weight of gerbera increased to 105% in both DW and P·f + PAA on day 1 (Fig. 3A). The relative fresh weight in DW and P·f + PAA remained above 100% until two days after harvest. In contrast, when using P·f treatment, it was already reduced to 94% immediately following harvest. The water uptake rate was lowest after the P·f treatment on day 1 after treatment and decreased further by the end of vase life (Fig. 3B). In contrast, the water uptake rates in P·f + PAA and P·f + F·L were higher than those in DW. Accordingly, the P·f + PAA treatment maintained both the relative fresh weight and water uptake in cut gerbera.
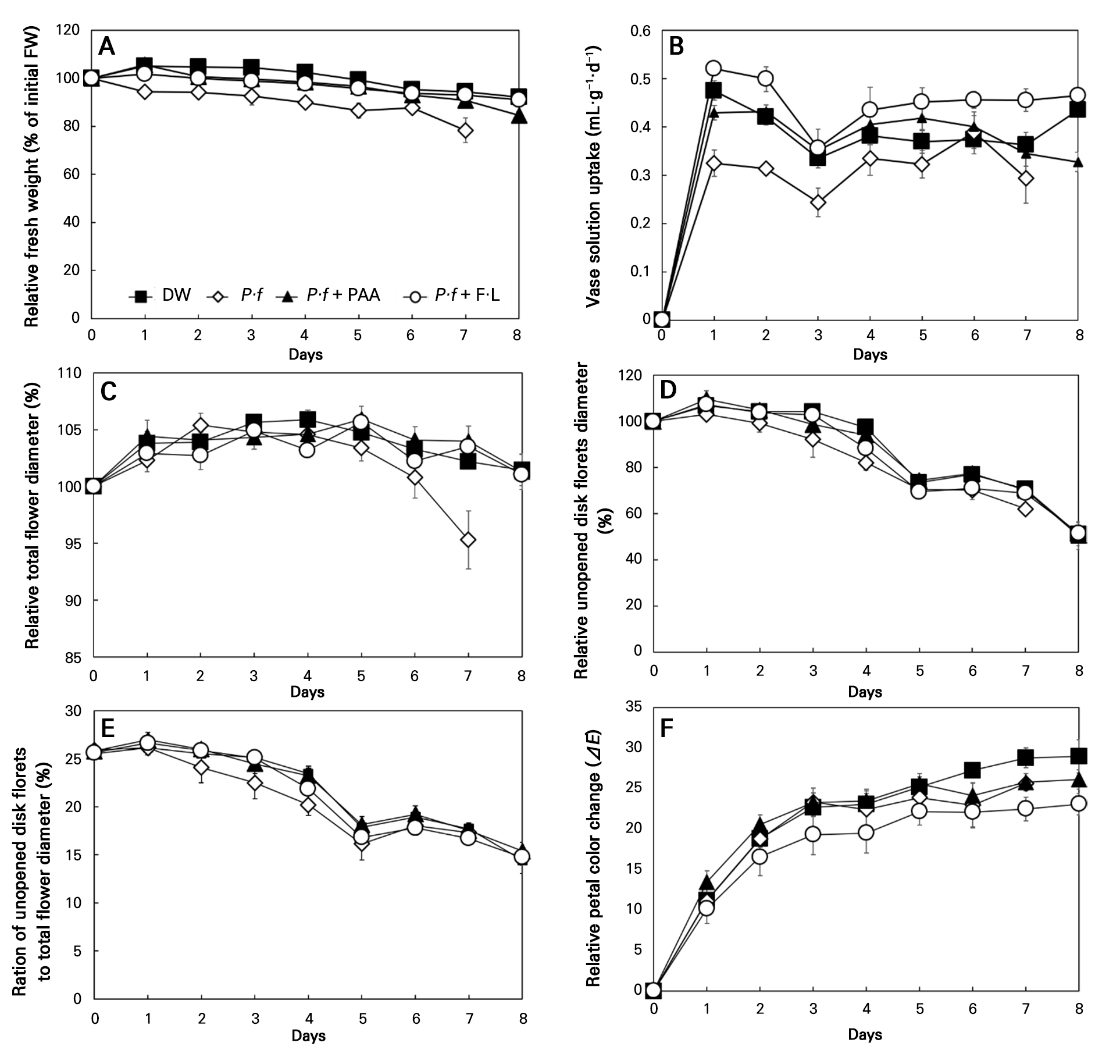
Fig. 3.
Effects of vase solution treatments on (A) the relative fresh weight, (B) vase solution uptake, (C) relative total flower diameter, (D) relative unopened disk floret diameter, (E) ratio of unopened disk florets to the total flower diameter, and (F) relative petal color change in cut gerbera flowers: treatments consisted of distilled water (DW), 1 × 108 CFU·L-1Pseudomonas fluorescens (P·f), Pseudomonas fluorescens with 0.2 mL·L-1 peracetic acid (P·f + PAA), and Pseudomonas fluorescens with 10 mL·L-1 Floralife (P·f + F·L). Vertical bars denote standard errors (n = 10).
After the P·f treatment, the relative total flower diameter declined sharply from five days after harvest compared to the initial value (Fig. 3C). At harvest, the ratio of unopened disk florets to the total flower diameter was 26% on average, whereas after eight days, this ratio decreased to 17% for the P·f treatment, 14% in the DW case, and 15% for both P·f + PAA and P·f + F·L (Fig. 3D and 3E). As the unopened disk florets decreased in diameter over time, the ratio of the unopened disk floret diameter to the total flower diameter also declined across all treatments, decreasing from about 25% to as low as 15% (Fig. 3C–3E). The relative petal color change was most noticeable in DW, whereas it was least noticeable in P·f + F·L, with no significant difference observed between P·f + F·L and P·f + PAA (Fig. 3F).
Chlorophyll fluorescence (Fv/Fm) was measured in disk florets rather than ray florets (Fig. 4B). The Fv/Fm values of disk florets were higher in the P·f + F·L and P·f + PAA cases compared to the DW and P·f treatment (Fig. 4A). Scape bending in gerbera was greater after the P·f treatment compared to the corresponding findings after the other treatments (Table 2). Scape firmness was highest in the P·f + PAA case, whereas it was lowest after the P·f treatment. On day 5, the vase life of 50% of the flowers in the P·f treatment case had terminated due to scape bending, whereas no termination caused by scape bending was observed in the other treatments at this stage (Table 2 and Fig. 2). PAA effectively reduced scape bending and maintained the highest scape firmness, thereby minimizing physical damage to the scapes caused by the P·f treatment.
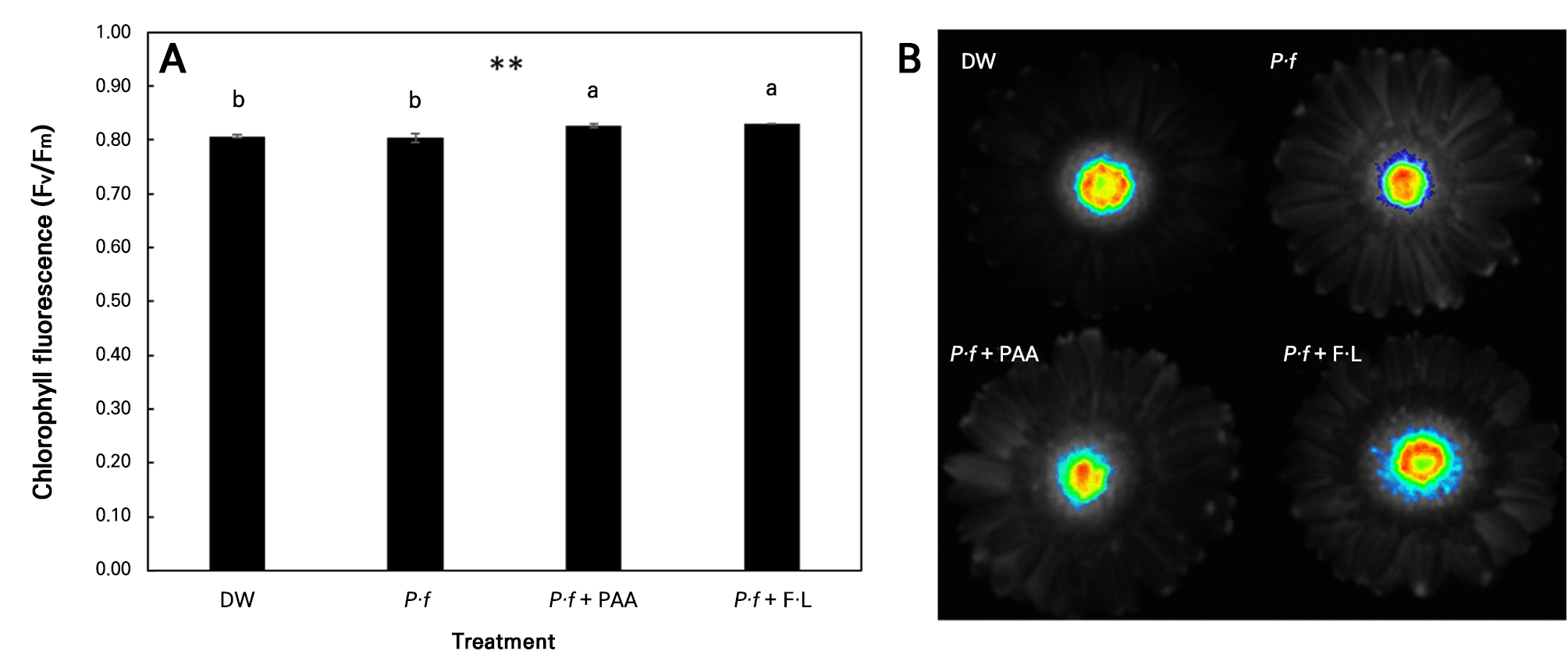
Fig. 4.
Effects of vase solution treatments on (A) chlorophyll fluorescence, and (B) an image of disk florets: treatments consisted of distilled water (DW), 1 × 108 CFU·L-1Pseudomonas fluorescens (P·f), Pseudomonas fluorescens with 0.2 mL·L-1 peracetic acid (P·f + PAA), and Pseudomonas fluorescens with 10 mL·L-1 Floralife (P·f + F·L). Vertical bars denote standard errors (n = 3). Data were analyzed by means of a one-way ANOVA followed by Duncan’s multiple range test (A) (p < 0.05).
Table 2.
Effects of vase solution treatments of distilled water (DW), Pseudomonas fluorescens (P·f), Pseudomonas fluorescens with peracetic acid 0.2 mL·L-1 (P·f + PAA), and Pseudomonas fluorescens with Floralife 10 mL·L-1 (P·f + F·L) on scape bending, scape firmness, terminated vase life due to scape bending, optical density, and bacterial count on day 5 in cut gerbera flowers
| Treatment |
Scape bending (°) |
Scape firmness (N) |
Terminated vase life due to scape bending (%) | Optical density (OD600) | Bacterial count (CFU·mL-1) |
| DW | 6.20 bz | 12.20 ab | 0 | 0.03 c | 3.53 × 105 c |
| P·f y | 65.00 a | 9.66 b | 50 | 0.11 a | 1.77 × 106 b |
| P·f + PAA | 0.70 b | 15.97 a | 0 | 0.06 b | 2.00 × 104 d |
| P·f + F·L | 9.00 b | 15.25 ab | 0 | 0.11 a | 2.75 × 106 a |
| F-value | 56.65 | 5.38 | - | 519.05 | 3179.07 |
| P-value | < 0.001*** | 0.046* | - | < 0.001*** | < 0.001*** |
Browning from the base did not occur after the DW or P·f treatment but was observed in the P·f + PAA and P·f + F·L cases (Fig. 5A). Scape elongation was most significant in DW, whereas no elongation occurred after the P·f treatment (Fig. 5B). In the P·f + PAA and P·f + F·L cases, it was 52.5% and 65.0% lower than in DW, respectively. Observation of cut sections beneath the receptacle after the vase solution treatments showed that cavity formation expanded quickly after the P·f treatment (Fig. 5C). However, no cavity formation was observed when PAA and F·L were applied together with the P·f treatment. Therefore, although scape browning increased in the P·f + PAA and P·f + F·L cases, cavity formation was suppressed, contributing to the maintenance of water conductivity.
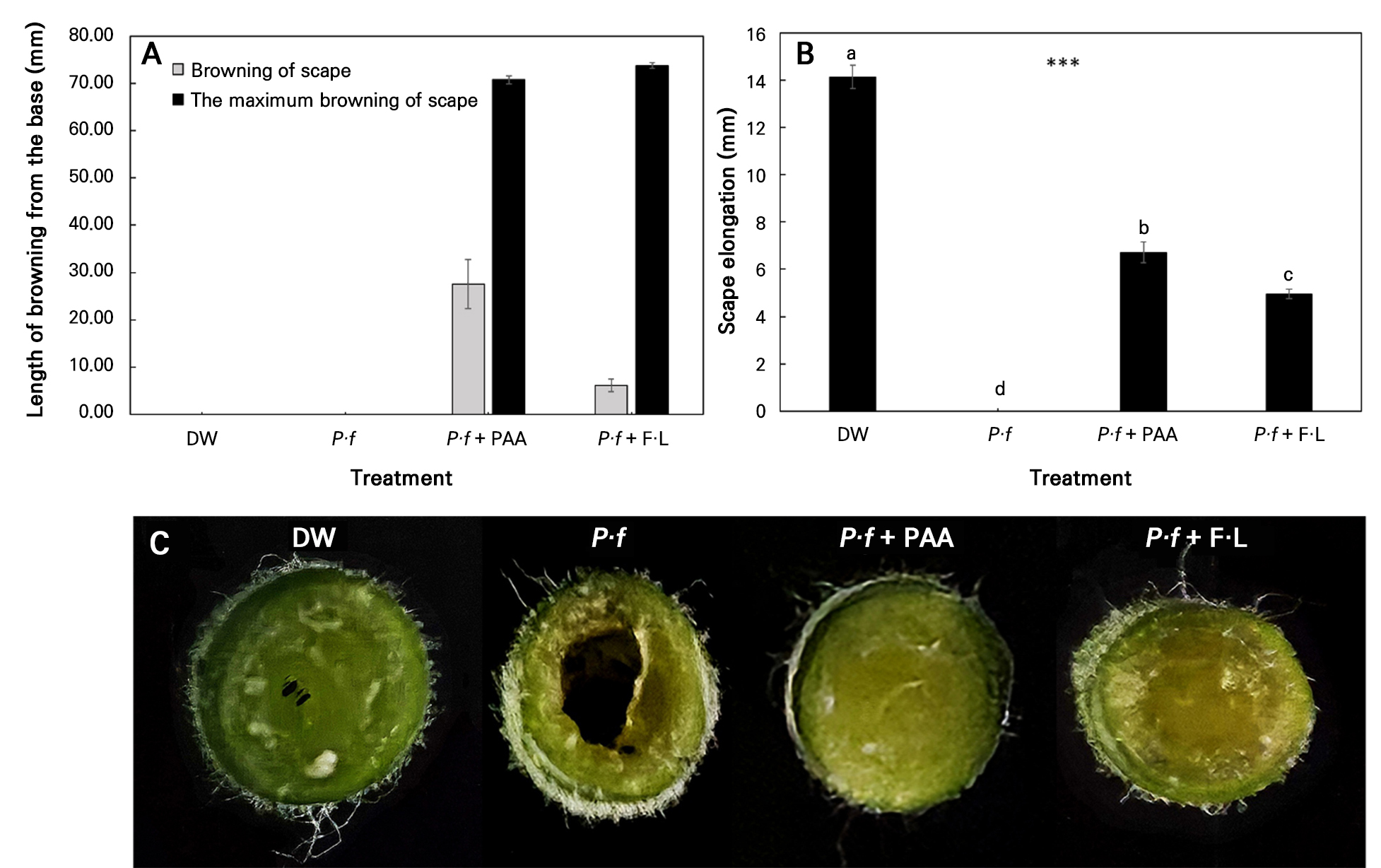
Fig. 5.
Cut gerbera flowers subjected to different treatments: distilled water (DW), 1 × 108 CFU·L-1Pseudomonas fluorescens (P·f), Pseudomonas fluorescens with peracetic acid 0.2 mL·L-1 (P·f + PAA), and Pseudomonas fluorescens with Floralife 10 mL·L-1 (P·f + F·L): (A) browning of scape and maximum browning of scape, (B) scape elongation, and (C) appearance of the cut surface of the scape on day 5. Vertical bars denote standard errors (n = 10). Data were analyzed by means of a one-way ANOVA followed by Duncan’s multiple range test (B) (p < 0.05).
In DW, the OD600 value of the vase solution was the lowest; however, when cultured on a medium, bacterial counts reached a relatively high level (Table 2). In the P·f and P·f + F·L cases, OD600 values were similar to the initial values on day 1. In contrast, the P·f + PAA treatment showed a reduced OD600 value, confirming the antibacterial activity of PAA against the P·f treatment. Bacterial counts were highest in the P·f + F·L case, whereas P·f + PAA exhibited the lowest bacterial count. This represents an approximately 99% reduction compared with the P·f treatment, clearly demonstrating the antibacterial effect of PAA in the vase solution.
Discussion
Vase life was shortened more rapidly when termination occurred due to scape bending than when it was caused by petal curling or wilting (Fig. 2). This was consistent with previous reports (Tonooka et al. 2024). Gerbera generally had a shorter vase life when scape bending occurred, whereas the vase life of flowers without scape bending was maintained for more than six days. Several studies have demonstrated that the vase life of cut gerbera is strongly influenced by water transport characteristics and vascular blockages related to scape bending. Microorganisms rapidly multiply within the stem immediately following harvest, blocking water transport and shortening the vase life (Put 1990). Scape bending in gerbera is more closely linked to water stress and endogenous abscisic acid (ABA) regulation than to ethylene (Ge et al. 2019). Analyses of differentially expressed genes (DEGs) involved in hormone signaling pathways, along with measurements of ABA, suggest that stem bending may be an ethylene-independent process and one controlled by ABA. In this study, scape bending is influenced not only by bacterial abundance in the vase solution but also by the ability of the plant to maintain vascular function (Table 2 and Fig. 5C).
Pseudomonas spp. are major bacteria that cause vascular occlusion in cut flowers. In cut carnations, this species accounts for 50% of the bacterial population in stems and vase solutions at ten days. A previous study found that when the concentration of P. aeruginosa exceeded 108 CFU·mL-1 in the vase solution of cut gerbera, scape bending exceeded 90°, leading to the end of vase life (van Doorn and de Witte 1994; van Doorn et al. 1995). In cut roses, the concentration of P. fluorescens in the vase solution was found to be significantly correlated with vase life (p = 0.0009) (Robinson et al. 2015).
In cut gerbera, Bacillus and Pseudomonas spp. in the vase solution adheres to the cut stem surface, disrupting water uptake and acting as a filter that restricts bacterial movement into the xylem (Put 1990). Other microorganisms, such as Fusarium oxysporum and Kluyveromyces marxianus, are absorbed into xylem cells. Therefore, the specific bacterial species, rather than the total bacterial population in the vase solution, plays a crucial role in inhibiting water uptake. In this study, cut flowers treated with P. fluorescens also showed a significant decline in water uptake over time (Fig. 3B).
The bacterial count in the vase solution of commercial gerbera farms is reportedly 8.40 × 103 CFU·mL-1, which is a level known to cause a water imbalance due to vascular occlusion in cut flowers (Supplementary Table S1; Li et al. 2012). Isolated from the gerbera preservative solution, these bacteria closely cluster with P. sichuanensis and Lysobacter enzymogenes (Fig. 1A and 1B). Pseudomonas sichuanensis was first isolated from hospital sewage in Chengdu, Sichuan Province, China (Qin et al. 2019). Although it is genetically close to P. putida, which is known to cause vascular occlusion in cut flowers, it has been recognized as a separate species through a 16S rRNA sequence analysis (Put 1990; van Doorn et al. 1991). As a recently described environmental bacterium, research on P. sichuanensis remains limited, meaning that further studies are needed (Qin et al. 2019).
Lysobacter enzymogenes is a gram-negative bacterium commonly found in soil, aquatic environments, and plant rhizospheres (Ramamoorthy et al. 2001; Folman et al. 2003). It can exist as an endophyte or epiphyte and may migrate into the scape from the vase solution or originate from the cultivation environment, such as the soil or irrigation water (Friesen 2011). It is known to have beneficial effects as a biological control agent against fungi, bacteria, and nematodes, with recent studies highlighting its positive role in biological control (Lin et al. 2023). These findings suggest that bacterial succession in vase solutions is highly dynamic and complex, which may explain the appearance of novel bacterial species such as P. sichuanensis and L. enzymogenes in preservative solutions from commercial gerbera farms (Fig. 1A and 1B).
Bacterial counts and OD600 values increased proportionally with the bacterial density (Fig. 1; Griffiths et al. 2011). In vitro, PAA concentrations ≥ 0.05 mL·L-1 showed similar antibacterial activity up to 0.3 mL·L-1 (Table 1). This observation indicates that antibacterial efficacy was maintained across this concentration range under the present experimental conditions. One hour after the PAA treatment, bubble formation was observed in the P. fluorescens suspension (Supplementary Fig. S1). Previous reports suggest that PAA may decompose in aqueous environments to produce hydrogen peroxide and acetic acid, accompanied by a release of oxygen (Janković and Sinadinović-Fišer 2005). Therefore, the observed bubbling may be associated with such decomposition reactions; however, additional chemical analyses would be required to confirm this mechanism.
In this study, treating vase solutions with the P·f treatment, also a common practice in commercial farms, reduced water uptake and vase life, whereas PAA increased both (Figs. 2A and 3B).
The bacterial counts found in the preservative solutions of gerbera farms were similar to those reported by Balestra et al. (2005), who detected 8.6 × 103 CFU·mL-1 in vase solutions and stem tissues immediately after harvest. Bacterial growth in vase solutions prevents lignin deposition, weakens stem firmness, and shortens vase lifetimes (Tonooka et al. 2023). Therefore, the growth of P. fluorescens and its suppression by PAA appear to be closely associated with scape quality, particularly through reduced lignin accumulation and water imbalance, which results in inadequate development of sclerenchyma tissue in the upper scape (Table 2 and Fig. 3; Perik et al. 2012; Tonooka et al. 2023).
Cavity formation inside the scape begins immediately after harvest and continues to expand during vase life. In this study, cavity expansion progressed rapidly after the P·f treatment by day 5 (Fig. 5C; Perik et al. 2014). Browning of the scape, which did not occur in DW, was observed in the P·f + PAA and P·f + F·L cases, likely due to bacterial contamination (Fig. 5A; Chen et al. 2023).
When bacterial growth in vase solutions is controlled by disinfectants, lignin deposition in scape cells increases, stem firmness improves, scape bending decreases, and vase life increases (Tonooka et al. 2023). In this experiment, the highest scape firmness and the lowest amounts of scape bending were observed after the P·f + PAA treatment (Table 2). The antibacterial effect of PAA against P. fluorescens in vase solutions and the bacterial count measurements confirm that PAA directly enhanced the scape quality.
The cause of stem browning in P·f + PAA may be the oxidative action of PAA, which converts phenolic compounds in plant cells into quinones. Another possibility is cell necrosis resulting from lipid peroxidation of membranes and protein denaturation from antibacterial agents in both the P·f + PAA and P·f + F·L cases, leading to tissue browning (Fig. 5A; Petrov et al. 2015). As a powerful oxidant, PAA plays a key role in plant tissue browning by oxidizing phenolic compounds to quinones (Mayer 2006). Reactive oxygen species (ROS) can also cause cell necrosis through membrane lipid peroxidation and protein denaturation, resulting in browning of plant tissues (Petrov et al. 2015). Browning at the stem base was also observed in the P·f + F·L case (Fig. 5). When Floralife PRG was used as a vase solution for gerbera, it more than doubled the vase life and reduced scape bending. However, FloraLife® Express 300 extended vase life most effectively for roses at a 100% concentration, whereas higher concentrations exceeding 200% resulted in stem browning (Floralife 2010). Commercial chlorine-based preservatives, such as FloraLife PRG 100 specifically for gerbera, can cause stem browning when applied at excessively high concentrations (GrowerTalks 2024). Stem browning in cut flowers can result from various factors, including the chlorine solutions and surfactants used, as well as a low pH and certain microorganisms (Chrysal 2025). Although many commercial preservatives can extend vase lifetimes, stem browning often occurs, and developing a technology to extend the vase life without causing stem browning remains a challenge. However, in herbaceous cut flowers, stem browning may also occur as part of natural floral senescence. Because the stem tissue stays structurally intact, browning does not necessarily shorten the vase life (Chrysal 2025). However, further investigations of lower PAA concentrations would be valuable to assess whether stem browning can be minimized without compromising antibacterial efficacy. In cut gerbera, scape elongation mainly occurs in the upper region during the first two days of vase life, with 10–15 mm of elongation observed in the upper 10 cm section (Perik et al. 2012). However, this elongation does not directly correlate with scape bending. Instead, scape bending results from insufficient development of sclerenchyma tissue in the upper stem or reduced lignin content caused by water loss, which decreases the structural support of the stem. Scape elongation is influenced by a complex interplay involving water uptake, cell wall strengthening, lignification, plant hormones, and water stress (Perik et al. 2012; Ge et al. 2019; Tonooka et al. 2023). In this study, stem elongation was highest in the DW case; however, this elongation did not significantly contribute to an increase in scape bending (Table 2 and Fig. 5B).
The head of a gerbera resembles a solitary flower but comprises three types of florets (ray, trans, and disk) attached to a receptacle and surrounded by bracts. The disk florets are arranged in a spiral phyllotactic pattern, with parastichies (Zhang et al. 2021). Harvesting occurs when the outer two to three rows of disc florets are perpendicular to the stalk. Following harvest, senescence coincides with the sequential opening of disc florets, marking the beginning of the wilting stage (Danziger 2025; TNAU Agritech Portal Horticulture 2025). In all treatments, as the disc florets started to open, the diameter of the unopened disc florets decreased and the ratio of unopened disc florets to total flower diameter was reduced (Fig. 3D and 3E). The Fv/Fm values of the disc florets stayed within the normal range (0.78–0.84) across all treatments. However, values were 0.81 in DW and 0.80 in the P·f treatment, whereas P·f + PAA and P·f + F·L maintained significantly higher values of 0.83 (Fig. 4A; Björkman and Demmig 1987). These results suggest that the disc florets in the P·f + PAA and P·f + F·L cases experienced less water stress compared to those in the DW and P·f treatments (Fig. 3B; Björkman and Demmig 1987). A decrease in Fv/Fm during fruit ripening has been reported in the calyx tissue of persimmon, suggesting that the PSII efficiency in chlorophyll-containing floral organs may be associated with physiological changes during development and senescence (Fathi-Najafabadi et al. 2021). In addition, chlorophyll has been identified in chrysanthemum inflorescences, including disc florets, indicating that chlorophyll fluorescence parameters may serve as supplementary indicators of physiological status in green floral tissues (Fu et al. 2021). Although chlorophyll contents in the disc florets were not directly quantified in the present study, the measured Fv/Fm values may be interpreted as reflecting relative differences in physiological stress levels among the treatments.
Consequently, PAA extended vase life by approximately 26% compared with DW and by 112% compared with the P·f treatment, exhibiting a quality preservation effect comparable to that of the commercial preservative F·L. Furthermore, PAA suppressed cavity formation, stem bending, and stem elongation while also slowing the opening rate of disk florets, thereby contributing to the maintenance of external floral quality.
Supplementary Material
Supplementary materials are available at Horticultural Science and Technology website (https://www.hst-j.org).
- HORT_20260013_Table_S1.jpg
Supplementary Table S1. Bacterial counts in the preservative solution of the cut gerbera farm during storage
- HORT_20260013_Fig_S1.jpg
Supplementary Fig. S1. Appearance of a Pseudomonas fluorescens bacterial suspension (OD600 = 1.0) one hour after treatment with PAA at the following concentrations: A, 0 mL·L-1; B, 0.05 mL·L-1; C, 0.1 mL·L-1; D, 0.2 mL·L-1; and E, 0.3 mL·L-1.
