

Introduction
Materials and Methods
Experimental site
Plant materials
Experimental procedure
Analytical methods
Rooting observation and data collection
Data processing and analysis
Results
Analysis of the root formation process in different types of tea
Analysis of rooting differences in different tea varieties
Changes in the nutrient content during the root formation process
Changes in oxidase activity during root formation
Changes in endogenous hormone levels and ratios
Discussion
Effects of physiological and biochemical indicator changes on the root formation of different tea varieties
Commonalities and differences in the root formation process of medium-leaf and large-leaf tea varieties
Conclusions
Introduction
Tea (Camellia sinensis (L.) O. Ktze.), a member of the family Theaceae and class Dicotyledons, is a widely consumed beverage. It is cultivated mostly in China, India and tropical South Asia. According to the China Tea Circulation Association, the total tea plantation area reached 3.4 million hectares in 2023, an increase of 102,900 hectares from the previous year, increasing the demand for high-quality tea seedlings. Tea plants can reproduce both sexually and asexually. However, problems such as self-incompatibility and trait segregation arise during sexual reproduction, resulting in significant differences in offspring. Therefore, asexual reproduction is often used in production to preserve the superior traits of the mother plant. Cutting propagation has been practiced in China for over 200 years and is popular for its simplicity and efficiency. Tea cutting propagation is primarily through stem cuttings, with the formation of adventitious roots (ARs) being crucial. Tea plants are considered a difficult-to-root species (Wei et al. 2013), and genotype plays a key role in rooting success (Almehdi et al. 2002). Rooting ability varies significantly among different tea varieties, depending on the specific variety due to its genetic and physiological properties. According to the Chinese National Standard (GB 11767-2003), tea plants are classified into large-leaf and medium-leaf varieties based on leaf size, and all superior tea varieties are categorized accordingly (Yang et al. 2003). Most cultivated tea plants belong to the genus Camellia L., section Thea (L.) Dyer, in the family Theaceae and are classified into one of two main varieties: C. sinensis var. sinensis (CSS) and C. sinensis var. assamica (Masters) Chang (CSA) (Wang et al. 2020). Of these, CSS is characterized by smaller leaves, cold tolerance, and a shrub or subshrub growth habit, whereas CSA has larger leaves and an arborous or a subarborescent growth habit (Chen et al. 2000; Yang et al. 2009). Studies have shown that large-leaf varieties generally have a lower root formation rate than medium-leaf varieties (Dong 1984). For example, the root formation rate of ‘Hainan Dayezhong’ is only 22.14%, while the root formation rates of ‘Wuniuzao’ and ‘Fuding Dabai’ are 90.2% and 66.8%, respectively (Wu et al. 2014). Some difficult-to-root varieties require significant labor and resources during the rooting process (Lima et al. 2013; Wei et al. 2014). These differences raise a critical question: Is the classification of large-leaf and medium-leaf varieties related to rooting ability? Understanding this relationship could provide valuable insights into improving propagation efficiency, optimizing resource allocation, and ultimately increasing tea production.
The propagation of tea cuttings is a complex physiological and biochemical process involving interactions among various endogenous hormones, enzyme activities, and nutrients. Recent progress has been made in understanding the physiological and biochemical aspects of tea cuttings. The content of endogenous hormones in cuttings is closely related to the formation of ARs. Typically, endogenous hormones studied in tea cuttings include indole-3-acetic acid (IAA), cytokinins (CTK), and abscisic acid (ABA). These hormones play a crucial role in regulating the distribution of nutrients to the root formation site, increasing the activity of related enzymes, and promoting AR formation (Husen 2011). Changes in endogenous hormone levels during the root formation process vary depending on the tea variety. In addition, polyphenol oxidase (PPO), peroxidase (POD), and indole-3-acetic acid oxidase (IAAO) significantly affect growth, development, root formation, and other metabolic processes in cuttings during propagation. Studies have shown that higher levels of soluble sugars in cuttings can promote callus and AR formation (Rapaka et al. 2005; Tombesi et al. 2015). Furthermore, nitrogen-containing compounds, such as soluble proteins, are also closely related to root formation in cuttings (Rout 2006). Despite extensive research, systematic comparative analyses of tea varieties with different rooting abilities remain limited. In particular, comprehensive studies that integrate various physiological and biochemical indicators remain insufficient.
This study aims to provide a systematic analysis of the changes and interactions among nutrients, key enzymes, and endogenous hormones during the root formation process in different tea varieties. By comparing the physiological and biochemical properties of cutting root formation in six tea varieties with varying root formation abilities, this research seeks to identify the key factors influencing root formation ability. The results are expected to provide new insights into the propagation of tea plants from cuttings.
Materials and Methods
Experimental site
The experimental site was located at a tea plantation of the Hainan Academy of Agricultural Sciences in Haikou, Hainan Province, China (110°22'E, 20°0'N). This region is characterized by a tropical monsoon climate with indistinct seasonal changes, minimal annual temperature variations, and a high annual average temperature.
Plant materials
The test materials consisted of six tea varieties that have significant economic importance for tea production, including three large-leaf varieties: ‘Hainan Dayezhong’ (HD), ‘Yunnan Dayezhong (YK10)’ (YD), ‘Fuan Dabai’ (FA), as well as three medium-leaved varieties: ‘Fuding Dabai’ (FD), ‘Jinxuan’ (JX), and ‘Huang Guanyin’ (HG). HD, YD, and FA are commonly used for black and green tea production due to their high yield potential and good processing quality. FD is widely cultivated to produce high-quality white tea, black tea, and green tea. JX is highly regarded in oolong tea production because of its unique aroma. HG is a hybrid of ‘Tieguanyin’ and ‘Huangdan’ and is known for its strong aroma and disease resistance, making it a preferred variety for oolong tea production. The medium-leaf and large-leaf varieties used were classified based on definitions in an existing catalog of approved tea varieties and on the findings of previous studies (Yang et al. 2003; Wang et al. 2020).
The cuttings were collected from the clonal mother plant garden of the Provincial Tea Germplasm Resource Bank in Baisha once a month from February 20 to April 20 of 2023 (three samples in total). The cuttings were taken from ten mother plants of each variety, the mother plants being five years old. Every October, the trees were heavily pruned 40–50 cm above the ground to stimulate the growth of new shoots. Semi-lignified branches from the current year’s growth were selected for cuttings; these branches were healthy, free of pests and diseases, and had new shoots more than 25 cm long and a trunk diameter of 3–5 mm. The branches were cut into segments about 3 cm long, each containing a healthy axillary bud and a leaf. In this study, a randomized complete block design was adopted, and a total of six tea varieties were included, with three replicates per variety and 150 cuttings per replicate. A total of 2700 cuttings were required.
Experimental procedure
A soil bed measuring 700 cm × 120 cm was prepared in each replicate using yellow heart soil as the cutting substrate. The evening before planting, the soil was thoroughly watered and disinfected with a 200-fold dilution of 30% hymexazol.
Analytical methods
The soluble sugar content was determined using the anthrone colorimetric method (Loewus 1952), while the soluble protein content was measured using the Coomassie brilliant blue method (Asryants 1985). The POD, PPO, and IAAO activities were determined using kits from the Shanghai Xinyu Biotechnology Company (spectrophotometer method), following the instruction manual. Plant hormones, in this case auxin (IAA), cytokinin (CTK), and abscisic acid (ABA), were measured using an enzyme-linked immunosorbent assay (ELISA). The absorbance levels of IAA, CTK, and ABA at 490 nm were measured with an ELISA spectrophotometer. All antibodies against each hormone were monoclonal and were provided by Shanghai Xinyu Biotechnology Co., Ltd. Each measurement was performed in triplicate for all indicators.
Rooting observation and data collection
Root observations were conducted every three days starting from the fifth day after planting. Ten cuttings were randomly selected for examination. The planting day was recorded as S0. Callus tissue formation was noted as S1. The appearance of visible ARs through the cortex was recorded on the day of AR initiation (S2). The day when the number of ARs reached ≥ 3 and the root length reached ≥ 1 cm was recorded as the day of AR proliferation (S3). The callus induction period was designated as P1 (S0–S1), the AR initiation period as P2 (S1–S2), and the AR expression and elongation periods as P3 (S2–S3). Seventy days after planting, the root formation process was completed for each treatment. At this time, 60 cuttings were collected from each group to determine the root formation rate, the number of roots per cutting, and the total root length per cutting. The rooting index was calculated as follows:
Rooting Index = Rooting Rate × Average Root Length × No. of Root per Shoot
Data processing and analysis
The data were processed using Microsoft Excel 2021. Statistical analyses were performed using IBM SPSS 18.0 software (SPSS Inc., Chicago, IL, USA). The experiment followed a completely randomized block design with three biological replicates per variety, and each replicate included 150 cuttings. Multiple comparisons were made using the least significant difference (LSD) method at a significance level of α = 0.05. Charts were created using Microsoft Excel 2021.
Results
Analysis of the root formation process in different types of tea
The data in Table 1 indicate that HD required the longest time to complete the rooting process, taking 69.9 days, while JX required the shortest time, taking only 38.4 days. YD and FA exhibited extended durations during the AR initiation period (P2), taking 24.0 days and 24.9 days, respectively. Significant differences (p < 0.05) were observed among the varieties during the callus induction period (P1) and the AR expression and elongation periods (P3).
Table 1.
Analysis of the root formation process in different tea varieties
| Varieties | Occurrence time of ARs /d | |||
| Callus induction period(P1) | AR initiation period(P2) | AR expression and elongation periods(P3) | P1+P2+P3 | |
| HD | 34.50 ± 0.67 a | 15.60 ± 0.60 c | 19.80 ± 0.49 a | 69.90 ± 1.01 a |
| YD | 16.20 ± 0.49 b | 24.00 ± 0.63 a | 16.50 ± 0.50 b | 56.70 ± 1.04 b |
| FA | 15.30 ± 0.30 b | 24.90 ± 0.64 a | 12.90 ± 0.64 d | 53.10 ± 1.10 c |
| FD | 13.80 ± 0.49 c | 21.90 ± 0.46 b | 15.30 ± 0.54 bc | 51.00 ± 0.77 c |
| JX | 12.60 ± 0.40 c | 11.10 ± 0.46 d | 14.70 ± 0.94 bcd | 38.40 ± 1.47 e |
| HG | 15.30 ± 0.30 b | 12.60 ± 0.60 d | 14.40 ± 0.75 cd | 42.30 ± 1.22 d |
| F | 318.29 | 111.15 | 12.76 | 99.00 |
| P | 0.0001 | 0.0047 | 0.0001 | 0.0001 |
The above statistical data are presented as the mean ± SD of three repetitions for each tea variety: ‘Hainan Dayezhong’ (HD), ‘Yunnan Dayezhong’ (YD), ‘Fuan Dabai’ (FA), ‘Fuding Dabai’ (FD), ‘Jinxuan’ (JX), and ‘Huang Guanyin’ (HG). Different lowercase letters in the same column indicate significant differences at the 5% probability level (p < 0.05).
Analysis of rooting differences in different tea varieties
Through a comparative analysis of the rooting ability of different tea varieties (Fig. 1 and Table 2), it was found that YD had the highest rooting rate of 84.81%, with an average of 21 roots per shoot, while HD had the lowest root formation rate of 22.86%. Among the medium-leaf varieties, JX and FA had significantly higher root formation rates than HG, with root formation rates of 86.3%, 86.11%, and 51.52%, respectively. In terms of the average number of roots, YD, FD, and JX had more roots, with YD having the most at 21 roots. HG, HD, and FA had fewer roots on average. There were no significant differences in the average root length between the tested varieties. YD had the highest root index of 86.27, while HD had the lowest root index of only 8.33.

Table 2.
Rooting performances of different tea varieties
| Varieties | Rooting rate/% | No. of root per shoot | Average root length/cm | Rooting index |
| HD | 22.86 ± 3.35 d | 9.40 ± 1.70 b | 3.88 ± 0.56 ab | 8.33 ± 1.22 d |
| YD | 84.81 ± 2.11 a | 21.10 ± 3.90 a | 4.82 ± 0.44 a | 86.27 ± 2.14 a |
| FA | 69.41 ± 0.50 b | 9.10 ± 0.82 b | 3.63 ± 0.28 ab | 22.93 ± 0.16 c |
| FD | 86.11 ± 4.00 ab | 13.30 ± 1.63 ab | 3.02 ± 0.37 b | 34.56 ± 1.61 b |
| JX | 86.30 ± 1.54 ab | 16.00 ± 2.03 ab | 2.87 ± 0.12 b | 39.64 ± 0.71 b |
| HG | 51.52 ± 4.30 c | 9.70 ± 1.09 b | 2.56 ± 0.24 b | 12.77 ± 1.06 d |
| F | 66.28 | 5.10 | 5.19 | 458.15 |
| P | 0.0001 | 0.0007 | 0.0006 | 0.0001 |
The above statistical data are presented as the mean ± SD of three repetitions for each tea variety: ‘Hainan Dayezhong’ (HD), ‘Yunnan Dayezhong’ (YD), ‘Fuan Dabai’ (FA), ‘Fuding Dabai’ (FD), ‘Jinxuan’ (JX), and ‘Huang Guanyin’ (HG). Different lowercase letters in the same column indicate significant differences at the 5% probability level (p < 0.05).
Changes in the nutrient content during the root formation process
(1) Changes in the soluble sugar content: changes in the soluble sugar content among the different tea varieties mainly exhibited two trends (Fig. 2A). JX showed a stable increase from the initial to the final stage of the cutting process, with its soluble sugar content increasing from 147.54 mg·g-1 to 207.50 mg·g-1. In contrast, YD, HD, FA, FD, and HG showed an initial decrease followed by an increase in the soluble sugar content. YD initially decreased but then increased significantly to 216.09 mg·g-1. Although HD experienced similar fluctuations, its final content was lower. FA, FD, and HG exhibited similar trends but differed in terms of the recovery ability and final content.
(2) Changes in the soluble protein content: at different rooting stages, the rooting ability of a variety influences the trends in its soluble protein content (Fig. 2B). Varieties with strong rooting ability, such as YD and JX, exhibited a significant increase in the soluble protein content during the AR expression and elongation periods. During the P3 period, the soluble protein content in YD increased to 13.66 mg·g-1, while for JX it increased to 14.60 mg·g-1. In contrast, varieties with weaker root formation abilities, such as HD, showed a more significant decrease in the soluble protein content during the callus formation induction period and AR initiation period. Furthermore, their soluble protein contents increased less during the AR expression and elongation periods.

Fig. 2.
Changes in the nutrient content during the root formation process of different tea plant varieties: soluble sugar content (A) and soluble protein content (B). Different capital letters indicate significant differences among different varieties at the same period (p < 0.05), while different lowercase letters indicate significant differences within the same variety over different periods (p < 0.05).
Changes in oxidase activity during root formation
(1) PPO: As shown in Fig. 3A, during the cutting process of the six tea varieties, HD initially exhibited high PPO activity of 110.32 IU·L-1, which fluctuated throughout the rooting process and peaked at 140.55 IU·L-1 at 60 days. YD also demonstrated relatively high initial PPO activity of 117.16 IU·L-1, but showed a general downward trend in the form of a 15% decrease at 60 days from the initial value. FA exhibited the highest initial PPO activity of 122.59 IU·L-1, peaking at 139.37 IU·L-1 at 50 days before stabilizing at 129.72 IU·L-1 at 60 days. FD had lower PPO activity overall, starting at 90.96 IU·L-1, peaking at 111.63 IU·L-1 at 30 days, and decreasing to 83.73 IU·L-1 at 60 days, showing an 8% decrease from the initial level. JX started with a PPO activity level of 100.88 IU·L-1, which increased steadily to 123.40 IU·L-1 at 50 days before declining slightly to 106.48 IU·L-1 at 60 days. HG started with the lowest PPO activity, peaking after ten days before gradually decreasing. HG exhibited the lowest initial PPO activity of 83.95 IU·L-1, peaking at 122.65 IU·L-1 at ten days, then fluctuating, and finally reaching 124.59 IU·L-1 at 60 days, showing a 48.4% increase compared to its initial value.
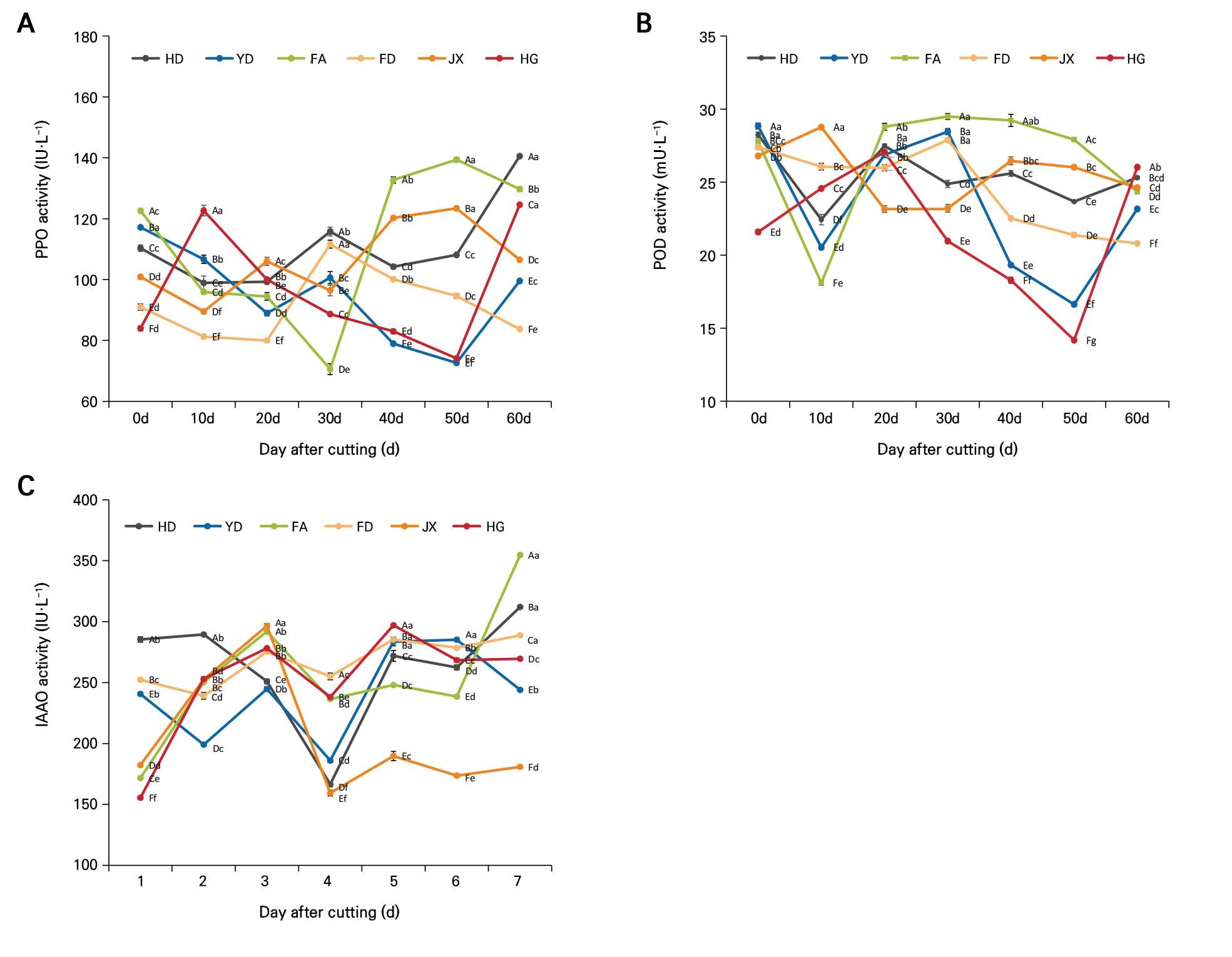
Fig. 3.
Changes in oxidase activity levels during the rooting process of different tea plant varieties: PPO activity (A), POD activity (B), and IAAO activity (C). Different capital letters indicate significant differences among different varieties at the same period (p < 0.05), while different lowercase letters indicate significant differences within the same variety over different periods (p < 0.05).
(2) POD: Fig. 3B shows that HD initially had a POD activity level of 28.24 mU·L-1, which decreased to 22.44 mU·L-1 at ten days, increased to 27.49 mU·L-1 at 20 days, and then became stabilized at 25.31 mU·L-1 at 60 days. YD started with a POD activity level of 28.84 mU·L-1, which fell sharply to 20.53 mU·L-1 at ten days, reached a peak of 28.46 mU·L-1 at 30 days, and then declined steadily to 23.16 mU·L-1 by 60 days. FA showed initial POD activity of 27.79 mU·L-1, which decreased to 18.10 mU·L-1 after ten days with a reduction of 34.9% and then peaked at 29.49 mU·L-1 after 30 days before falling to 24.34 mU·L-1 at 60 days. FD showed relatively stable POD activity starting at 27.40 mU·L-1, with a slight peak of 27.88 mU·L-1 after 30 days and a decrease to 20.80 mU·L-1 after 60 days. JX started at 26.79 mU·L-1 and peaked at 28.76 mU·L-1 after ten days before settling at approximately 26.02 mU·L-1 after 50 days. HG had the lowest initial POD activity at 21.59 mU·L-1, which increased to 27.05 mU·L-1 at 20 days and then fluctuated and sharply decreased to 14.18 mU·L-1 at 50 days. After 60 days, it recovered to 26.02 mU·L-1, an increase of 20.5% compared to its initial value.
(3) IAAO: The IAAO activity exhibited significant differences among the tea varieties tested here (Fig. 3C). HD had the highest initial IAAO activity at 285.39 IU·L-1, followed by FD and YD, while HG had the lowest initial IAAO activity. In terms of trends, HD reached its IAAO activity peak of 289.44 IU·L-1 at ten days and then began to decline, reaching a trough of 166.57 IU·L-1 at 30 days, followed by an increase. YD showed a rapid decrease in IAAO activity after cutting, reaching its first trough of 199.07 IU·L-1 at ten days, followed by an increase and then another decrease at 30 days to 186.04 IU·L-1 before a gradual rise again. FA displayed a gradual increase in IAAO activity after cutting, peaking at 20 days with a value of 291.90 IU·L-1, followed by fluctuating increases. FD maintained relatively stable IAAO activity throughout the rooting stage, starting at 252.36 IU·L-1 and ending at 288.78 IU·L-1 at 60 days, showing a steady increase. JX experienced dramatic changes in IAAO activity, with a slight increase during the induction period followed by sharp declines during the initiation and elongation stages, reaching a trough at 30 days with a value of 159.40 IU·L-1 and then quickly fluctuating upwards. HG had the lowest initial IAAO activity, which rapidly increased after cutting, peaking at 20 days at 278.25 IU·L-1, with fluctuating activity then ensuing.
Changes in endogenous hormone levels and ratios
(1) IAA: The IAA content changes among the different tea varieties (Fig. 4A) reveal significant differences in how the endogenous hormone levels are adjusted in each variety. YD initially showed a high IAA content at 77.58 µg·L-1, peaking early at 84.53 µg·L-1 on day 20, followed by a fluctuating decline, ending at 71.79 µg·L-1 on day 60. In contrast, HD achieved the highest IAA values much later at 50 days with a value of 103.64 µg·L-1. FA showed a significant peak IAA value of 99.11 µg·L-1 on day 30, followed by a rapid decline to 73.27 µg·L-1 by day 60. FD showed a gradual increase in the IAA content from a relatively low initial value of 58.92 µg·L-1 to 83.78 µg·L-1 after 60 days, slightly lower than the peak value of 84.96 µg·L-1 observed at 40 days. JX and HG showed relatively stable IAA levels throughout the cutting period, with JX ranging from 65.85 to 83.28 µg·L-1. However, HG recorded a significant increase in IAA at 30 and 60 days, with a peak value of 94.65 µg·L-1 on day 30.
(2) CTK: We also analyzed the dynamic changes in the CTK content during the cutting process of the six tea varieties (Fig. 4B). HD, which had the lowest root formation rate, initially exhibited a relatively high CTK level of 59.11 µg·L-1, remaining consistently elevated throughout the rooting process and ending at 53.88 µg·L-1 on day 60. In contrast, HG started with a similarly high initial CTK value of 58.19 µg·L-1 but showed a gradual decrease, reaching its lowest level of 35.78 µg·L-1 at 50 days before recovering slightly by day 60. JX, which demonstrated the highest rooting rate, showed a continuous decrease in its CTK value from the time of cutting, starting at 41.31 µg·L-1 and reaching a nadir of 30.79 µg·L-1 at 50 days before increasing slightly to 43.61 µg·L-1 by day 60. YD, which also exhibited strong rooting ability, had an initial CTK value of 52.58 µg·L-1, which dropped significantly to 32.78 µg·L-1 at 50 days before recovering. The CTK levels in FA and FD peaked at 20 days and then decreased before increasing slightly after 60 days.
(3) ABA: In this study, we also observed changes in the ABA content during the cutting process of the six tea varieties (Fig. 4C). YD had the highest initial ABA content at 425.93 µg·L-1, subsequently showing an overall declining trend, reaching the lowest level of 277.72 µg·L-1 at 30 days. In contrast, HD and JX showed significant fluctuations in the ABA content. Notably, JX reached a peak of 448.97 µg·L-1 after ten days and then declined rapidly. FA exhibited a significant decrease in the ABA content after 20 days, followed by a notable increase to 431.15 µg·L-1 after 60 days. FD and HG maintained relatively stable ABA levels throughout the cutting process.
(4) Changes in endogenous hormone ratios: According to Fig. 4D, the IAA/CTK ratio for HD increased from an initial value of 1.28 to a peak of 1.94 after 50 days before declining slightly. YD showed a continuous increase in the IAA/CTK ratio, reaching a peak of 2.44 on day 50. FA had an IAA/CTK ratio that peaked mid-cycle and then slightly declined. FD peaked at 2.13 after 30 days, followed by a gradual decline. JX maintained a relatively high IAA/CTK ratio throughout the cutting cycle. HG exhibited a high IAA/CTK ratio in the early and middle stages of the cutting process. According to Fig. 4E, HD and YD showed relatively small fluctuations in the IAA/ABA ratio during the cutting process. FA reached a peak in the IAA/ABA ratio after 20 days. FD, JX, and HG showed higher IAA/ABA ratios during key rooting periods. According to Fig. 4F, HD and YD showed relatively small changes in the CTK/ABA ratio throughout the entire cutting cycle. FA and FD exhibited increases in the CTK/ABA ratio at specific time points. JX and HG showed higher CTK/ABA ratios during critical rooting periods.
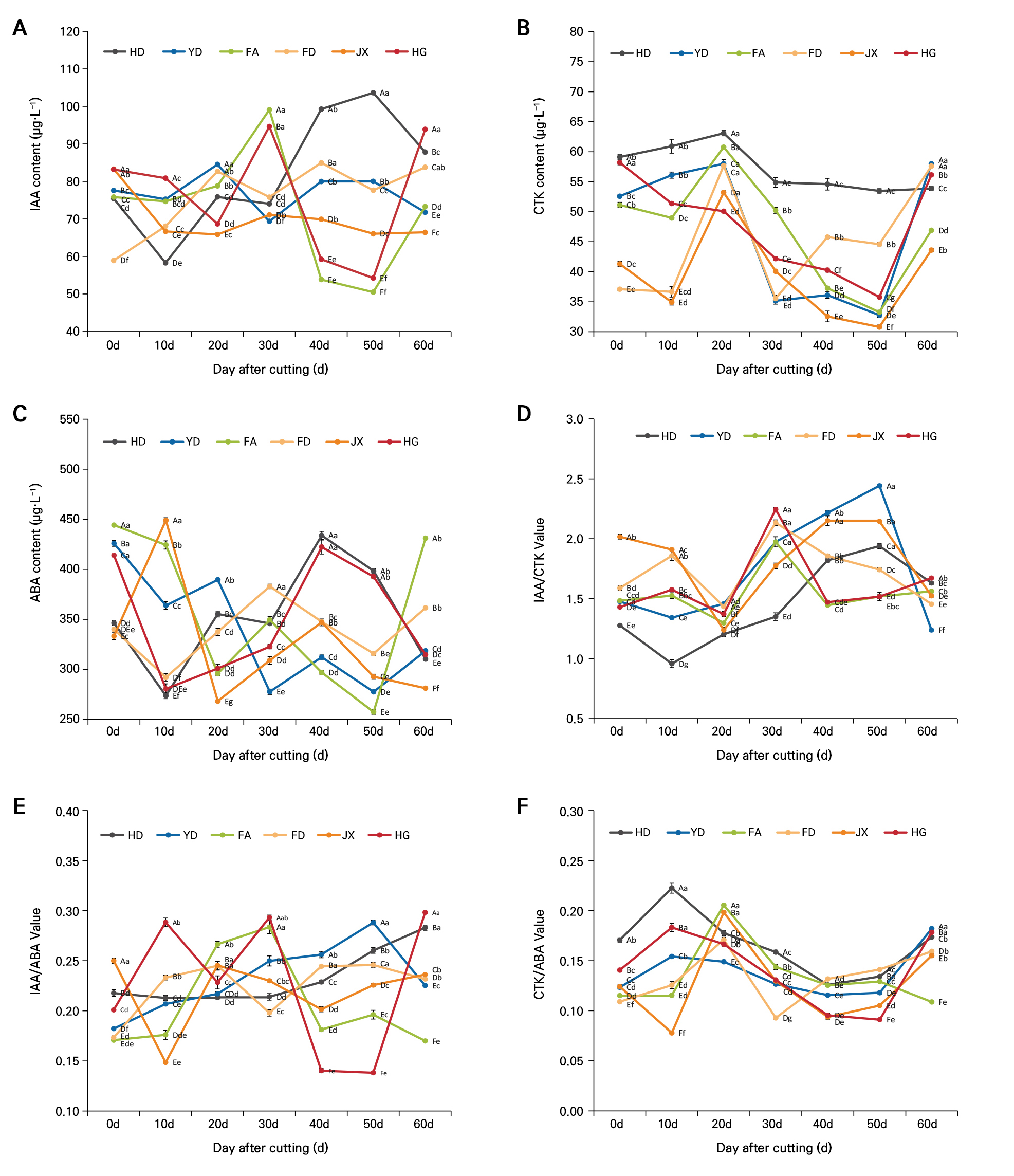
Fig. 4.
Changes in the endogenous hormone contents and ratios during the rooting process of different tea plant varieties: IAA content (A), CTK content (B), ABA content (C), IAA/CTK (D), IAA/ABA (E) and CTK/ABA (F). Different capital letters indicate significant differences among different varieties at the same period (p < 0.05), while different lowercase letters indicate significant differences within the same variety over different periods (p < 0.05).
Discussion
Effects of physiological and biochemical indicator changes on the root formation of different tea varieties
Changes in the soluble sugar and protein contents are fundamental to supporting the root formation process (Tombesi et al. 2015). The pattern observed during the cutting process showed that both the soluble sugar and protein contents decreased initially and then increased. In the early stages of cutting, the cuttings experienced mechanical damage, consuming large amounts of sugars and proteins for wound healing, cell division, and root primordium formation. In the later stages, as new roots formed and grew, photosynthesis and metabolic activities resumed, leading to increased synthesis of sugars and proteins, as reflected in their rising levels. This conclusion aligns with the findings from the rooting and biochemical property analyses of other plants, such as Callicarpa nudiflora (Li et al. 2016) and Eucommia ulmoides (Lv et al. 2021). Additionally, this study found that high rooting rate varieties, such as FA, exhibited a higher soluble sugar content in the early rooting stage, providing the necessary energy for new root formation. Conversely, varieties with weaker rooting abilities, such as HD and HG, did not show significant increases in the soluble sugar content during rooting, which may affect their rooting efficiency. The study also observed that the callus formation period for HD was longer and that the callus volume was larger during the rooting process compared to other varieties. Determining whether extensive callus formation competes with AR formation for nutrients, resulting in lower rooting rates of cuttings with excessive callus growth, requires further investigation. Previous research by Liang et al. (1985) on the rooting of tea cuttings in ‘Zhenong 12’ also found that callus formation and AR differentiation were almost simultaneous but independent processes. Furthermore, the lignification of the callus after its formation can inhibit AR production (Chang et al. 2023). Research has suggested that removing the basal callus can promote rooting in cuttings (Li et al. 2014).
PPO, POD, and IAAO are closely related to rooting. Studies have shown that increased PPO activity may contribute to callus formation, while enhanced POD activity may protect against oxidative stress (Lv et al. 2022). High PPO activity can catalyze the formation of “IAA-phenolic acid compounds,” thus promoting the formation of ARs. The changes in PPO activity are closely linked to the rooting process of cuttings. Research on Eucommia ulmoides (Lv et al. 2022) and Lycium barbarum (Su et al. 2023) has also shown that PPO and POD activities initially rise and then fall after cuttings, outcomes that are closely related to the rooting process. In this study, early increases in PPO and POD activities were particularly evident in the high-rooting rate varieties. YD exhibited high initial PPO activity, after which its PPO activity gradually decreased during the rooting process. JX showed the highest PPO activity among the six varieties on day 20, with the peak occurring during the critical period of AR development. This indicates that these varieties completed most of the callus and root primordium formation steps earlier than in the other varieties, reducing the demand for PPO in the later stages. In contrast, HD had high initial PPO activity but continued to fluctuate until peaking on day 60, indicating slower callus formation and a slower rooting process, consistent with its slow rooting progression. Most studies have shown that lower IAAO activity levels during the induction and development stages of ARs favor the accumulation of IAA in the cuttings, promoting the formation and elongation of ARs (Zhang et al. 2017). In this study, all six varieties showed a decrease in IAAO during the critical stages of AR induction and development. JX maintained a consistently lower IAAO level compared to the other varieties during the late rooting stages, promoting AR formation, which corresponded with its high rooting rate. HD, FA, and HG maintained high IAAO activity levels during the mid to late rooting stages, resulting in relatively low rooting rates, similar to previous research findings (Porfirio et al. 2016; Denaxa et al. 2019). While increased PPO and POD activities and decreased IAAO activity levels generally favor rooting, the final rooting efficiency is influenced by multiple physiological and molecular factors, including the dynamic balance of other hormones, the activities of various oxidases, and environmental conditions. For a more comprehensive understanding and optimization of the root formation process, further research into the molecular basis of these complex interactions is required.
Previous studies have shown that the formation of ARs centers on auxin, with various endogenous hormones regulating this process through a dynamic hormonal balance (Pacurar et al. 2014; Mello et al. 2016). IAA is a thoroughly investigated endogenous auxin, with numerous studies confirming its crucial role in AR formation (Jing and Strader 2019). The present study demonstrates that changes in endogenous hormones can affect the rooting rate and rooting index in different tea varieties significantly. Dynamic changes in hormones such as IAA, CTK, and ABA are closely related to rooting outcomes. During the AR initiation period, both YD and HD exhibited high levels of IAA and CTK. However, YD had lower ABA levels than HD, resulting in a higher rooting rate for the former. This suggests that moderately increasing IAA and CTK levels while controlling ABA levels during critical periods can improve rooting rates (Da-Costa et al. 2013). This finding aligns with research on Camellia sinensis (Fan et al. 2021), Paeonia (Wen et al. 2023), and Camellia oleifera (Liu 2022), where higher IAA/CTK and IAA/ABA ratios are typically associated with higher rooting rates and rooting indices. The dynamic balance between hormone levels and their ratios plays a key role in the cutting-rooting process of tea plants. Successful rooting depends not only on the level of a single hormone but also on the combined regulation of oxidase activities and environmental factors.
Commonalities and differences in the root formation process of medium-leaf and large-leaf tea varieties
In this study, the classification of medium-leaf and large-leaf varieties was based on the definitions in an existing catalog of approved tea varieties and on the findings of previous studies (Yang et al. 2003; Wang et al. 2020), with large-leaf and medium-leaf varieties mainly differing according to the leaf area, a widely accepted criterion in tea plant breeding and research. In terms of common characteristics, both medium-leaf and large-leaf varieties demonstrated that IAAO activity plays a crucial role in the rooting process of different varieties, particularly during the callus induction period and the AR expression and elongation stages. Additionally, high levels of soluble sugars and proteins positively impacted the rooting process. Increased levels of IAA and CTK during critical growth stages, such as the AR initiation period, and a subsequent decrease in CTK levels during the later stages of AR development, help improve rooting rates, while excessively high levels of ABA inhibit rooting. Moderately controlling ABA levels enhances rooting efficiency.
Regarding differences, this study found that most medium-leaf varieties had higher rooting rates and rooting indices compared to large-leaf varieties. Genetic variation plays an important role in determining how different varieties respond to rooting hormones, nutrients and environmental factors (Mello et al. 2016). Leaf area has been identified as one of the key factors affecting rooting success in cuttings (Atangana et al. 2006). Previous studies have shown that medium-leaf varieties typically have higher rooting rates than large-leaf varieties (Dong 1984; Wu et al. 2014). Variations in the cutting leaf area can indirectly affect physiological properties by affecting photosynthesis, nutrient accumulation and transpiration. For example, the assimilation products produced by the leaves through photosynthesis provide important nutrients for the root development of cuttings (Zhang et al. 2023). However, during the early rooting phase, the nutrients in the cuttings come primarily from their own stored reserves and not from assimilates (Tsafouros et al. 2019). In addition, leaf transpiration results in excessive water loss, which causes cuttings to wilt and negatively affects root formation, especially on cuttings with short stems (Tchoundjeu and Leakey 1996). Therefore, the balance between assimilate accumulation and transpiration could be one of the key factors explaining why most medium-leaf varieties have higher root formation rates than large-leaf varieties. In addition, studies of the evolution and phylogenetic relationships of tea germplasm have shown that during domestication, small- and medium-leaf populations are subjected to greater selection pressure for disease resistance and taste than large-leaf populations. This indicates that already bred small and medium-leaved varieties generally have better disease resistance, making their cuttings less susceptible to root rot and allowing them to achieve higher survival rates during propagation (Wang et al. 2020).
Although leaf size can indirectly affect the rooting process, it is not the determining factor. Significant differences in root formation ability were observed between two large-leaf cultivars, HD and YD. Endogenous hormone regulation plays a key role in the rooting mechanism, with medium-leaved varieties demonstrating more favorable dynamic changes in IAA and CTK levels for rooting. In contrast, the relatively low root formation rate observed in HD was associated with consistently high CTK levels, which negatively impacted the root formation efficiency.
Conclusions
This study systematically analyzed the dynamic changes occurring in physiological and biochemical indicators during the rooting process of cuttings from six tea varieties with different rooting abilities. The rooting times varied significantly among the varieties, with HD having the longest rooting time (69.90 days) and JX the shortest (38.40 days), indicating notable differences in rooting dynamics and genetic backgrounds among the varieties. YD, FD, and JX exhibited higher rooting rates and rooting indices, demonstrating strong rooting abilities, whereas HD and HG showed weaker rooting abilities. During critical growth periods, varieties with strong rooting abilities, such as JX and YD, exhibited higher soluble sugar and protein contents, while HD demonstrated relatively weaker physiological regulation and adaptability. Changes in PPO and POD activities were closely related to the rooting process, with high-rooting varieties showing elevated early activities that later declined. The decrease in IAAO activity contributed to the accumulation of IAA, promoting the formation of ARs. The dynamic changes in endogenous hormones, including IAA, CTK, and ABA, significantly affected rooting rates and rooting indices. The ratios of IAA/CTK and IAA/ABA were positively correlated with the rooting rates. YD and JX exhibited higher IAA/CTK ratios, facilitating their rapid and efficient rooting. In summary, the differences among tea varieties significantly impacted the efficiency of cutting propagation. High-rooting varieties demonstrated strong rooting abilities through higher nutrient contents, appropriate oxidase activities, and effective endogenous hormone regulation. These findings provide a solid foundation for improving propagation techniques and understanding the physiological mechanisms of root formation. Future research could focus on identifying the key genes and molecular pathways involved in hormone metabolism and oxidase activity during the root formation process. Furthermore, studying the interactions between certain environmental factors (e.g., temperature, humidity, light) and physiological traits could help optimize cutting propagation practices for different tea varieties. Such studies could pave the way for more efficient propagation strategies, especially for difficult-to-root varieties, and ultimately contribute to improved tea production and resource utilization.
