

서 언
Berchemia속 식물은 낙엽활엽교목인 망개나무(Berchemia berchemiaefolia(Makino) Koidz.)를 포함하여 덩굴성 식물인 먹넌출(B. racemosa var. magna Makino)과 청사조(B. racemosa Siebold & Zucc. var. racemosa) 등 3 수종이 자생하고 있으며 모두 희귀식물로 지정되어 있다(KNA, 2008; Lee, 2003). 그 중 먹넌출은 낙엽활엽의 덩굴성 식물로서 한국과 일본에 분포하며, 국내에서는 안면도에만 유일하게 분포하는 수종이다(KNA, 2008; Lee, 2003). 먹넌출의 기본종인 청사조는 전통적으로 약용으로 활용되었는데 최근 추출물이 백혈병 세포 억제에 효과적인 것으로 나타났으며, 먹넌출 또한 추출물을 이용한 항산화 또는 미백 기능이 입증되어 유용한 식물자원으로서의 가치를 인정받고 있다(Chen and Carsten, 2007; Ji et al., 2012; Wang et al., 2006).
먹넌출은 국가단위에서 취약종(VU/Vulnerable)으로 평가되어 있으며, 최근 기후변화에 따른 서식지 환경 악화로 인해 소멸 위험성이 매우 높아 자생지 확인 및 생명자원 보존을 위한 현지내・외 보전대책이 요구되고 있다(KNA, 2008). 덩굴성 식물로서 나무를 지지하고 올라가며 종자에 의한 유성번식과 줄기를 지표면에 내려 개체를 형성하는 무성번식을 하는 수종이다. 집단 내 개체수가 적고 보존 가치가 높은 먹넌출의 효율적인 보존을 위해서는 유전다양성뿐만 아니라 집단 내 형성되어 있는 공간적 유전구조를 파악하는 것이 매우 중요하다(Escudero et al., 2003). 또한 먹넌출과 같이 유・무성번식을 겸하는 식물은 특히 무성번식의 강도가 집단내 유전적 공간구조에 큰 영향을 주기 때문에 집단의 유전적 연구와 병행하여 클론 형성능(clonality)을 파악해야 한다(Chung and Epperson, 2000).
지금까지 국내에서 Berchemia속에 대한 연구를 살펴보면, 망개나무에서만 자생집단의 식생군집 및 수목활력도 분석(Park and Cho, 2011), 잎의 형태적 변이(Song et al., 2008) 및 유전적 특성(Lee et al., 2003)에 대한 연구가 수행되었다. 중국과 일본에서는 청사조 추출물에 대한 약용적 효과가 보고되고 있으나(Sato et al., 1995; Wang et al., 2006) 생태적 특성과 유전적 특성에 대한 연구는 미흡하다. 국내에서는 청사조에 대한 식생구조 및 개체군 동태에 대한 연구와 먹넌출에 대한 삽목증식법(Song et al., 2011), 추출물에 대한 기능성 연구가 수행된 바 있으나(Beon and Kim, 2008) 소멸위험성을 고려한 현지외 보존을 위한 표본추출 전략 개발이 요구되고 있다. 특히, 국제적으로는 생물다양성협약(CBD)의 지구식물보전전략(GSPC)에서 알려진 위협종의 최소 75%를 현지외 보전토록 요구하고 있어 보존 기술 개발이 시급한 실정이다(UNEP, 2010).
일반적으로 국소환경 조건에 민감한 고산 수종을 포함한 희귀수종은 집단크기(개체수) 감소나 환경 급변에 적응력이 떨어지기 때문에 기후변화로 인한 지구온난화에 더욱 심각한 영향을 받을 수 있다(Pearman et al., 2007). 유전다양성의 유지는 대상 종의 생존과 보존을 위해서 매우 중요하며, 특히 임목의 경우 한 장소에서 오랫동안 생존하며 다양한 환경변화에 대하여 적응해야 되기 때문에 유전다양성의 확보는 생존과 진화를 위해서 필수적이다.
따라서 본 연구는 I-SSR 표지자 분석을 이용 소멸위기 수종인 먹넌출의 유전다양성과 공간분포에 따른 유전적 구조를 분석하고 현지외 생명자원 보존을 위한 보존표본 추출 전략을 제시하고자 수행되었다.
재료 및 방법
조사지역 개황 및 시료채취
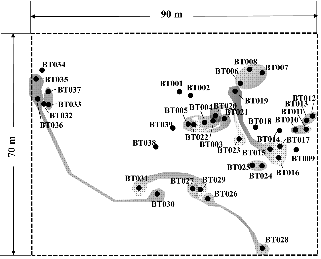
본 연구의 조사지는 국내에서 유일한 안면도 먹넌출 자생집단(해발고 약 53m, 북서사면)을 대상으로 하였다. 조사지 주변에는 소나무(Pinus densiflora Siebold & Zucc.), 졸참나무(Quercus serrata Thunb.), 때죽나무(Styrax japonicus Siebold & Zucc.), 산뽕나무(Morus bombycis Koidz.), 말오줌때(Euscaphis japonica (Thunb.) Kanitz), 다래(Actinidia arguta (Siebold & Zucc.) Planch. ex Miq.) 등이 서식하고 있다. 가로 90m(남동방향)와 세로 70m(북서방향)의 조사구를 설치하고, 조사구에 분포하는 총 39개의 먹넌출 개체의 크기와 위치를 기록하였다. 줄기가 서로 분리되어 있으며 독립된 줄기를 가지는 특징을 근간으로 개체를 구분하였으며 각 개체의 유전적 동질성을 파악하기 위하여 모든 개체에서 유엽을 채취하였다.
DNA 분리 및 I-SSR PCR
채취된 먹넌출의 유엽에서 DNeasy Plant Mini Kit(Qiagen, Germany)를 이용하여 DNA를 분리하였다. DNA는 분광광도계(ND-1000, Nano-Drop Technologies, USA)를 이용하여 정량하였다. 중합효소연쇄반응(Polymerase chain reaction, PCR)은 유전자증폭기(Gene Amp PCR System 9700, Applied Biosystems, USA)를 사용하여 주형 DNA 6ng, 10X buffer, 1mM dNTP, 0.025% BSA, 2mM MgCl2, 1.5μM I-SSR primer 3.75μL, Promega Flexi GoTaq(Promega Corp., Madison, WI) 0.6U 및 증류수가 포함된 총 15μL의 반응액을 95°C에서 5분 동안 1차 해리한 후, 해리 95°C 1분, annealing 50-54°C 1분, extension 72°C 2분으로 구성된 반응을 30회 반복한 후 최종적으로 72°C에서 10분간 extension하여 수행하였다. I-SSR primer는 UBC primer Set #9(University of British Columbia, Canada)중 46개를 사용하였으며, 이 가운데 재현성이 우수하고 다형성을 보이는 8개의 primer를 선정하였다(Table 1). PCR 증폭산물은 1×TBE buffer를 사용하여 2% agarose gel에서 전기영동하고, ethidium bromide로 정색하여 UV trans- illuminator에서 증폭산물을 확인하였다. 증폭산물의 유무에 따라 ‘1’과 ‘0’으로 통계분석 자료를 입력하였다.
자료분석
Clark and Evans(1954)의 군집지수(aggregation index, R)를 사용하여 조사지역 내 먹넌출의 분포형태를 파악하고, 밀도, 피도, clump간 평균거리, clump의 평균면적 등을 구하여 집단의 개황을 분석하였다. Clark and Evans(1954)의 군집지수(R)는 개체들의 공간적 분포가 임의 분포일 경우는 1이고, 1보다 작은 경우에는 집중분포를 나타내며, 1보다 클 경우는 균일한 분포를 의미한다(Krebs, 1999).
I-SSR PCR에 의해 생성된 다형성 증폭산물을 바탕으로 POPGENE 1.32 프로그램(Yeh et al., 1997)을 이용하여 Shannon의 다양성지수(diversity index, I; Lewontin, 1972)를 구하여 먹넌출 집단의 유전다양성을 추정하였다. 모든 개체의 유전자형을 비교하여 개체별 유성생식체(genet)의 구성 여부를 판단하였고 조사된 모든 위치에서의 증폭산물 표현형이 일치하는 개체들은 해당 유전자에서의 유전자형이 동일한 것으로 간주하여 무성생식체(ramet)로 처리하였다.
클론형성능 분석을 위해 유전자형 비율(G/N), 유전자형 다양성(D)과 유전자형 균등도(E) 등 세가지 통계량을 계산하였다. 유전자형 비율은 관찰된 유성생식체의 수(G)를 총 개체수(N)로 나눈 비율로 계산하였다(Ellstrand and Roose, 1987). 유전자형 다양성(genotype diversity)은 제한된 표본크기를 고려하여 변형된 Simpson의 다양성지수(Pielou, 1969)를 이용하였고 유전자형 균등도(genotype evenness)는 Fager (1972)의 방법을 이용하였다.
유전적 공간구조를 구명하기 위해 Tanimoto distance를 이용하여 먹넌출 집단의 공간적 자기상관 분석을 실시하였으며, 자료분석은 SGS v.1.0d 프로그램(Degen et al., 2001)을 사용하였다. Distogram에서의 거리등급(distance class)은 6m 간격으로 10개 거리등급으로 나눴고, 각 거리등급에서 3,000회의 permutation분석에 의해 95% 신뢰구간을 산출하여 자기상관성의 유의성 여부를 검정하였다. 또한 무성번식이 유전적 공간구조에 미치는 영향을 알아보기 위해 유성생식체 수준에서의 공간구조도 동일한 방법으로 조사하였다. 2개 이상의 개체에서 동일 유전자형이 나타나는 경우 근원경이 큰 개체를 모수로 가정하였다. 유전적 거리와 지리적 거리의 상관관계를 알아보기 위해 유성생식체 수준에서 Mantel 검정을 실시하였으며, 자료분석은 Isolation by Distance Web Service(IBDWS) v.3.21 프로그램(Jensen et al., 2005)을 사용하였고 10,000회의 permutation 분석에 의해 유의성 여부를 검정하였다. 유전적거리는 Nei(1978)의 방법을 이용하였으며, Phylip v.3.69 프로그램(Felsenstein, 2009)을 사용하여 계산하였다.
결 과
먹넌출 집단의 현황 및 공간분포
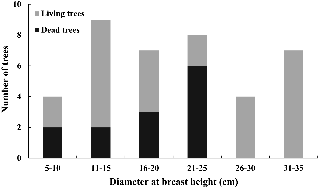
조사구 6,300m2(70m × 90m) 내에 총 39개체가 분포하였으며, 밀도는 0.006개/m2였다(Fig. 1). 먹넌출의 평균 근원경은 3.7cm였으며, 1.5cm의 가장 작은 개체부터 6.5cm의 가장 큰 개체까지 다양한 크기로 분포하였다. 개체의 중심간 평균 거리는 5.96m였다. 먹넌출의 지지식물은 소나무, 산뽕나무, 나도밤나무, 때죽나무, 졸참나무로 전체 5수종으로 나타났으며, 평균 흉고직경(diameter at breast height, DBH)은 19.5cm였다. 동일 수목에 먹넌출이 2개체에서 많게는 5개체까지 올라가는 경우도 있었다. 전체 지지식물의 33%는 고사목이였으며 이러한 현상은 25cm 이하의 직경이 작은 개체에서만 나타났다(Fig. 2). 채집한 먹넌출의 위치자료를 바탕으로 군집지수(R)를 계산한 결과 군집지수는 0.706(P < 0.001)으로 먹넌출 개체들은 약한 집중분포 형태를 나타내고 있었다.
먹넌출 집단의 유전구조
8개 I-SSR primer에서 생산된 증폭산물 중에서 식별이 뚜렷한 총 50개를 분석 대상으로 하였다. 이 중 다형성 증폭산물은 13개, 단형성 증폭산물은 37개로서 다형성 증폭산물의 비율은 26.0%였다(Table 1).

13개의 다형성 증폭산물을 이용하여 모든 개체의 유전자형(multilocus genotype)을 비교한 결과 총 39개체 중 유전자형이 서로 다른 21개의 유성생식체가 식별되어 유전자형 비율(G/N)이 53.8%로 나타났으며, Shannon의 다양성지수는 0.598로 나타났다(Table 2). 동일한 유전자형을 가진 개체들은 대부분 지리적으로 근접해 있었고, 특별한 물리적 장애물이 존재하지 않았으므로 무성생식체로 판단할 수 있었다. 일부 개체들(BT022, BT023, BT028, BT030, BT031)은 지리적으로 최대 47m까지 떨어져 분포하고 있었다. 21개 유성생식체의 분포양상을 살펴보면 단일유전자형을 지닌 유성생식체가 9개(43%)로 가장 높게 나타났으며, 나머지 12개 유성생식체는 2-4개의 무성생식체로 구성되었다(Fig. 3). 조사구 내의 먹넌출 집단의 유전자형 다양성은 0.966, 유전자형 균등도는 0.946로 나타났다. 유전자형 다양성(D)은 집단내에서 두 개체를 추출할 때 두 개체가 서로 다른 유전자형을 가지는 확률로서 모든 개체가 동일한 유전자형을 가질 경우 0의 값을, 모든 개체가 서로 다른 유전자형을 가질 경우 1의 값을 가진다. 또한 유전자형 균등도(E)는 유전자형의 균일한 정도를 나타내는 지수로서 모든 개체가 동일한 유전자형을 가질 경우 0의 값을, 모든 개체가 서로 다른 유전자형을 가질 경우 1의 값을 가진다.
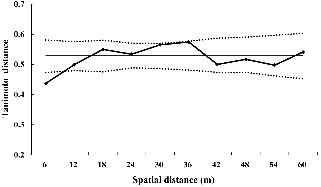
Tanimoto distance를 이용한 공간적 자기상관성 분석을 실시한 결과 조사구 내의 먹넌출은 약 6m 이내에서 분포하는 개체들 간에 유전적 유사성이 높아 자기상관성이 인정되었고, 6m 이상에서 임의분포를 나타냈다(Fig. 4). 공간적 유전구조는 클론 형성의 정도에 따라 영향을 받으므로(Reusch et al., 1999) 유성생식체 수준에서의 공간구조를 파악한 결과 6m 내에 존재하는 개체 간 유전적 거리가 0.44에서 0.62로 높아지면서 유전적 동질성이 완화되었으며, 모든 구간에서 임의분포를 보여 무성생식체 수준의 공간적 유전구조와 차이를 나타냈다(data not shown). 유성생식체 수준에서 Mantel 검정을 실시한 결과 먹넌출은 유전적 거리와 지리적 거리간의 상관관계가 나타나지 않았다(r = 0.06, P = 0.31).
고 찰
I-SSR 표지자로 분석한 안면도 먹넌출 집단의 Shannon의 다양성지수(I)는 0.598로 소나무 0.450(Hong et al., 2004), 눈향나무 0.463(Choi et al., 2004b), 눈잣나무 0.567(Song et al., 2012), 복분자딸기 0.576(Kang et al., 2003) 수종들에 비해 높은 값을 나타냈다. 기존의 I-SSR 표지자를 이용한 연구는 4-9개 I-SSR primer를 사용하여 21-48개의 다형성밴드가 나타났는데(Choi et al., 2004a, 2004b; Kang et al., 2003; Song et al., 2012), 먹넌출은 8개의 I-SSR primer에서 13개의 다형성밴드를 보여 다른 수종들에 비해 적은 특징을 보였다. 일반적으로 식물종이 가지는 유전다양성은 지리적 분포특성과 교배양식 및 번식방법의 영향을 크게 받는다(Hamrick et al., 1992). 현재의 동북아시아의 먹넌출과 근연종인 청사조의 분포를 살펴보면 중국에서는 해발 2,600m 이하의 중부 이남에 넓게 분포하며 일본에서도 북해도를 비롯하여 혼슈 지역에 넓게 분포하고 있으나 한반도에서는 안면도와 군산 지역에서만 제한적으로 분포하고 있다(Chen and Carsten, 2007; Kawamoto, 1943; KNA, 2008; Lee, 2003). 이러한 분포는 Kong(2002)이 언급한 한반도의 일부 고산식물의 분포와 일치하며, 기존 한반도의 격리・잔존 식물종에 대한 연구결과에서 추정된 바와 같이 과거 빙하기 때 먹넌출이 현재보다 넓은 지역에 걸쳐 연속적으로 분포하였으며, 이때 안면도 집단이 분포 중심지에 가깝게 존재했을 것으로 추정된다(Kwon and Kim, 2002; Lee et al., 1997). 먹넌출은 무성번식과 유성번식을 겸하는 번식방법을 가지고 있고 집단 내 대부분의 개체들이 각기 다른 유성생식체로 이루어진 것에서 알 수 있듯이(Fig. 3) 무성번식보다는 종자에 의한 유성생식에 의해 개체가 전파되고 집단이 형성되는 비율이 크기 때문에 비교적 높은 유전다양성을 유지할 수 있었으며, 이러한 결과는 제한적으로 분포하는 눈향나무(Choi et al., 2004b), 눈잣나무(Song et al., 2012), 시로미(Choi et al., 2004a)의 경우에서도 보고된 바 있다. 먹넌출의 근연종인 망개나무의 경우 먹넌출과 마찬가지로 제한적인 분포를 보이는데(KNA, 2008), 동위효소를 이용한 유전다양성 분석에서 평균이형접합도 기대치(He)가 0.019로서 희귀식물인 미선나무 0.125에 비해 낮게 나타났으며(Chang et al., 2003), Lee et al.(2003)이 I-SSR 표지자와 동위효소를 이용한 분석에서도 유전적 변이가 매우 낮게 나타났다. 망개나무는 인간의 무분별한 벌채로 인해 교란을 받았으며, 교목성 수종으로 종자에 의한 유성생식을 통해 개체가 형성되는데, 열매가 성숙하기 전 7-8월에 집중호우로 인한 낙과(Chang et al., 2003)로 인해 차대 형성이 어렵기 때문에 유전다양성이 낮게 나타나는 것으로 추정된다.
종 수준에서 유전자형 비율(G/N)은 53.8%로 종자 및 무성번식을 겸하는 눈잣나무와 우묵사스레피에서 보고된 유전자형 비율(61.3-61.5%)에 비해 낮게 나타났는데(Chung and Epperson, 2000; Song et al., 2012), 이러한 원인은 Beon and Kim(2008)이 근연종인 청사조에서 보고된 바와 같이 덩굴성인 먹넌출이 줄기에 의한 무성번식 비율이 관목성인 타 수종에 비해 높기 때문으로 추정된다. 무성번식의 정도가 타수종에 비해 높게 나타났으나 유성생식의 비율이 무성번식에 비해 높고 유전자형다양성과 유전자형균등도가 각각 0.966, 0.946으로 높게 나타나서 전체적인 집단의 유전다양성이 높게 유지되는 것으로 판단된다. 무성번식의 공간적 자기상관성 분석 결과 먹넌출은 약 6m 내에 분포하는 개체들 간에 자기상관성을 보였으며(Fig. 4), 무성번식과 종자번식을 겸하는 식물 중 수고가 낮은 시로미(Choi et al., 2004a)와 눈잣나무(Song et al., 2012)와 비슷한 수준(8-12m)으로 나타났다.
안면도 먹넌출 집단은 유전다양성이 비교적 높은 수종으로 분석되었으나 분포면적이 매우 제한적이고 개체수가 많지 않기 때문에 앞으로도 유전적 부동의 영향을 받을 가능성이 높다. 현지외 유전자 보존을 위한 개체 선발시 최소 6m 이상의 거리를 두면 이질적인 개체를 선발할 가능성이 매우 높으나, 개체수가 많지 않고 분포면적이 좁은 것을 고려할 때 전체 개체에서 종자를 채취하여 모든 유전자형을 확보하는 것이 타당할 것이다. 아울러 먹넌출 집단의 적극적인 보존을 위해서 현 자생지의 지속적인 모니터링과 종자 및 삽목을 통한 번식방법에 대한 연구를 통해 현지외 보존에 대한 접근도 함께 이루어져야 할 것이다.
