

Introduction
Materials and Methods
Fruit samples, treatments, and storage conditions
Assessments of physiological characteristics
RNA extraction and expression of ethylene-related genes
Statistical analysis
Results
Ethylene production and respiration rate
Firmness, titratable acidity (TA), and soluble solids content (SSC)
Weight loss and starch pattern index (SPI)
Fruit peel color and fruit appearance
Expression of ethylene biosynthesis and receptor genes
Correlation coefficient matrix
Discussion
Introduction
Ethylene is a natural plant hormone that regulates ripening in several climacteric fruits, including apples (Seymour et al. 2012). The biological ripening process occurs after ethylene binds to receptor complexes, after which the fruit begins to soften and turns yellow (Barry and Giovannoni 2007; Seymour et al. 2012). However, excessive ethylene production during ripening adversely affects fruit texture, flavor, and appearance, ultimately shortening the shelf life of the fruit (Barry and Giovannoni 2007).
In general, ethylene regulates the ripening process by regulating the gene expression of key enzymes involved in ethylene biosynthesis and signaling pathways (Liu et al. 2015b; Latt et al. 2025). For example, the ethylene biosynthesis pathway initiates with methionine as the precursor and finally yields ethylene via the action of S-adenosyl methionine, 1-aminocyclopropane-1-carboxylate (ACC) synthase (ACS), and ACC oxidase (ACO) (Yang and Hoffman 1984; Bakshi et al. 2015). The critical roles of ACS and ACO in fruit ripening have been reported for several apple cultivars (Wiersma et al. 2007; Win et al. 2025). After synthesis, ethylene is recognized by ethylene receptors (ETRs, ERSs, and EINs), which interact with downstream constitutive triple response 1 (CTR1). Subsequently, CTR1 transmits the signal to ethylene insensitive 2 (EIN2) (Wang et al. 2002; Wang et al. 2007; Park et al. 2023). EIN2 represses the ethylene signal to ethylene insensitive 3 (EIN3) and ethylene insensitive 3-like (EILs) and then activates the ethylene signal to ethylene response factor 1 (ERF1) (Solano et al. 1998). Thus, ethylene combines with its receptors, resulting in signal transduction to initiate fruit senescence (Mata et al. 2021). The shelf life of climacteric fruits can be extended by inhibiting ethylene production and its effects (Giovannoni 2007; Martinez-Romero et al. 2007; Akcay et al. 2025).
1-Methylcyclopropene (1-MCP) is extensively used to inhibit ethylene production and its effect on apple ripening (Zhang et al. 2020; Sidhu et al. 2024). 1-MCP is an ethylene perception inhibitor that blocks ethylene at the receptor sites (Sisler and Serek 1997). Ethylene biosynthesis and perception genes in apples can be downregulated via 1-MCP-mediated inhibition of ethylene production (Tatsuki and Endo 2006; Tatsuki et al. 2009; Latt et al. 2025). However, the effects of 1-MCP vary depending on the cultivar (Watkins 2006; Watkins 2008; Sidhu et al. 2024). Salicylic acid (SA), a naturally occurring phenolic compound, is used to enhance resistance to pathogens and control postharvest decay in horticultural crops (Xu et al. 2018; Zhao et al. 2021). SA can reduce ethylene production along with ACS and ACO activities during ripening (Zhang et al. 2021; Li et al. 2022). Xu et al. (2018) found that the combination of SA+1-MCP maintains an optimal quality of bananas. Recently, the combination treatment of SA+1-MCP has been shown to induce disease resistance, activate the mechanism of disease resistance-related enzymes, and maintain quality of postharvest fruits (Meng et al. 2023). However, the effect of SA, 1-MCP, and their combination treatment on the ripening and ethylene perception mechanisms of fruits, particularly apples, is not fully understood.
‘Picnic’ is a locally developed new Korean apple cultivar that has recently gained popularity owning to its superior quality attributes (Ban et al. 2014; Yoo et al. 2016). A recent study reported that ‘Picnic’ is prone to wilting during postharvest storage which could be related with the climacteric ripening or fruit maturity (Kwon et al. 2025). ‘Picnic’ apples have a low internal ethylene concentration (IEC) among other new Korean apple cultivars at harvest and during storage (Yoo et al. 2016). Hence, we investigated the changes in the physiological characteristics of this fruit associated with ethylene production and perception during ripening. We also aimed to identify the optimal process to extend the shelf life of the fruit. Therefore, we evaluated the effects of SA, 1-MCP, and their combination treatment on the ripening mechanism of ‘Picnic’ apples during storage. We also determined the responses of ethylene-related candidate genes (MdACS1, MdACO1, MdETR1, MdERS2, MdCTR1, MdEIN2, MdEIL1, and MdERF1), which are mainly involved in ethylene production and perception mechanism.
Materials and Methods
Fruit samples, treatments, and storage conditions
‘Picnic’ apples were harvested from a commercial orchard in Gunwi-gun, Korea, on September 11. The uniform fruits without any visible infection or injuries were selected for the experiment.
The selected fruits were assigned to four groups. In the 1-MCP group, the fruits were treated with 1.0 µL L‒1 1-MCP (SmartFreshTM, 3.3% w/w; Agrofresh, Yakima, WA, USA) for 18 h at 20°C (Win et al. 2019). In the SA group, the fruits were dipped in 3 mM SA solution for 5 min and then air dried for 6 h at 20°C. In the SA+1-MCP group, the fruits were dipped in SA solution first and then fumigated with 1-MCP. Untreated fruits were used as a control. Fruits from all groups were stored at 20 ± 1°C under 70–75% relative humidity for up to 5 weeks. This temperature was selected to simulate ambient shelf-life conditions during postharvest distribution and retail storage in Korea, where ‘Picnic’ apples are commonly marketed. Therefore, this period (5 weeks) represents a practical timeframe for evaluating fruit quality and physiological changes under typical room-temperature handling conditions.
Assessments of physiological characteristics
For internal ethylene concentration (IEC) determination, internal gas (1 mL) from the core cavity of each fruit was obtained and injected into a gas chromatograph (HP7820A-GC; Agilent, Wilmington, NC, USA) equipped with a Porapak Q column. During IEC measurement, the oven temperature was set at 90°C, and the temperatures of the injector and flame ionization detector were set at 200°C and 100°C, respectively. To determine the respiratory rate, the fruits were individually sealed in tightly closed plastic containers (1.6 L) for 1 h, and gas (1 mL) was collected using a syringe and injected into a gas chromatograph (GC-2010; Shimadzu, Kyoto, Japan) equipped with a Porapak Q column. During respiratory rate measurement, the oven temperature was set at 90°C, and the injector and detector temperatures were both set at 100°C. IEC was calibrated using an internal standard curve.
The fruit weights were recorded, and the weight loss of fruit was calculated as the difference in weight before and after storage (Win et al. 2019). Firmness (11 mm probe) was estimated using a firmness tester (COMPAC-100II; Sun Scientific, Tokyo, Japan). The starch pattern index (SPI) was scored according to the Cornell starch index test and scored from 1–8 scales (Blanpied and Sislsby 1992). Titratable acidity (TA) was measured using the malic acid equivalent method by titrating the juice sample with 1N NaOH until pH 8.1 (Win et al. 2019). The soluble solid content (SSC) was determined using a refractometer (PR-201α; ATAGO, Tokyo, Japan) with a refractive index accuracy of ±0.1% and a range of 0–45% with temperature correction. The peel color (L*, a*, and b*) was determined using a chroma-meter (CR-400; Konica Minolta, Tokyo, Japan). All physiological characteristics were assessed in 15 fruits per treatment. Each treatment consisted of three biological replicates, and each replicate represented the mean of five individual fruits. Therefore, a total of 15 fruits (5 fruits × 3 replicates) were used per treatment at each sampling time, and statistical analyses were conducted using three biological replicates. Flesh tissue samples were collected for further gene expression analysis.
RNA extraction and expression of ethylene-related genes
The extraction and expression analysis of the candidate genes were performed according to Ai et al. (2016). A RNAqueous kit was used to extract total RNA from tissue samples according to the manufacturer’s protocol. 1 µg of total RNA was used to synthesize cDNA using a reverse transcription kit (ReverTra Ace-a, Toyobo, Osaka, Japan) with an oligo dT20 primer. A real-time polymerase chain reaction PCR system (Thermo Fisher Scientific, Waltham, MA, USA) was used to analyze the expression of each gene, and Md-Actin was used as the internal control. The samples of the untreated fruit at harvest were used as a positive control, and the expression levels of genes were evaluated by the 2−ΔΔCT method. All analyses were performed in triplicate. The primers were selected based on previous studies (Yang et al. 2013; Singh et al. 2017). Primers and PCR conditions used in this study was described in Supplementary Table S1.
Statistical analysis
All data were analyzed using two-way analysis of variance (ANOVA), and the mean results of treatments were compared using least significant difference (LSD) test at p < 0.05 using SPSS version 25 software. All statistical analyses were performed using three biological replicates per treatment (each replicate = mean of 5 fruits). The correlation coefficient matrix system was analyzed using MetaboAnalyst 6.0 system (Chong et al. 2018).
Results
Ethylene production and respiration rate
The control and SA groups showed higher internal ethylene concentration (IEC) than those of the 1-MCP and SA+1-MCP groups (Fig. 1A). However, at 3–4 weeks of storage, the IEC and its peak value were lower in the SA group than in the control group. The IEC increase during storage was significantly inhibited by 1-MCP treatment. Similarly, the SA+1-MCP group showed low IEC during storage. However, the inhibitory effect of SA+1-MCP treatment on IEC was not greater than that of 1-MCP treatment alone (Fig. 1A).
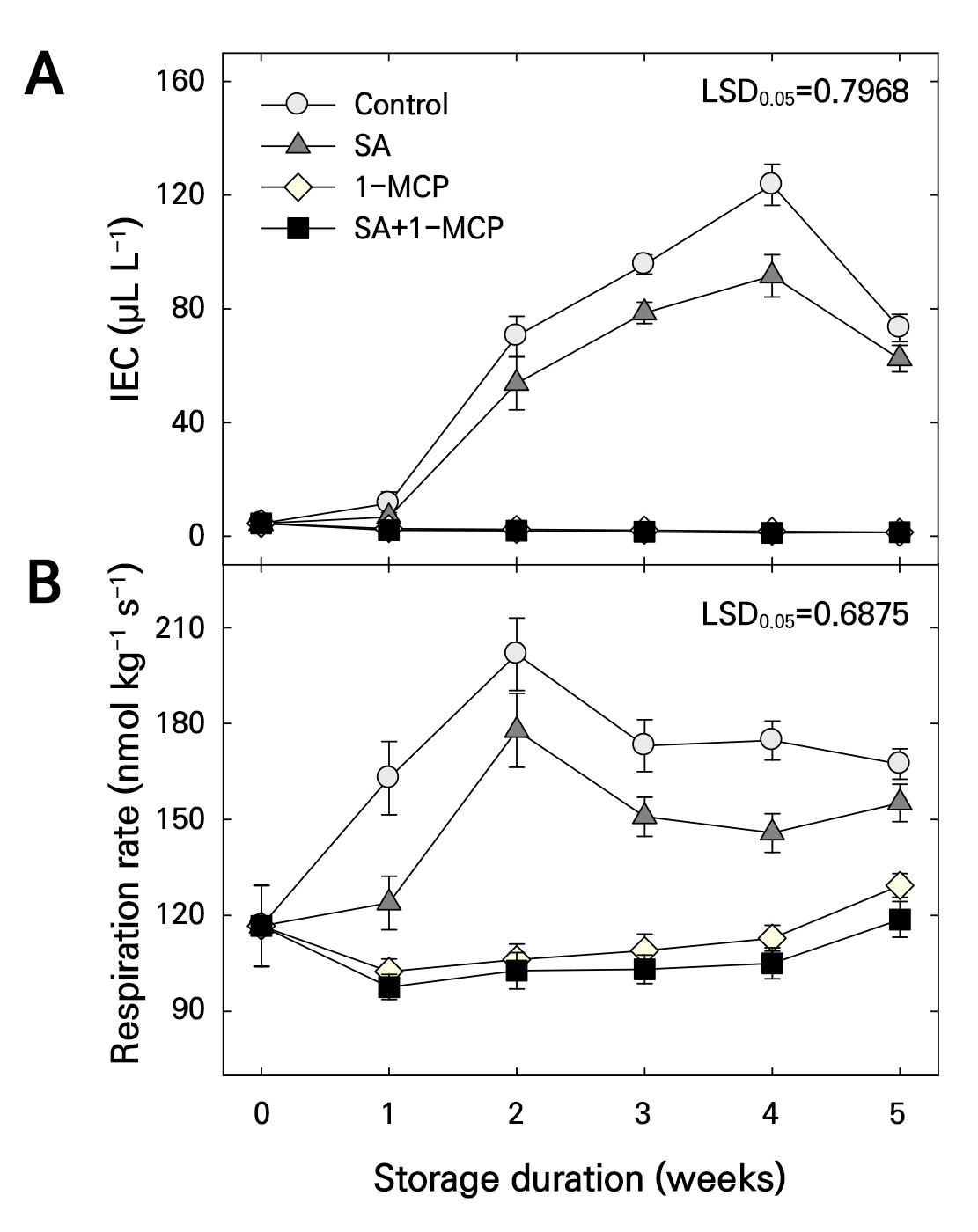
Fig. 1.
Internal ethylene concentration (IEC) (A) and respiration rate (B) of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.
The respiratory rate was higher in the control and SA groups than in the 1-MCP and SA+1-MCP groups during storage (Fig. 1B). However, at 1, 3, and 4 weeks of storage, the SA group showed lower respiratory rates than that of the control group. Further, the 1-MCP group showed significantly lower respiratory rates during storage. However, the SA+1-MCP group showed a similar respiratory rate to that of the 1-MCP group (Fig. 1B).
Firmness, titratable acidity (TA), and soluble solids content (SSC)
Firmness and TA were reduced in all treatment groups during storage (Fig. 2A and 2B). However, the loss of firmness and TA during storage was not maintained in the SA compared to that in the control group. The loss of firmness and TA during storage was significantly maintained in the 1-MCP group compared to that in the control and SA groups. Firmness and TA during storage were maintained by the SA+1-MCP treatment. Additionally, the SA+1-MCP treatment maintained firmness at higher levels than that maintained by 1-MCP treatment at 2 and 5 weeks of storage (Fig. 2A). In contrast, the 1-MCP and SA+1-MCP groups showed a similar TA (Fig. 2B). SSC was slightly increased in all treatment groups during storage (Fig. 2C). The SSC was higher in the control group than in the treatment groups at 2 and 4 weeks of storage. However, the SSC after SA treatment did not differ from that after 1-MCP or SA+1-MCP treatment (Fig. 2C).
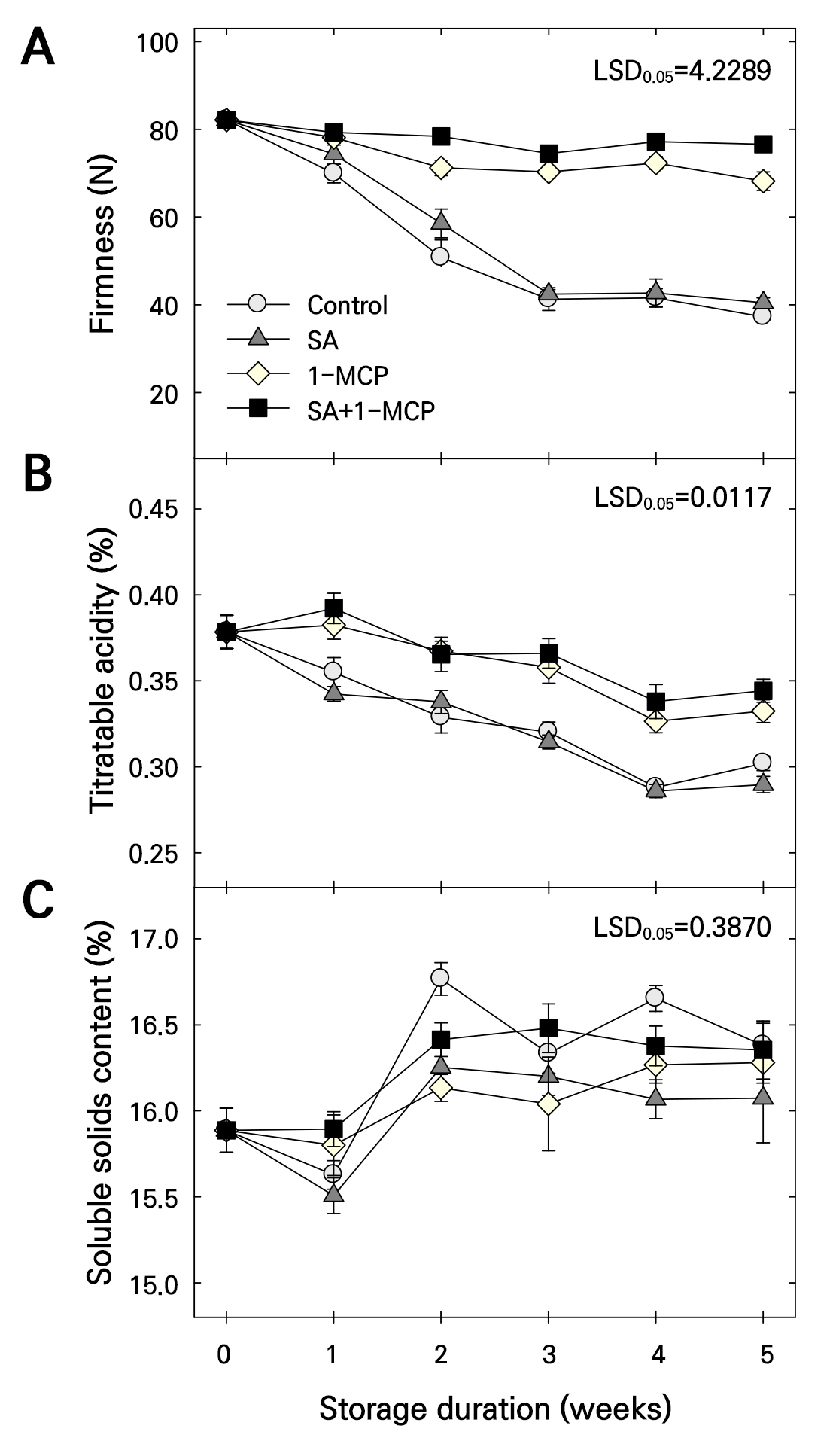
Fig. 2.
Firmness (A), titratable acidity (B), and soluble solids content (C) of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.
Weight loss and starch pattern index (SPI)
The weight loss increased in all treatment groups during storage (Fig. 3A). However, except at 4 weeks of storage, SA treatment did not reduce the weight loss compared with that of the control. Compared with the control and SA treatments, weight loss was significantly reduced by 1-MCP treatment at 3–5 weeks and after 5 weeks of storage, respectively. Moreover, weight loss was reduced by SA+1-MCP treatment; however, its effects were similar to that of 1-MCP treatment (Fig. 3A).
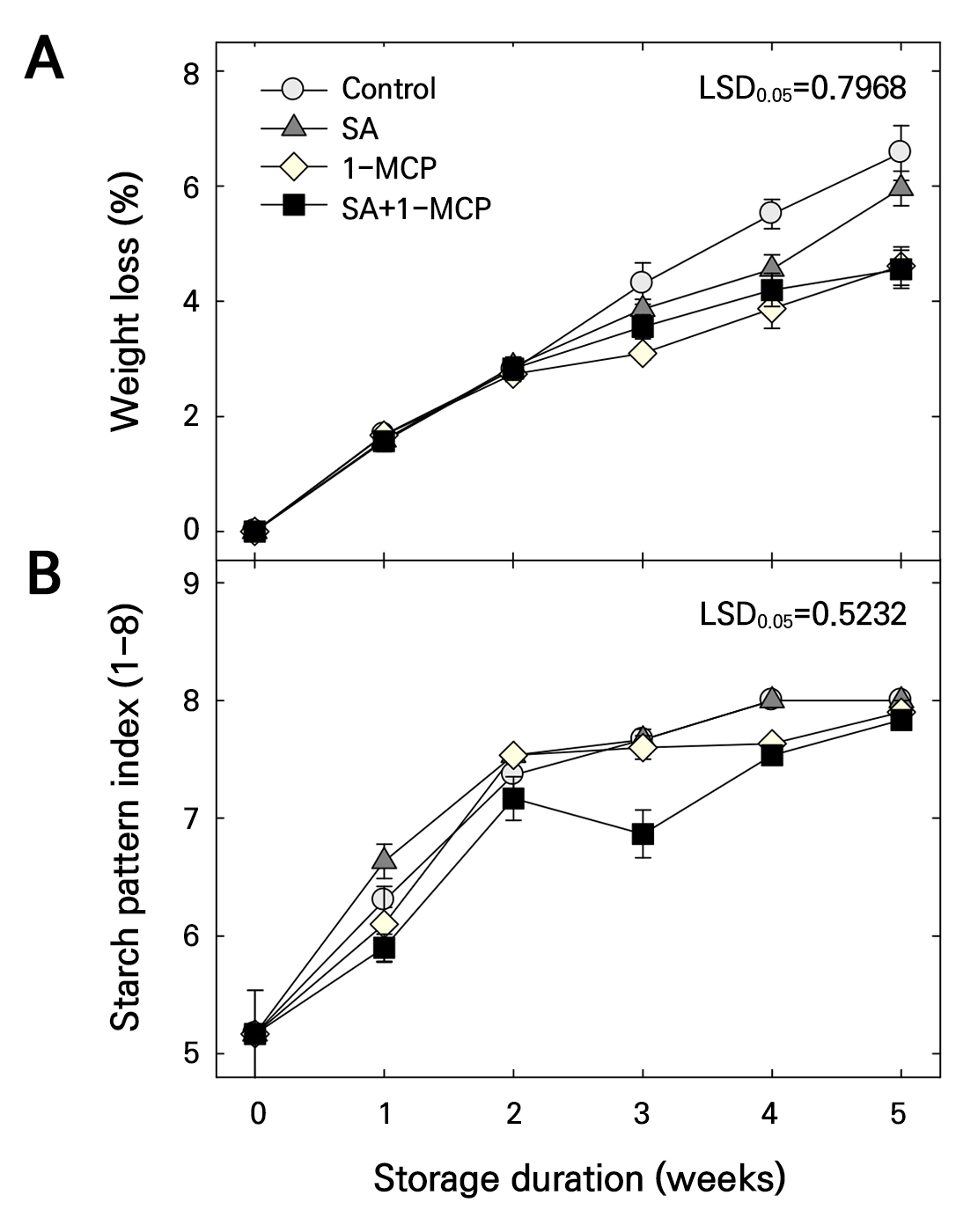
Fig. 3.
Weight loss (A) and starch pattern index (B) of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.
A slower reduction of the SPI score was observed in the SA+1-MCP group especially compared with the control until 3 weeks of storage (Figs. 3B and 4). However, at 4 and 5 weeks of storage, the SPI score did not differ between all treatments (Figs. 3B and 4).
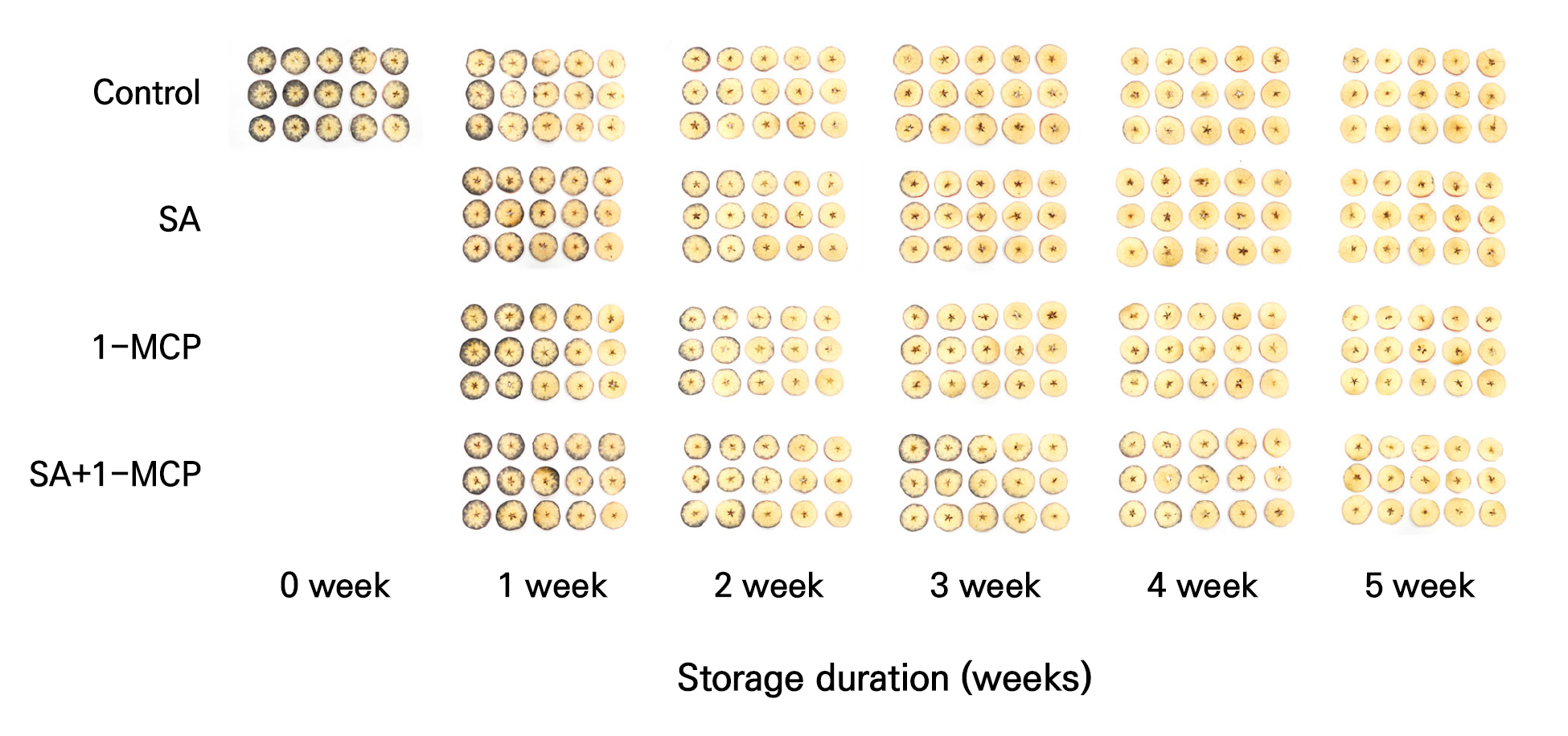
Fruit peel color and fruit appearance
Overall, the peel color (L*, a*, and b*) showed no consistent differences among treatments during storage (Fig. 5). However, after 4 weeks, fruits treated with SA exhibited slightly higher L* and b* values compared with the other treatments (Fig. 5A and 5C). No significant differences were observed in a* (redness) values across treatments (Fig. 5B). Accordingly, color differences were minimal and difficult to quantify, and all treatments maintained a similar visual appearance throughout storage (Fig. 6).
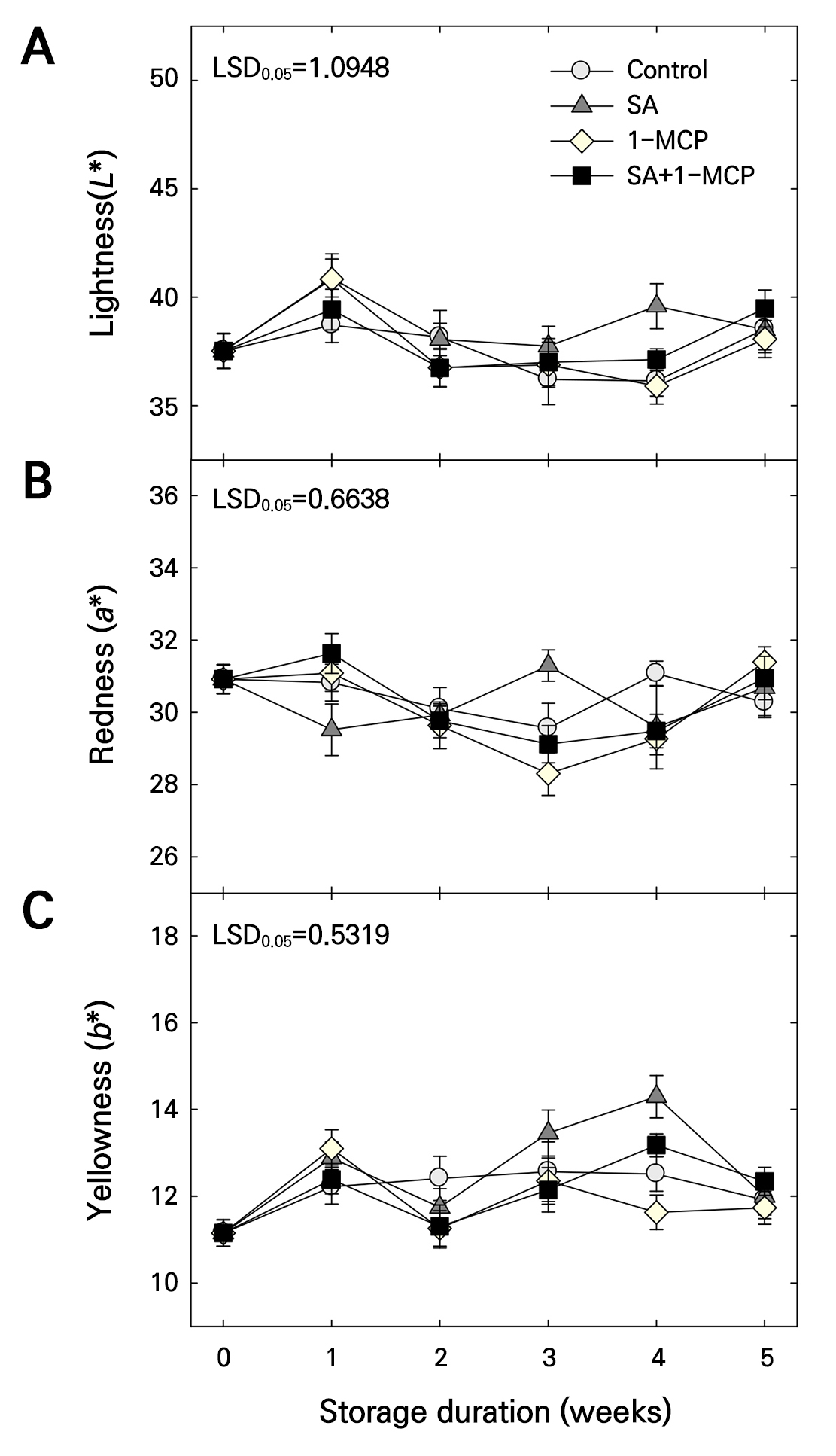
Fig. 5.
Fruit color L* (A), a* (B), and b* (C) of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.

Expression of ethylene biosynthesis and receptor genes
When the expression of ethylene biosynthesis genes was evaluated, MdACS1 expression was higher in the control group, especially at 4 and 5 weeks of storage, than that in the SA group. MdACS1 expression was also higher in the SA group than in the 1-MCP and SA+1-MCP groups (Fig. 7A). However, MdACS1 expression did not differ between the 1-MCP and SA+1-MCP groups (Fig. 7A). Higher MdACO1 expression was also observed in the control and SA groups than those of the 1-MCP and SA+1-MCP groups (Fig. 7B). Except at 5 weeks of storage, MdACO1 expression did not differ between the control and SA groups and between the 1-MCP and SA+1-MCP groups (Fig. 7B).
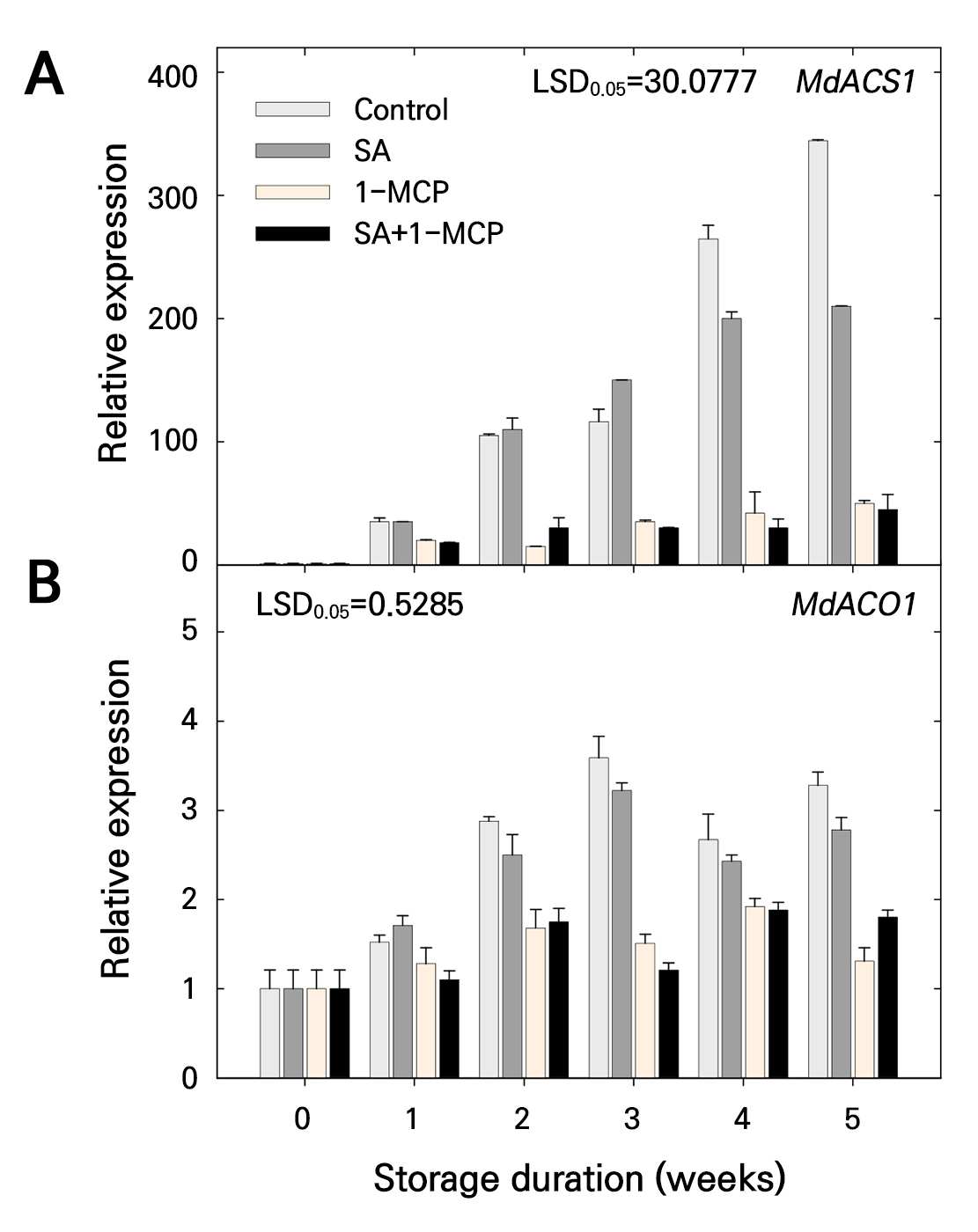
Fig. 7.
Relative expression of MdACS1 (A) and MdACO1 (B) genes of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.
Regarding ethylene receptor genes, the control and SA groups showed high MdETR1 expression, which was maintained until 4 weeks of storage (Fig. 8A). The control group showed higher MdERS2 expression, particularly at 1 and 4 weeks of storage, than those of the treatment groups (Fig. 8B). MdETR1 and MdERS2 were downregulated after 1-MCP treatment during storage when compared to those in the control and SA groups (Fig. 8A and 8B). The SA+1-MCP group showed lower MdETR1 and MdERS2 expression than those in the control and SA groups, although the expression was similar to that in the 1-MCP group (Fig. 8A and 8B). MdCTR1 expression was higher in the control and SA groups than those in the 1-MCP and SA+1-MCP groups (Fig. 8C). However, the MdCTR1 expression level did not differ between the control and SA groups and between the 1-MCP and SA+1-MCP groups (Fig. 8C).
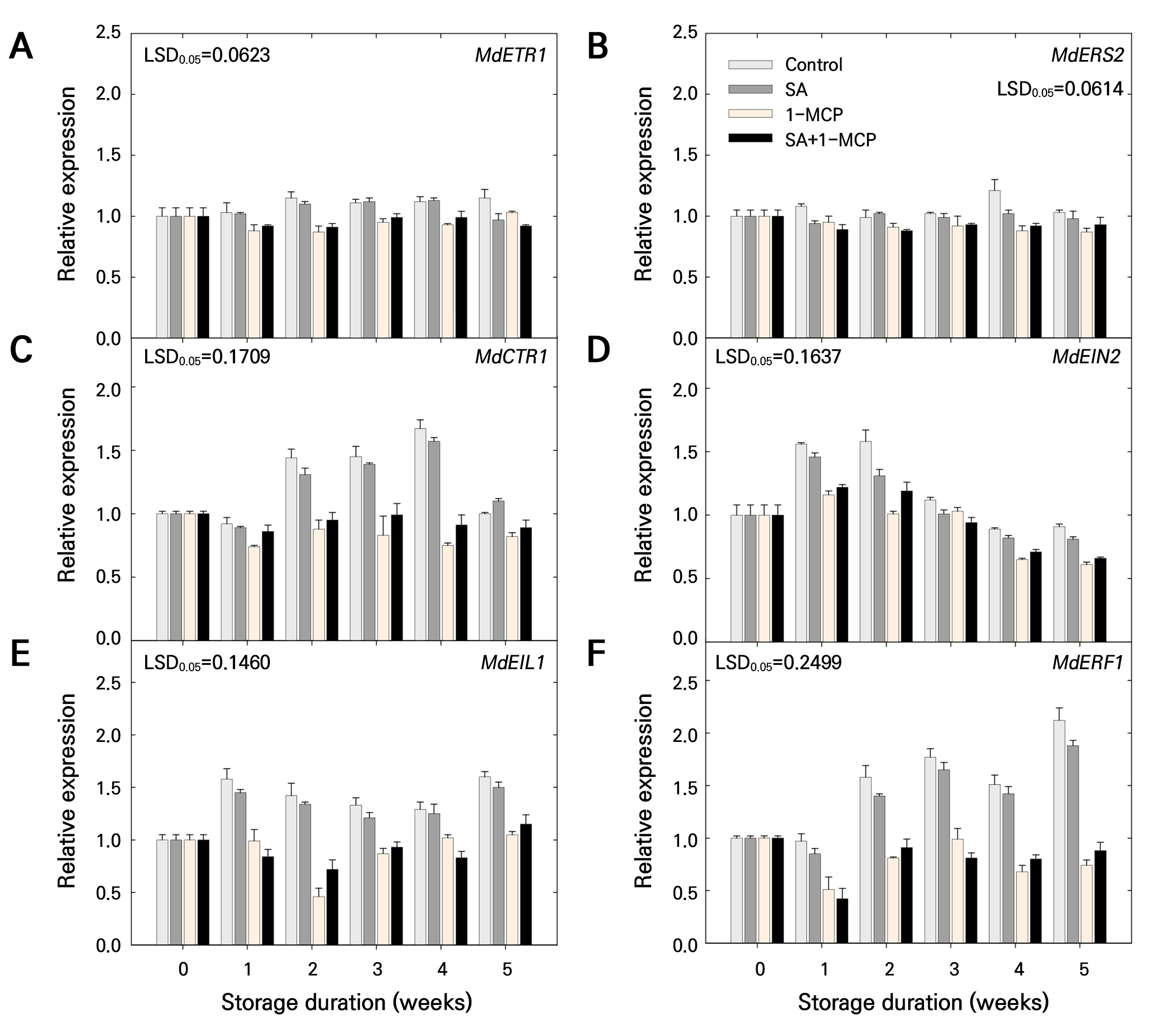
Fig. 8.
Relative expression levels of (A) MdETR1, (B) MdERS2, (C) MdCTR1, (D) MdEIN2, (E) MdEIL1, and (F) MdERF1, which are involved in ethylene receptors and the signal transduction pathway of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP and stored at 20 ± 1°C for 5 weeks. The bars indicate the mean ± standard error (n = 3). The least significant difference (LSD) test is applied to examine significant difference at p < 0.05.
Regarding ethylene signal transduction genes, MdEIN2 expression was lower in the SA group than that in the control group (Fig. 8D); however, no difference was observed in MdEIL1 and MdERF1 expression between these groups during storage (Fig. 8E and 8F). MdEIN2, MdEIL1, and MdERF1 were significantly downregulated in the 1-MCP group during storage compared to those in the control and SA groups. Additionally, ethylene signal transduction genes were downregulated in the SA+1-MCP group compared to those in the control and SA groups. However, this expression remained similar to that in the 1-MCP group (Fig. 8D–8F).
Correlation coefficient matrix
Based on the overall correlation coefficient matrix (Fig. 9), firmness was positively correlated with TA, but negatively correlated with both IEC and respiratory rate. SSC was positively correlated with the SPI, but negatively correlated with TA. IEC was positively correlated with respiratory rate and ethylene genes (MdACS1, MdACO1, MdETR1, MdERS2, MdCTR1, MdEIL1, and MdERF1), except for MdEIN2. Additionally, the ethylene biosynthesis and receptor gene expression was positively correlated with each other (Fig. 9).
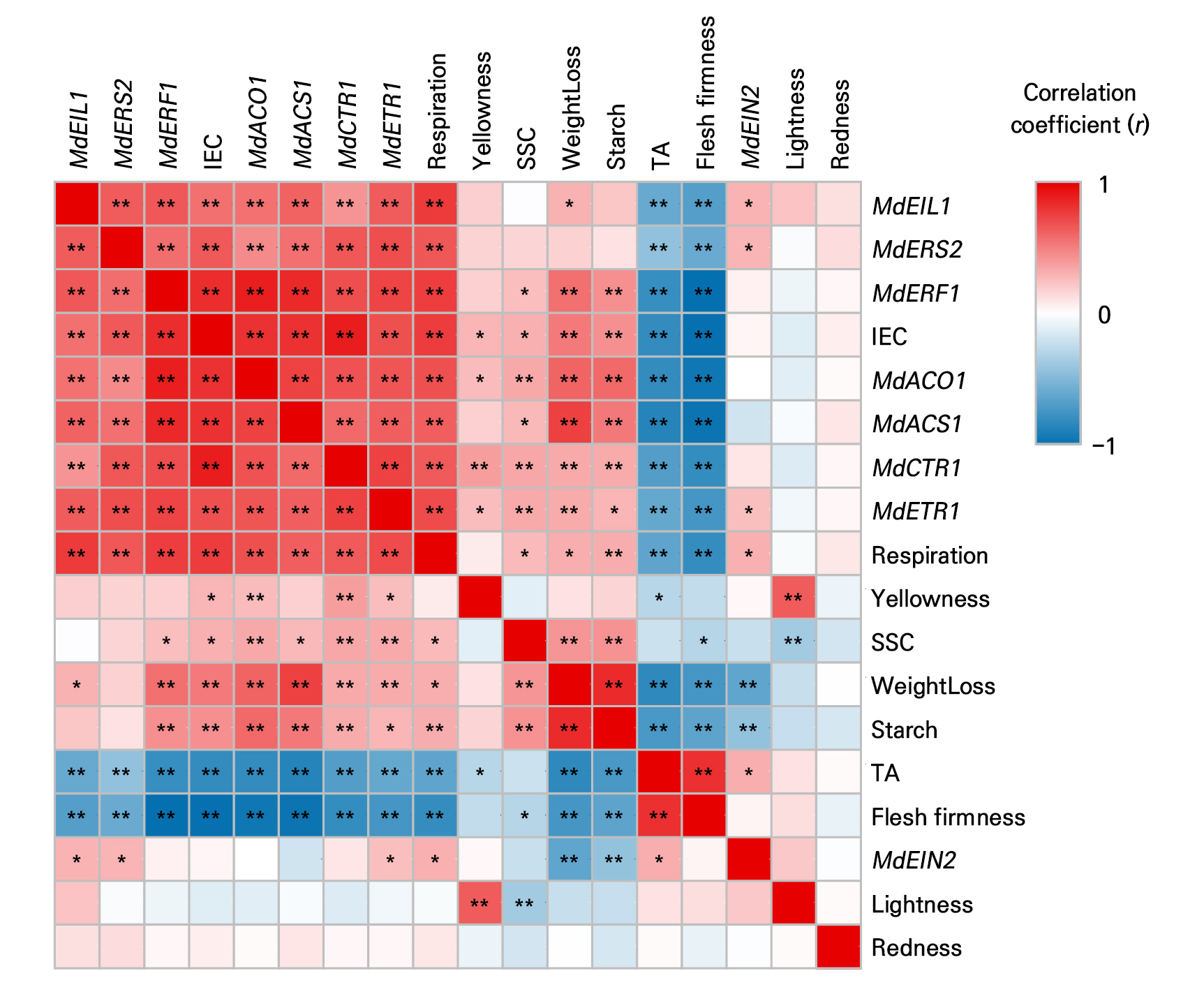
Fig. 9.
Correlation coefficient analysis of ‘Picnic’ apples treated with salicylic acid (SA), 1-methylcyclopropene (1-MCP), or SA + 1-MCP at harvest and stored at 20 ± 1°C for 5 weeks. * and ** indicate significant correlation levels at p < 0.05 and p < 0.01. IEC, internal ethylene concentration; SSC, soluble solids content; TA, titratable acidity.
Discussion
In this study, we assessed the effects of SA, 1-MCP, and their combination on fruit quality changes in ‘Picnic’ apples stored at 20°C. The IEC of the control group was higher than that of other treatment groups. Similarly, the respiratory rate of the control group was higher than that in other treatment groups. Additionally, IEC was strongly and positively correlated with respiratory rate during fruit ripening. However, the IEC and respiratory rate increased following SA treatment unlike in the 1-MCP and SA+1-MCP groups. 1-MCP treatment can reduce the IEC and respiratory rate in apples (Win et al. 2019), and at least 85% of ethylene production and 34% of the respiratory rate has been previously reduced by 1-MCP treatment (Zhang et al. 2020). In the current study, the 1-MCP group showed complete reduction in IEC and low respiratory rates. However, the SA group reduced the IEC and respiration rate of fruit, especially after 3 and 4 weeks of storage. Zhao et al. (2021) reported substantial ethylene production, although it may be inhibited by SA treatment, similar to that observed in our study. We also found that compared with 1-MCP treatment, ethylene production was not entirely blocked by SA treatment. The low IEC in the SA group may be due to the inhibition of ACC synthesis and ACC conversion to ethylene (Zhang et al. 2003; Li et al. 2022). In banana, 1-MCP+SA treatment shows maximum inhibition of ethylene production and respiratory rate during storage (Xu et al. 2018). These results differed from our results, and the inhibitory effect of SA+1-MCP treatment on IEC and respiratory rate was not greater than that of 1-MCP treatment.
Firmness is measured to evaluate fruit softening (Harker et al. 2008), and it generally decreases during ripening (Goulao and Oliveira 2007). TA showed a similar trend and was strongly and positively correlated with firmness. In the present study, SA+1-MCP showed the most effective result in maintenance of firmness and TA. Additionally, 1-MCP group maintained firmness and TA greater than that of SA group. 1-MCP treatment has been shown to maintain the reduction in firmness and TA in most apple cultivars; however, its effect varies depending on the cultivar (Kwon et al. 2024). Another study reported that the change in firmness and TA after SA treatment varies based on the fruit variety. Kim et al. (2019) revealed that firmness and TA in table grapes are maintained by SA+1-MCP treatment but not by SA or 1-MCP treatment alone. However, in wax apples, both 0.5 and 1.0 mM SA concentrations maintained firmness but had no effect on TA (Supapvanich et al. 2017). Baswal et al. (2020) reported that firmness and TA vary depending on the SA concentration in mandarin. Firmness is not affected in SA-treated peaches during storage (Zhao et al. 2021). Regarding the previous studies, the effects 1-MCP or SA treatments on the firmness and TA largely depends on the fruit crop, specific cultivar, concentration, and storage conditions. Additionally, Kazemi et al. (2011) reported that the SA-treatment-mediated reduction in fruit softening may be attributed to inhibition of ACO activity and decreased ACC conversion to ethylene. Furthermore, SA could reduce cell wall swelling and activate antioxidant enzymes (Shafiee et al. 2010). Therefore, a greater firmness was observed in the SA+1-MCP group, which may be due to the synergistic effect of SA and 1-MCP, as previously reported in apricots (Liang et al. 2023). Moreover, 1-MCP+SA treatment shows greater maintenance in firmness and TA in banana during storage (Xu et al. 2018), similarly observed in this study.
The SSC increased in the control group at 2 and 4 weeks of storage when compared to that in the control group in this study. The SSC may increase or decrease following 1-MCP treatment depending on the cultivar (Watkins 2006). SA treatment does not alter the SSC in table grapes and wax apples (Supapvanich et al. 2017); however, SA treatment reduces and increases SSC in bananas (Xu et al. 2018) and peaches (Zhao et al. 2021), respectively. Therefore, the effect of SA and 1-MCP treatments on SSC highly depends on the cultivar and storage conditions. In the present study, a slightly lower SSC was observed in the SA, 1-MCP, and SA+1-MCP-treated fruits than in the control fruits during storage. Therefore, the SSC could be reduced by applying SA or 1-MCP alone, and their combination had a similar effect on SSC.
Weight loss was reduced by 1-MCP treatment but not by SA treatment when compared with that in the control. Weight loss in the SA group reduced after 5 weeks of storage but remained similar to that in the 1-MCP group during storage when compared with that in the SA+1-MCP group. Weight loss has been delayed by 1-MCP treatment; however, this effect varies depending on the cultivar (Win et al. 2019; Zhang et al. 2020). Weight loss is affected by high and not low SA concentrations in wax apples (Supapvanich et al. 2017). In general, weight loss is determined based on fruit respiration and transpiration; however, the respiratory rate observed in our study was higher in the control group than in the SA group. Nonetheless, the weight loss response to SA treatment in the present study remains unclear. Weight loss did not differ between the SA+1-MCP and 1-MCP groups, which may be due to the similar fruit respiratory rates during storage. A slower reduction of the SPI was observed in the SA+1-MCP group. 1-MCP treatment can reduce the rate of starch hydrolysis during fruit maturation and ripening (Doerflinger et al. 2015).
Fruit appearance and peel color alterations were not affected by 1-MCP treatment. Additionally, peel color alterations were not strongly correlated with IEC in this study. 1-MCP treatment does not affect peel color alterations in ‘Honggeum’ apples (Yoo et al. 2020); however, the maintenance of color by 1-MCP treatment depends on the storage temperature and time of application (Zhang et al. 2020). SA treatment increases L* value in mango and fresh goji fruits compared to that in the control (Khedr 2016; Zhang et al. 2021). In the present study, high L* and b* values were observed in SA-treated fruits at 4 weeks of storage. Therefore, our findings revealed that 1-MCP treatment could not maintain fruit peel color during storage.
ACS and ACO represent the two key enzymes involved in the ethylene biosynthesis pathway; however, these enzymes are encoded by multi-gene families (Liu et al. 2015b). ACS1 and ACO1 were evaluated because their expression levels are strongly correlated with climacteric ethylene bursts during ripening (Wang et al. 2007). In this study, ACS1 and ACO1 expression was higher than that in the control, whereas their expression was reduced following 1-MCP and SA+1-MCP treatments. Thus, the upregulation of ACS1 and ACO1 in untreated fruits indicated that the regulation of these genes coincided with ethylene production (Win et al. 2025). A lower expression of ACS1 and ACO1 in 1-MCP group is an indicator of ethylene production inhibition by 1-MCP, which occurs at the transcript levels in the ethylene biosynthesis pathway (Wakasa et al. 2006; Win et al. 2025). A lower expression levels of ethylene biosynthesis genes in SA group might be related to the IEC production levels during storage. However, high ACS1 and ACO1 expression in the SA group may be due to still substantial of IEC during storage.
In the ethylene perception mechanism, ethylene receptors generally function as negative regulators of ethylene responses. Reduced ethylene production in 1-MCP-treated fruits may be due to the binding of 1-MCP to ethylene receptors, thereby decreasing the sensitivity of fruits to ethylene signaling (Wiersma et al. 2007; Costa et al. 2010). Additionally, Tatsuki and Endo (2006) reported that ethylene is required for the expression of these receptor genes; however, their expression does not depend on the ethylene production level. A strong relationship was observed between IEC and receptor gene expression. Similarly, in our study, MdETR1 and MdERS2 expression was largely unchanged in all treatment groups. However, their expression was higher in the control group than in the 1-MCP group. This result is consistent with that of previous studies. ETR1 and ERS2 expression reduced in apples after 1-MCP treatment indicating that 1-MCP binds to ethylene at receptor-binding sites (Tatsuki et al. 2009). Ethylene receptors closely interact with downstream CTR1, which also negatively regulates the ethylene response (Park et al. 2023). However, CTR1 expression is induced by ethylene, and it is upregulated during ripening, as previously reported in apples (Dal Cin et al. 2006). 1-MCP treatment can reduce CTR1s expression in apples (Yang et al. 2013), as observed following 1-MCP and SA+1-MCP treatment. Interestingly, the SA treatment did not noticeable reduce the gene expression levels in the receptor side, except some lower expression levels in the MdERS2 gene. High expression of receptor genes in SA treated fruits would be probably due to the higher IEC that facilitates binding ethylene receptors during ripening.
EIN2 transmits ethylene signals from receptors to the nucleus. EIN2 is closely involved in the regulation of the fruit ripening process in tomatoes, and ripening-related genes are downregulated in response to EIN2 downregulation (Hu et al. 2010). EIN2A and EIN2B expression is not regulated by ethylene, but is reduced after 1-MCP treatment in apples (Yang et al. 2013). Similarly, we found that MdEIN2 expression was reduced by 1-MCP and SA+1-MCP treatments during storage. Solano et al. (1998) reported that EIN3/EILs function as positive regulators of ethylene signaling, but their functions differ between distinct family genes or tissues. The inhibition of EIL1 expression by 1-MCP treatment has been previously reported in mulberry and durian (Liu et al. 2015a; Thongkum et al. 2018); the expression level is similar in 1-MCP- and SA+1-MCP-treated fruits. In the ethylene signaling pathway, ERF1 activation represents the last step of signal transduction in the nucleus (Solano et al. 1998).
Wang et al. (2007) revealed that high ERF1 expression is likely due to the high level of ethylene production, as observed in control fruits in the current study. Hence, the low ERF1 expression in 1-MCP-treated apples indicated that ERF1 expression is regulated by ethylene (Wang et al. 2007; Yang et al. 2013). Similarly, a strong relationship between IEC and ERF1 expression was observed in our study. Additionally, a significant response to ERF1 expression was observed at the late storage time, especially in the control and SA groups. Because the expression of ethylene receptor genes was not apparently reduced by SA treatment, high EIN2, EIL1, and ERF1 expression, which can induce ethylene signaling in fruits during ripening, was also observed. The high expression of ethylene receptor and signal transduction genes in SA group may be due to the high IEC, which allows the binding of ethylene to its receptors during ripening. Therefore, the effect of SA treatment on the ethylene production mechanism was not prominently observed at the receptor-binding sites in this study.
In conclusion, 1-MCP treatment reduced the IEC, respiratory rate, and expression of candidate genes involved in the ethylene perception mechanism in ‘Picnic’ apples. SA treatment had less effects on ethylene inhibition and fruit quality maintenance than that of 1-MCP treatment. SA+1-MCP treatment did not exhibit more potent effects on ethylene inhibition mechanisms than that of 1-MCP treatment. However, a combination of SA+1-MCP treatment showed some greater effects on fruit quality maintenance than that of 1-MCP. Altogether, our study revealed that the application of 1-MCP is economically viable for ethylene inhibition mechanisms of apple, but a combination of SA+1-MCP is recommended for fruit quality maintenance of ‘Picnic’ apples during ambient temperature storage. Further research is warranted to elucidate the role of SA in the apple ripening mechanism during postharvest storage.
Supplementary Material
Supplementary materials are available at Horticultural Science and Technology website (https://www.hst-j.org).
- HORT_20250083_Table_S1.pdf
Supplementary Table S1. Primers information used in the expression analysis of ethylene biosynthesis and perception genes
