

서 언
재료 및 방법
식물 재료
DNA 추출
염기서열 분석용 프라이머 디자인
염기서열 분석 및 유전자 변이 탐색
ZEP 및 CCS 유전자 내 염기서열 변이 구분용 프라이머 디자인
HRM 분석
결과 및 고찰
후보유전자의 염기서열 분석 및 변이 탐색
ZEP 및 CCS 유전자 기반 분자표지 개발
개발된 분자표지의 효용성 검정
CCS 및 ZEP 유전자형 조합과 파프리카 과색
서 언
국내에서 파프리카(Capsicum annuum L.)는 착색단고추(colored sweet pepper)를 의미하며 과형에 따라 블로키, 미니, 코니컬 등으로 구분된다(Lee et al., 2010). 파프리카는 크기, 과색, 광택성, 경도, 단맛 등에 의해 품질이 결정되며, 특히 숙과색은 파프리카를 구매하는데 주된 결정요인으로 작용한다(Lee et al., 2016; Kim et al., 2018; Byeon et al., 2023; Nie et al., 2023).
카로티노이드(carotenoid)는 식물의 2차 대사산물 geranylgeranyl pyrophosphate(GGPP) 두 분자가 phytoene synthase (PSY)에 의해 결합을 하여 phytoene이 합성되며, 이후 여러 생합성 효소들에 의해 다양한 카로티노이드 물질로 합성이 되어 식물의 과실, 태좌, 종자, 뿌리 등에 축적되는 것으로 알려져 있다(Fig. 1; Ruiz-Sola and Rodríguez-Concepción, 2012; Hassan et al., 2019; Villa-Rivera and Ochoa-Alejo, 2020). 이러한 카로티노이드는 종류에 따라 다양한 건강 기능성 효과가 있다(Hassan et al., 2019; Villa-Rivera and Ochoa-Alejo, 2020). 파프리카에서 카로티노이드의 축적은 빨간색, 노란색, 주황색의 주요 성숙 과색을 결정짓는 주된 요인으로 작용한다(Havaux, 2014; Hassan et al., 2019; Hermanns et al., 2020; Kye et al., 2022).
고추(Capsicum annuum L.)의 다양한 숙과색은 카로티노이드 생합성에 관여하는 c1, c2 및 y 유전자좌에 의한 것이라고 보고되었다(Hurtado-Hernandez and Smith, 1985). 이 후 연구에서 고추의 숙과색은 phytoene synthase 1 유전자(PSY1, c2 locus on chromosome 4), capsanthin-capsorubin synthase 유전자(CCS, y locus on chromosome 6), pseudo-response regulator 2 유전자(PRR2, c1 locus on chromosome 1), zeaxanthin epoxidase 유전자(ZEP, chromosome 2) 및 β-carotene hydroxylase 유전자(BCH, chromosomes 3 and 6)의 영향을 받는 것으로 보고되었다(Fig. 1; Lefebvre et al., 1998; Popovsky and Paran, 2000; Huh et al., 2001; Jeong et al., 2020; Lee et al., 2020a; Lee et al., 2021). 그리고 phytoene synthase 2 유전자(PSY2, chromosome 2)는 PSY1 유전자와 동일한 기능을 할 수 있는 것으로 보고되었다(Jang et al., 2020).
PSY1, CCS 및 PRR2 유전자가 모두 정상이면 고추 숙과색은 빨간색이 되고, 이들의 여러 유전자형 조합에 따라 다양한 숙과색이 나타나게 된다(Hurtado-Hernandez and Smith, 1985). PSY1 유전자의 기능상실은 고추 숙과색을 주황색으로 만든다(Huh et al., 2001; Kim et al., 2010). CCS 유전자의 손상은 고추 과색을 주황색 또는 노란색으로 만든다(Lefebvre et al., 1998; Popovsky and Paran, 2000; Lang et al., 2004; Kim et al., 2017). 그리고 CCS의 다양한 대립유전자 변이가 보고되었다(Naresh et al., 2012; Jeong et al., 2019; Omori and Sasanuma, 2022). PRR2 유전자의 기능상실은 전체적인 카로티노이드 함량을 낮추는데, PSY1, CCS 및 PRR2 세 유전자 모두 기능상실 대립유전자를 가지고 있으면 고추 과색은 흰색이 된다(Jeong et al., 2020; Lee et al., 2020a). CCS 유전자의 기능이 없을 때, ZEP 유전자의 손상은 노란색 과실을 주황색으로 만든다고 보고하였다(Lee et al., 2021). EMS 처리를 통한 BCH 유전자 돌연변이체에서 베타카로틴(β-carotene)의 축적과 빨간색에서 주황색으로의 과색 변화를 보고하였다(Borovsky et al., 2013). 또한 분리된 고추 열매에서 카로티노이드 생합성 유전자의 바이러스 유도 유전자 침묵(virus-induced gene silencing, VIGS) 실험은 PSY1 및 CCS 유전자 뿐 아니라 LCY-B 및 BCH 유전자 침묵에서도 주황색 또는 노란색으로의 과색 변화를 일으켰다(Tian et al., 2014).
고추 숙과색에 따라 카로티노이드 종류별 함량에 차이가 있다(Hassan et al., 2019; Hermanns et al., 2020). 빨간색에서는 캡산틴(capsanthin)과 캡소루빈(capsorubin)의 함량이 높게 나타났으며, 노란색에서는 바이올락잔틴(violaxanthin)과 루테인(lutein)의 함량이 높았으며, 주황색에서는 베타카로틴(β-carotene)과 제아잔틴(zeaxanthin)의 함량이 높은 것으로 보고되었다(Fig. 1; Kilcrease et al., 2013; Berry et al., 2019; Hassan et al., 2019; Hermanns et al., 2020).
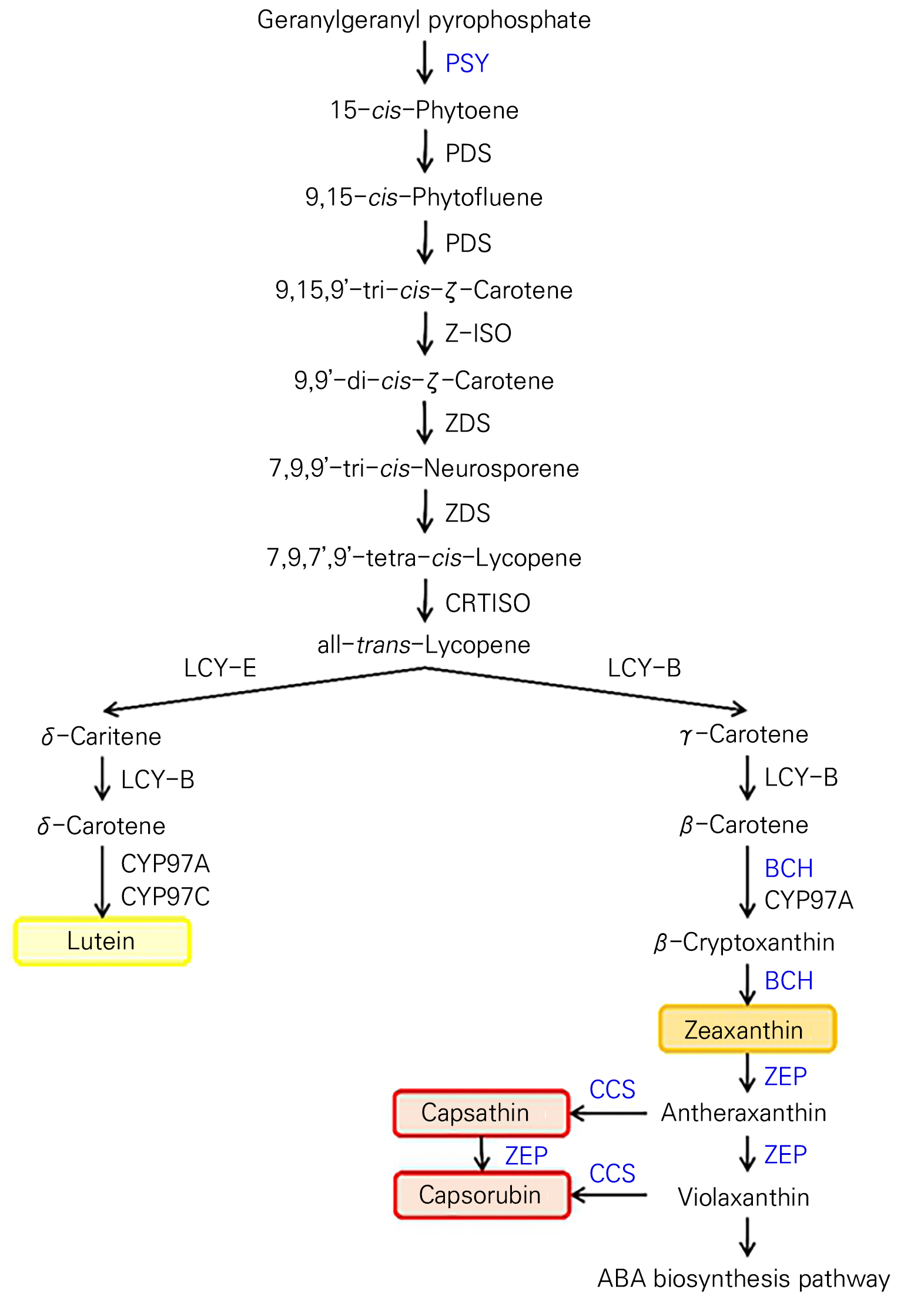
Fig. 1.
Carotenoid biosynthetic pathway in pepper. PSY (phytoene synthase), PDS (phytoene desaturase), Z-ISO (ζ-carotene isomerase), ZDS (ζ-carotene desaturase), CRTISO (carotene isomerase), LCY-B (β-lycopene cyclase), LCY-E (ε-lycopene cyclase), BCH (β-carotene hydroxylase), CYP97A and B (β-carotene hydroxylase cytochrome 450 types A and B), ZEP (zeaxanthin epoxidase), and CCS (capsanthin-capsorubin synthase). Red paprika contains high levels of capsanthin and capsorubin (red box). Orange paprika contains a high level of zeaxanthin (orange box). Yellow paprika contains a high level of lutein (yellow box).
빨간색 파프리카의 주요 카로티노이드는 고추와 동일한 캡산틴과 캡소루빈이었으나, 노란색과 주황색 파프리카는 고추와 주요 카로티노이드가 다른 것으로 보고되었다(Fig. 1; Kim et al., 2016; Kye et al., 2022). 노란색의 경우 루테인의 함량이 가장 높고, 주황색의 경우 제아잔틴과 루테인의 함량이 높은 것으로 보고되었다(Fig. 1; Kim et al., 2016; Kye et al., 2022).
하지만 앞에서 설명하였던 여러 유전자들 중에서 파프리카의 세 가지 과색(빨강, 주황, 노랑)을 결정하는 유전자와 그 대립유전자의 변이에 대한 연구는 아직 이루어지지 않은 상황이다.
따라서 본 연구에서는 후보유전자 접근 방법을 이용하여 파프리카의 카로티노이드 합성에 관여하는 유전자인 PSY, β-carotene hydroxylase(BCH), ZEP, CCS에서 염기서열 변이를 탐색하고, 세 가지 파프리카 숙과색을 구별할 수 있는 분자표지를 개발하고자 하였다.
재료 및 방법
식물 재료
식물 재료로 44개의 파프리카 F1 품종, 31개의 내혼계(inbred lines), 31개의 ‘GINA’ F2 식물체, 79개의 ‘JBR-10’ F2 식물체 및 380개의 유전자원을 사용하였다. 파프리카는 전라북도 군산시 대야면 전북농업기술원 과채류연구소 파프리카시험장에서 재배하였으며, DNA 추출을 위해 잎을 채취하여 사용하였다. 후보유전자의 염기서열을 분석하기 위해서 유전적 배경과 숙과색이 다른 10개의 F1 품종을 선발하여 사용하였으며 선발한 품종은 표1과 같다(Table 1). 다형성 검정에는 숙과색 표현형이 확실한 44개의 F1 품종을 이용하였으며, 이후 연관성 검정에는 내혼계, F2 집단 및 유전자원을 사용하였다. 과색은 불연속변이로 나타나 육안으로 조사하였으며 색깔에 따라 빨간색, 노란색, 주황색으로 구분하였다(Suppl. Fig. S1).
Table 1.
List of paprika materials used for genetic variation identification
DNA 추출
샘플링한 파프리카 잎으로부터 Lee et al. (2020b)에 의한 방법에 따라 DNA를 추출하였다. 추출된 DNA는 BioDrop Lite(Biochrom, England)를 이용하여 DNA 농도를 측정하여 20ng·µL-1로 희석하여 사용하였다.
염기서열 분석용 프라이머 디자인
염기서열 분석용 프라이머를 디자인하기 위해서 National Center for Biotechnology Information(https://www.ncbi.nlm.nih.gov/)에서 카로티노이드 생합성 유전자인 PSY, CCS, ZEP 및 BCH의 염기서열을 확보하였다(Table 2). 확보된 염기서열을 이용하여 Sol Genomics Network(http://www.sgn.cornell.edu/)의 Capsicum annuum cv. CM334(ver. 1.55) 표준유전체에서 유전자를 탐색하였고, 탐색된 유전자의 위치를 알아냈다(Table 2).
선발된 유전자들은 Primer 3 Input(ver. 0.4.0) (https://bioinfo.ut.ee/primer3-0.4.0/)과 Basic Local Alignment Search Tool(BLAST, https://blast.ncbi.nlm.nih.gov/)을 이용하여 프라이머 쌍을 포함하여 700–800bp의 amplicon이 특이적으로 증폭되도록 디자인하였다(Suppl. Table S1).
염기서열 분석 및 유전자 변이 탐색
Table 1에 나열된 식물재료의 DNA와 Table 3의 프라이머를 사용하여 PCR 산물을 만들었다. PCR 과정은 Lee et al. (2022)의 방법과 동일하게 사용하였다. PCR 산물은 1% 아가로스 젤에서 전기영동 후, IncloneTM Gel and PCR purification kit(Inclone Co., Ltd., Seongnam, Korea)를 이용하여 PCR 산물을 분리·정제하였다. 정제된 PCR 산물은 direct sequencing 방법으로 ㈜바이오닉스(Seoul, Korea)에서 수행되었다. 최종 확보된 염기서열은 ClustalX ver. 1.83을 이용하여 alignment하여 유전자 변이를 탐색하였다.
Table 2.
List of candidate genes selected for DNA sequencing
Table 3.
Primer sets for marker development information
ZEP 및 CCS 유전자 내 염기서열 변이 구분용 프라이머 디자인
ZEP 유전자 내에서 발견한 SNP를 확인할 수 있는 HRM 분석용 프라이머를 디자인하였다(Table 3). 확인된 SNP 주변 염기서열을 이용하여 Primer 3 Input(ver. 0.4.0)에서 프라이머를 디자인하였고, NCBI 사이트(https://blast.ncbi.nlm.nih.gov/)에서 amplicon을 BLAST하여 서열 특이적인지 확인하였다.
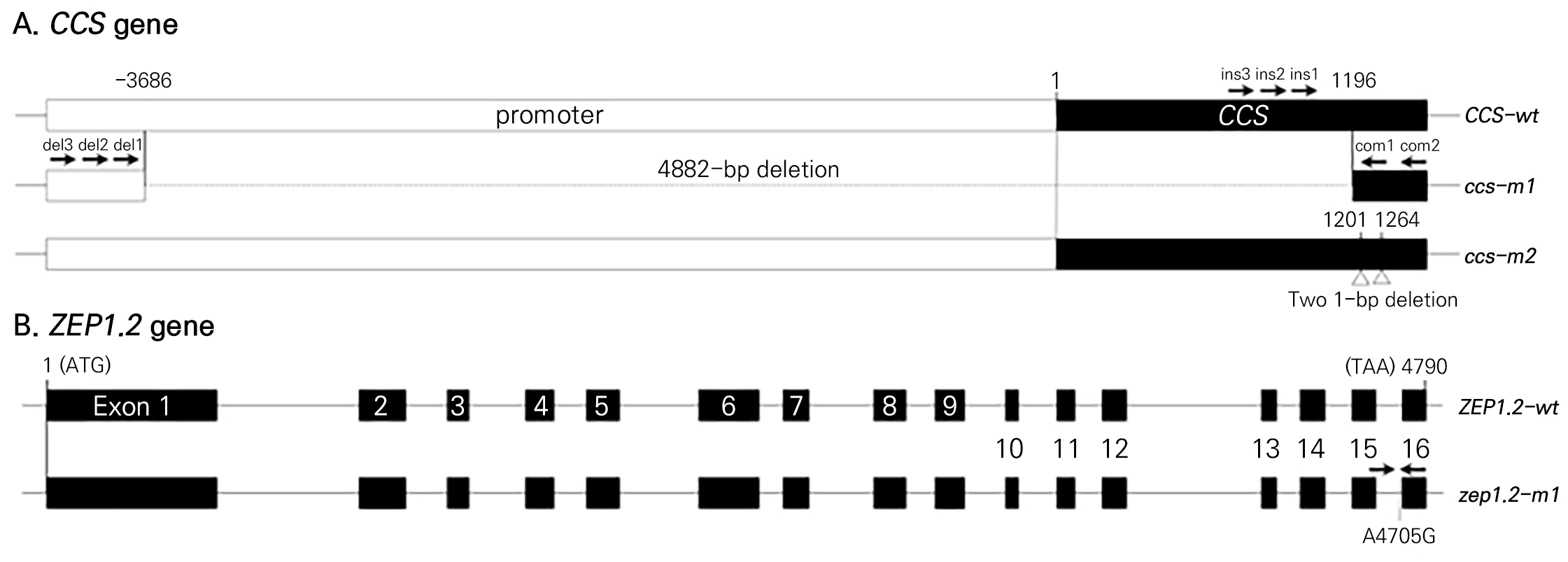
CCS 유전자 내에서 발견한 indel을 확인할 수 있는 HRM 분석용 프라이머를 위와 같은 방법으로 디자인하였다(Table 3). 정상 유전자 및 결실(deletion) 유전자에 각각 3개씩의 정방향(forward) 특이적 프라이머를 디자인하였고, 공통적으로 존재하는 엑손 부위에 2개의 역방향(reverse) 프라이머를 디자인하였다(Fig. 2 and Table 3).
HRM 분석
HRM 분석은 LightCycler® Real-Time PCR(Roche, Swizerland)을 사용하여 분석을 진행하였다. HRM 반응액 조성, PCR 방법 및 melting curve 분석 방법은 Lee et al. (2022)의 방법에 따라 수행하였다.
결과 및 고찰
후보유전자의 염기서열 분석 및 변이 탐색
총 9개의 후보유전자에 대해 염기서열 분석을 진행하여 과색별 염기서열 차이가 있는지 확인하였다(Table 1). 그 중 6번 염색체 위의 CCS 유전자와 2번 염색체 위의 ZEP1.2 유전자에서만 염기서열 변이를 탐색할 수 있었다(Fig. 2).
CCS 유전자는 빨간색 파프리카에서만 PCR 증폭이 되었으며 주황색과 노란색의 파프리카에서는 PCR 증폭이 되지 않았다. 따라서 주황색 및 노란색의 파프리카에서는 CCS 유전자에 결실(deletion)이 있을 것이라 추정하였다. 이전 연구에서 네 가지 종류의 CCS 돌연변이 대립유전자(ccs-p1, ccs-p2, ccs-p3 및 ccs-in)가 보고되었다(Jeong et al., 2019). 그 중에서 ccs-p1 대립유전자에 유전자 결실이 있어 이를 기준으로 다시 프라이머를 디자인하여 PCR 수행해 보았는데 PCR 증폭이 되었다. PCR 산물을 염기서열 분석하였는데 4882bp 결실을 확인하였다(Fig. 2). 기존 연구에서는 4879bp 결실을 보고하였는데 본 연구의 결과와 3bp 차이가 있었다. 이는 다른 대립유전자이기 때문인지 분석 실수 때문인지는 추가 확인이 필요해 보인다.
ZEP 유전자에 대해서는 빨간색, 주황색, 노란색 모든 파프리카에서 성공적으로 염기서열 분석을 완료하였다. 그 결과, 4790-bp 크기의 ZEP 유전자 내 4705-bp 위치에서 한 개의 SNP만 발견하였다(Fig. 2). 노란색 파프리카에서는 모두 A 염기서열이었고, 주황색 파프리카에서는 모두 G 염기서열이었다. 그러나 빨간색 파프리카에서는 품종에 따라 A 또는 G 염기서열을 가지고 있었다. 이 SNP 위치는 ZEP 유전자의 15번째 인트론(intron)에 위치하는 것인데, 인트론이 끝나는 신호인 AG 염기서열의 첫번째 염기에 해당하는 위치였다(Fig. 2). 따라서 이 염기서열의 SNP는 스플라이싱(splicing)에 영향을 미칠 것으로 생각되었다. 노란색 파프리카에서는 A 염기서열이기 때문에 정상적인 AG 염기서열이 만들어져 정상적인 스플라이싱이 일어날 것이라고 생각되었고, 주황색 파프리카에서는 G 염기서열이기 때문에 GG 염기서열이 만들어져 정상적인 스플라이싱이 일어나지 않을 것이라고 추정되었다. 이 결과는 Lee et al. (2021)의 이전 연구와 일치하는 결과였다.
ZEP 및 CCS 유전자 기반 분자표지 개발
노란색과 주황색 파프리카의 CCS 유전자는 프로모터와 유전자 부위를 포함하여 4882-bp의 결실이 있었다(Fig. 2). 이러한 결실로 인하여 CCS 유전자가 정상적인 기능을 하지 못할 것으로 생각되었다(Jeong et al., 2019). 이전 연구에서 CCS 유전자를 탐색하는 분자표지인 CCS524+CCS525의 경우 우성 SCAR(sequence characterized amplified region) 분자표지로 CCS 유전자의 이형접합형을 구분할 수 없었다(Popovsky and Paran, 2000). 최근 연구에서는 CCS 유전자의 이형접합형을 구분할 수 있는 공우성 SCAR 분자표지를 개발하였으나 아가로스 젤 전기영동을 이용하여 분석하는 방법을 사용하였다(Omori and Sasanuma, 2022). 본 연구에서는 CCS 유전자의 정확한 유전자형을 구분하기 위해 결실이 되지 않은 프로모터 부위(CCS-del1-F, CCS-del2-F, CCS-del3-F)와 결실부위(CCS-ins1-F, CCS-ins2-F, CCS-ins3-F)에 각각 3개의 정방향(forward) 프라이머를 디자인하였고, 결실부위 밖 엑손(exon) 영역에 공통적인 역방향(reverse) 프라이머(CCS-com1-R, CCS-com2-R)를 사용하여 실험을 진행하였다(Fig. 2 and Table 3). 이러한 프라이머 중 세 개의 프라이머를 이용한 다양한 조합에 대해 HRM을 분석한 후 melting curve 결과를 봤을 때, CCS 유전자형의 분리가 명확하게 나타난 CCS-com1-R, CCS-ins3-F 및 CCS-del3-F 프라이머 조합을 선발하였다(Suppl. Fig. S2). 이 조합으로 분석한 HRM 분자표지를 CCS-indel-HRM으로 명명하였고, 이후 CCS 유전자형 분석은 CCS-indel-HRM 분자표지를 이용하여 분석하였다(Fig. 3A and Suppl. Fig. S3). Fig. 3A에서 melting 온도가 78–79°C 사이에서 peak가 보이는 것은 정상 대립유전자인 Ccs를 가지고 있는 것을 나타내고, melting 온도가 85–86°C 사이에서 peak가 보이는 것은 결실 대립유전자인 ccs를 가지고 있는 것을 나타낸다(Fig. 3A). 이형접합체(heterozygote)는 두 군데 모두 peak를 보였다(Fig. 3A).
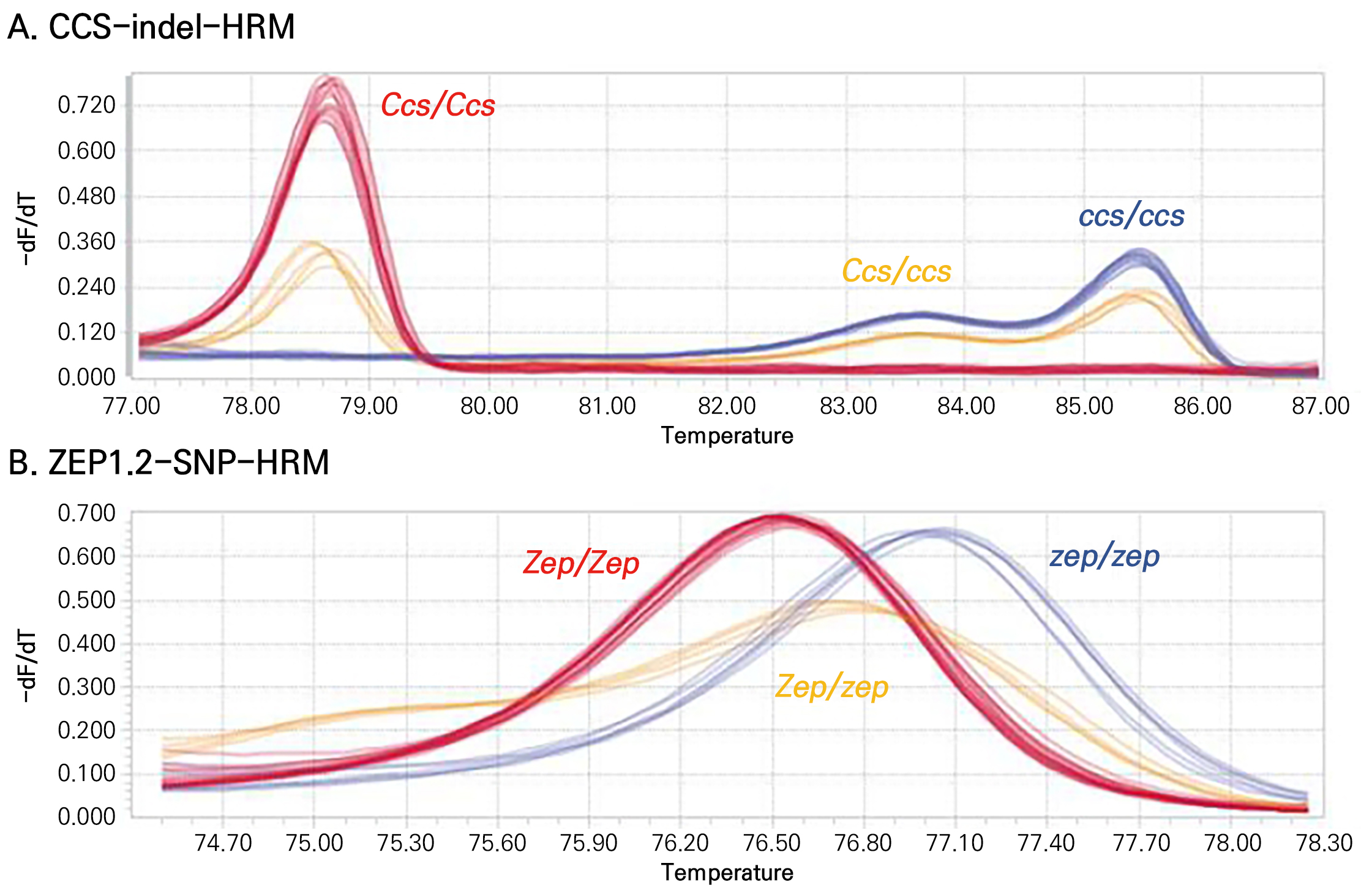
Fig. 3.
Melting curves of CCS-indel-HRM (A) and ZEP1.2-SNP-HRM (B) markers. Ccs, dominant normal allele; ccs, recessive mutant allele; Zep, dominant normal allele; zep, recessive mutant allele; Ccs/Ccs or Ccs/ccs, red fruits; ccs/ccs, orange or yellow fruits; Zep/Zep or Zep/zep, yellow fruits; zep/zep, orange fruits.
ZEP 유전자에서 탐색된 한 개의 SNP를 이용하여 ZEP1.2-SNP-HRM 분자표지를 개발하였다(Figs. 1 and 2B). Fig. 3B에서 melting 온도가 76.5°C에서 peak가 보이는 것은 정상 대립유전자인 Zep를 가지고 있는 것을 나타내고, melting 온도가 77.1°C에서 peak가 보이는 것은 돌연변이 대립유전자인 zep를 가지고 있는 것을 나타낸다(Fig. 3B). 이형접합체(heterozygote)는 두 peak 사이인 76.8°C에서 peak를 보였다(Fig. 3B). 이후 ZEP 유전자형 분석은 ZEP1.2-SNP-HRM 분자표지를 이용하여 분석하였다(Fig. 3B and Suppl. Fig. S4).
개발된 분자표지의 효용성 검정
앞에서 개발된 CCS-indel-HRM과 ZEP1.2-SNP-HRM 분자표지는 44개의 F1 품종, 31개의 내혼계(inbred lines), 31개의 ‘GINA’ F2 식물체, 79개의 ‘JBR-10’ F2 식물체 및 380개의 유전자원에 적용하여 보았다(Suppl. Figs. S3 and S4).
파프리카 F1 품종 분석에서 빨간색은 Ccs/Ccs 또는 Ccs/ccs 유전자형 그리고 Zep/Zep 또는 Zep/zep 유전자형을 가지고 있었고, 노란색은 ccs/ccs 유전자형과 Zep/Zep 또는 Zep/zep 유전자형을 가지고 있었고 주황색은 ccs/ccs 유전자형과 zep/zep 유전자형을 가지고 있었다(Table 4). 본 결과를 통해 빨간색은 정상 Ccs 대립유전자를 최소한 1개는 가지고 있어야 한다는 것을 알았고, 노란색과 주황색은 정상 Ccs 대립유전자가 없을 때 나타난다는 것을 알 수 있었다(Tables 4 and 5). 노란색은 정상 Ccs 대립유전자가 없을 때 정상 Zep 대립유전자를 최소한 1개를 가지고 있어야 나타난다는 것을 알았고, 주황색은 정상 Ccs 대립유전자와 정상 Zep 대립유전자 둘 모두 없어야 나타난다는 것을 알 수 있었다(Tables 4 and 5).
Table 4.
Fruit colors and genotypes of CCS-indel-HRM and ZEP1.2-SNP-HRM markers in F1 paprika
파프리카 내혼계 분석에서도 빨간색은 정상 Ccs 대립유전자를 모두 가지고 있었고, 노란색은 정상 Ccs 대립유전자가 없었고 정상 Zep 대립유전자를 가지고 있었다(Suppl. Table S2).
파프리카 F1 품종 ‘GINA’의 F2 식물체 분석에서 CCS 및 ZEP 유전자 모두 분리하였는데, Ccs/Ccs 또는 Ccs/ccs 유전자형을 가지고 있는 것은 모두 빨간색이었고, ccs/ccs 유전자형을 가지고 있는 것은 모두 노란색이었다(Suppl. Table S3). 이러한 결과는 본 연구에서 개발된 CCS-indel-HRM 분자표지가 과색 표현형과 연관되어 있다는 것을 보여주는 것이다. 또한 본 분리집단에서 노란색은 ccs/ccs 유전자형과 Zep/Zep 또는 Zep/zep 유전자형을 가지고 있을 때 나타났으나, 주황색은 나타나지 않았는데, 집단 크기가 작아 ccs/ccs, zep/zep 유전자형이 나타나지 않았기 때문이다(Suppl. Table S3). 이러한 결과는 Zep 대립유전자가 zep 대립유전자에 대해 우성으로 작용하는 것을 보여주는 것이다(Table 5).
파프리카 F1 품종 ‘JBR-10’의 F2 식물체 분석에서 CCS 유전자는 분리되고 ZEP 유전자는 정상 Zep 대립유전자로 고정되어 있었다(Suppl. Table S4). 따라서 빨간색과 노란색이 분리되었는데, CCS-indel-HRM 분자표지의 유전자형과 100% 일치하는 결과를 얻었다(Suppl. Table S4).
파프리카 유전자원 분석에서 ccs/ccs 및 zep/zep 유전자형을 가지고 있는 개체는 총 24개였는데 모두 주황색이었다(Suppl. Table S5). 그리고 ccs/ccs 및 Zep/Zep 유전자형을 가지고 있는 개체는 총 45개였는데 모두 노란색이었다(Suppl. Table S5). 나머지 빨간색은 Ccs/Ccs 또는 Ccs/ccs 유전자형을 가지고 있었으며, ZEP 유전자에 대해서는 세 가지 유전자형(Zep/Zep, Zep/zep, zep/zep)이 모두 나타났다(Suppl. Table S5). 이러한 결과는 CCS 유전자가 ZEP 유전자에 대해 상위유전자라는 것을 보여주는 것이다(Table 5).
파프리카 유전자원 분석에서 30–5, 54–6, 54–7, 54–8번의 4개 식물체는 Ccs/Ccs 및 Zep/Zep 유전자형을 가지고 있으면서 노란색 과색 표현형을 보였다(Suppl. Table S5). 이는 앞에서의 결과와 맞지 않는 것으로 CCS 유전자에서 결실이 일어나지 않은 다른 돌연변이일 것이라 생각해 염기서열 분석을 수행하였다. 그 결과 CCS 유전자의 1201번째 염기와 1264번째 염기에서 결실이 일어난 것을 확인하였다(Fig. 2A). 이 돌연변이 유전자를 ccs-m2로 명명하였다(Fig. 2A). 본 연구의 파프리카 유전자원은 농업유전자원센터에서 분양 받은 것인데, 이 4개 식물체는 파프리카로 분양 받았지만 실제로 재배하여 보았을 때, 과형은 파프리카 보다 일반 고추와 더 유사하였다. 따라서 ccs-m2 돌연변이 대립유전자는 일반 고추에서 유래한 것으로 추정된다.
CCS 및 ZEP 유전자형 조합과 파프리카 과색
카로티노이드는 파프리카의 성숙 과색에 영향을 주며, 일부 카로티노이드 생합성 유전자의 돌연변이로 인하여 노란색과 주황색 등 다양한 과색을 나타내게 된다(Gómez-García and Ochoa-Alejo, 2013). 본 연구에서는 빨간색, 노란색, 주황색 성숙 과색에 영향을 주는 카로티노이드 생합성 유전자(CCS와 ZEP)의 변이를 탐색했다(Fig. 2). 고추종의 과색에 관한 선행 연구 결과, PSY1 유전자와 CCS 유전자의 변이와 유전형에 의해 빨간색, 노란색, 주황색의 과색이 결정된다고 보고되었다(Jang et al., 2020). 그러나 본 실험 결과 파프리카에서는 PSY1 유전자의 변이가 탐색되지 않았으며, 미니 파프리카와 동일한 ZEP 유전자에서 SNP가 확인이 되었다(Fig. 2B; Lee et al., 2021). 파프리카의 CCS 유전자의 변이는 2019년 보고된 CCS 유전자의 변이와 유사한 결실 부위를 확인을 하였다(Fig. 2A; Jeong et al., 2019).
CCS 유전자는 ZEP 유전자에 대하여 상위성을 가지는 것을 확인하였다(Table 5 and Suppl. Table S5). CCS 유전자가 Ccs/Ccs 또는 Ccs/ccs 유전자형일 때, ZEP의 유전자형(Zep/Zep, Zep/zep, zep/zep)에 관계없이 빨간색의 과색이 나타났으며, CCS 유전자가 ccs/ccs 유전형일 때, ZEP 유전자가 Zep/Zep 또는 Zep/zep 유전자형일 때는 노란색의 과색을 나타내고, zep/zep 유전자형일 때는 주황색의 과색을 나타내는 것으로 확인되었다(Table 5 and Suppl. Table S5).
Table 5.
Relationship between CCS/ZEP genotypes and fruit colors in paprika
| CCS z genotype | ZEP y genotype | Fruit color of paprika |
| Ccs/Ccs or Ccs/ccs | Zep/Zep or Zep/zep | Red |
| zep/zep | Red | |
| ccs/ccs | Zep/Zep or Zep/zep | Yellow |
| zep/zep | Orange |
본 연구에서 개발한 분자표지는 새로운 빨간색, 노랑색, 주황색 파프리카 품종을 육성할 때 유용하게 사용될 수 있을 것으로 생각되며, 서로 다른 과색의 파프리카를 교배하여 육종할 수 있어 파프리카 품종 다양성을 확대하는데 기여할 수 있을 것이다.
Supplementary Material
Supplementary materials are available at Horticultural Science and Technology website (https://www.hst-j.org).
- HORT_20240052_Fig_S1.pdf
Supplementary Fig. S1. Three fruit colors, red (A), yellow (B), and orange (C), of paprika.
- HORT_20240052_Fig_S2.pdf
Supplementary Fig. S2. Melting curves of HRM analysis for genotyping of CCS gene. (A) HRM analysis with three primers including CCS-com1-R, CCS-ins1-F and CCS-del3-F; (B) HRM analysis with three primers including CCS-com1-R, CCS-ins3-F and CCS-del3-F; (C) HRM analysis with three primers including CCS-com2-R, CCS-ins1-F and CCS-del3-F.
- HORT_20240052_Fig_S3.pdf
Supplementary Fig. S3. Melting curves of CCS-indel-HRM markers in various paprika materials. (A) paprika inbred lines; (B) and (C) are two F2 populations; (D), (E), (F), and (G) are paprika germplasms; Ccs, functional allele of capsanthin-capsorubin synthase gene; ccs, non-functional deleted allele of CCS gene.
- HORT_20240052_Fig_S4.pdf
Supplementary Fig. S4. Melting curves of ZEP1.2-SNP-HRM markers in various paprika materials. (A) paprika inbred lines; (B) and (C) are two F2 populations; (D), (E), (F), and (G) are paprika germplasms; Zep, functional allele of zeaxanthin epoxidase gene; zep, non-functional allele of ZEP gene.
- HORT_20240052_Table_S1.pdf
Supplementary Table S1. List of primer sets for DNA sequencing
- HORT_20240052_Table_S2.pdf
Supplementary Table S2. Fruit colors and genotypes of CCS-indel-HRM and ZEP1.2-SNP-HRM markers in inbred lines of paprika
- HORT_20240052_Table_S3.pdf
Supplementary Table S3. Fruit colors and genotypes of CCS-indel-HRM and ZEP1.2-SNP-HRM markers in F2 plants of the F1 cultivar ‘GINA’
- HORT_20240052_Table_S4.pdf
Supplementary Table S4. Fruit colors and genotypes of CCS-indel-HRM and ZEP1.2-SNP-HRM markers in F2 plants of the F1 cultivar ‘JBR-10’
- HORT_20240052_Table_S5.pdf
Supplementary Table S5. Fruit colors and genotypes of CCS-indel-HRM and ZEP1.2-SNP-HRM markers in paprika germplasms
