

Introduction
Materials and Methods
Plant Materials and Preparation of Explants
Culture Medium
Effect of Plant Growth Regulators on Direct Shoot Induction and Growth
Effect of Sucrose Concentration on Shoot Proliferation and Root Formation
Transplantation
Data Analysis
Results and Discussion
Effect of Explant Types and Growth Regulators on Plant Regeneration
Effect of Liquid Culture and BA Concentration on Shoot Multiplication
Effect of Sucrose Concentration on Bulblet Formation
Transplantation of in Vitro Propagated Plantlets
Conclusions
Introduction
Allium hookeri L. (Alliaceae family) is an important ethno-medicinal plant native to the Himalayan region of Asia (Ayam, 2011; Pandey et al., 2008). This plant has been utilized in the eastern Himalayan region of India as a vegetable and spice, and as a traditional medicine for cough, cold, painful swellings, and skin eruptions (Kala, 2005). However, preservation and cultivation of native A. hookeri germplasm has decreased in part due to the lack of fundamental research on this plant. After the recognition of the potent medicinal and nutritional value of indigenous A. hookeri, it began to be widely cultivated in fields and kitchen gardens in the mountainous regions of India and Myanmar (Ayam, 2011; Pandey et al., 2008). Since this plant was introduced in 2010, its cultivation has expanded very rapidly in many parts of South Korea.
A. hookeri has recently gained significant attention because it produces many useful compounds, such as high amounts of phenolic antioxidants, phytosterols, fiber, ascorbic acid, flavonoids, and allicin, as well as unknown organic sulfur compounds distinct from those of A. cepa (Ayam, 2011; Bae and Bae, 2012; Kala, 2005; Kim et al., 2012; Pandey et al., 2008; Rhyu and Won et al., 2013). Allicins are sulfur- containing compounds which have been demonstrated to have potent pharmacological properties, including anti-microbial activity, the ability to lower cholesterol levels, antiplatelet aggregation effects, the ability to lower the risk of heart attacks, anti-carcinogenic activity, and anti- inflammatory effects (Ayam, 2011; Bae and Bae, 2012; Kim et al., 2012; Rahman and Lowe, 2006; Rhyu and Park, 2013). However, the chemical structure of many novel compounds in A. hookeri and their biological effects on health remain to be elucidated.
Like other Allium species, A. hookeri produces a bulbous stem base, but with a relatively small underground rhizome with fibrous roots (Pandey et al., 2008). Because cultivated forms of A. hookeri are seed-sterile (Seidemann, 2005; Sharma et al., 2011), the propagation of this species has been conducted by conventional vegetative reproduction, which is commonly accomplished by splitting up the branched rhizomes (Lee et al., 2009). However, this method is inefficient for the provision of plantlet stocks for large-scale field cultivation due to the intensive labor requirements, high cost, time consumption, and the risk of viral transmission that could lead to lowered yields (Ma et al., 1994; Novák, 1990). Thus, although there is growing interest in A. hookeri in agriculture and the food industry, it will be difficult to coordinate the balance between demand and supply.
Efficient and rapid technology for in vitro plant propagation may provide an alternative method for the mass propagation of seed-sterile plants. Moreover, this technology is essential to select, expand, and improve valuable traits of plants. Thus, a wide range of methods for micropropagation via direct organogenesis, callus culture, and somatic embryogenesis have been developed to grow and improve the sterile Allium species over the last decade (Gantait et al., 2010). Plant regeneration of garlic (A. sativum L.), leek (A. porrum L. and A. ampeloprasum L.), and onion (A. cepa L.) has been successfully demonstrated using cultured callus (Abo El-Nil, 1977; Myers and Simon, 1999), shoot tips (Nagakubo et al., 1993), leaves (Rauber and Grunewaldt, 1988), inflo-rescences (Dunstan and Short, 1977; Matsubara and Hihara, 1978; Novák and Havel, 1981; Silvertand et al., 1995), and roots (Haque et al., 1997; Kim et al., 2003; Robledo-Paz et al., 2000). Based on those studies, the most critical factors determining plant regeneration are the type of explants and combinations of plant growth regulators used in the culture medium. It has been reported that both cytokinins and auxins synergistically affect in vitro plant regeneration of some Allium species, including A. sativum (Bhojwanii, 1980; Haque et al., 1997), A. ampeloprasum (Silvertand, 1995), A. ascalonicum (Mohamed-Yasseen et al., 1994), and A. wallichii (Wawrosch et al., 2001). Liquid or solid MS (Murashige and Skoog, 1962), Gamborg B5 (B5) (Gamborg et al., 1968), and LS (Linsmaier and Skoog, 1965) media have been widely used as basal media in the tissue culture of Allium species. However, despite the potential value of A. hookeri as a food crop and source of important medicines, Jha and Jha (1989) had only reported the in vitro plant regeneration of A. hookeri using basal segments of leaves. However, their research mainly focused on cytological studies of a group of A. hookeri Thw. rather than optimization of plant regeneration system in detail.
In this study, a highly efficient in vitro propagation system for A. hookeri was developed by in vitro culturing its flower bud receptacle for 9 weeks, with a maximum yield of 62 plantlets per receptacle.
Materials and Methods
Plant Materials and Preparation of Explants
Freshly grown Allium hookeri L. plants were collected from local farms located in the Jeonbuk region of South Korea during the period from July 15 to August 30 in 2013. The plant was separated into immature flower buds, roots, and leaves and treated with 70% (v/v) ethanol for 5 min. After washing with distilled water, tissues were surface sterilized in a 2% (v/v) sodium hypochlorite solution containing 0.01% (v/v) Tween-20 for 30 min and rinsed three times in sterilized distilled water. The tissues were used as explants by cutting them into small segments under aseptic conditions. The length of the root, flower stalk, and leaf explants was approximately 3 to 5 mm. Receptacle explants were prepared by removing involucre spathes and bulbils from immature flower buds.
Culture Medium
The culture medium used for in vitro regeneration of A. hookeri was Gamborg B5 (B5) (Gamborg et al., 1968) basal medium containing micro- and macro-elements with vitamins. The pH of all media was adjusted to 5.7 with 1 N KOH prior to autoclaving. The media was solidified by adding 2.5 g・L-1 Gerlite (Duchefa Biochemie, The Netherlands). Aliquots of the medium were dispensed into plastic petri dishes or glass culture bottles. Plant Preservative Mixture (PPM; Plant Cell Technology, Inc., USA) was included in all media at a final concentration of 0.1% (v/v) as an antimicrobial agent.
Effect of Plant Growth Regulators on Direct Shoot Induction and Growth
To examine the effects of 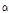 -naphthalene acetic acid (NAA; Duchefa Biochemie, The Netherlands) and 6-benzylaminopurine (BA; Duchefa Biochemie, The Netherlands) on shoot formation, each explant was incubated on B5 media containing 30 g・L-1 sucrose, 2.5 g・L-1 Gelrite and supplemented with different combinations and concentrations of BA and NAA at 24°C for 3 weeks under 2,500 lux light with a 16 h photoperiod in a growth chamber. The combinations of growth regulators included 0.25 μM BA + 2.5 μM NAA, 0.5 μM BA + 5 μM NAA, 1 μM BA + 10 μM NAA, 2.5 μM BA + 0.25 μM NAA, 5 μM BA + 0.5 μM NAA, and 10 μM BA + 1 μM NAA. Each experiment was carried out with 3 replicates of 20 explants. The percentage of shoot, root, and callus induction from the explants was recorded after 3 weeks of culture. The explants with shoots grown on the media containing 5 μM BA + 0.5 μM NAA or 10 μM BA + 1 μM NAA were then divided into 2 to 4 segments and transferred to solid or liquid B5 medium containing BA alone (0, 2.5, 5, or 10 μM) or 10 μM BA in combination with 1 μM NAA. After 3 weeks of subculture on a rotary shaker at 90 rpm, the multiplication of shoots per explant was measured. Each experiment was carried out with 3 replicates of 6 explants.
-naphthalene acetic acid (NAA; Duchefa Biochemie, The Netherlands) and 6-benzylaminopurine (BA; Duchefa Biochemie, The Netherlands) on shoot formation, each explant was incubated on B5 media containing 30 g・L-1 sucrose, 2.5 g・L-1 Gelrite and supplemented with different combinations and concentrations of BA and NAA at 24°C for 3 weeks under 2,500 lux light with a 16 h photoperiod in a growth chamber. The combinations of growth regulators included 0.25 μM BA + 2.5 μM NAA, 0.5 μM BA + 5 μM NAA, 1 μM BA + 10 μM NAA, 2.5 μM BA + 0.25 μM NAA, 5 μM BA + 0.5 μM NAA, and 10 μM BA + 1 μM NAA. Each experiment was carried out with 3 replicates of 20 explants. The percentage of shoot, root, and callus induction from the explants was recorded after 3 weeks of culture. The explants with shoots grown on the media containing 5 μM BA + 0.5 μM NAA or 10 μM BA + 1 μM NAA were then divided into 2 to 4 segments and transferred to solid or liquid B5 medium containing BA alone (0, 2.5, 5, or 10 μM) or 10 μM BA in combination with 1 μM NAA. After 3 weeks of subculture on a rotary shaker at 90 rpm, the multiplication of shoots per explant was measured. Each experiment was carried out with 3 replicates of 6 explants.
Effect of Sucrose Concentration on Shoot Proliferation and Root Formation
To examine the effect of sucrose concentration on shoot growth and root formation, explants with vigorous multiple shoots were divided into clumps of 2 to 5 shoots and incubated in B5 liquid medium containing 10, 30, 70, or 100 g・L-1 of sucrose without growth regulators for 3 weeks on a rotary shaker at 90 rpm. The fresh weight of the bulblet was determined by subtracting the initial value from the final value after 3 weeks. Each experiment was carried out with 3 replicates of 10 shoot clumps per culture bottle.
Transplantation
One hundred regenerated plantlets were removed from their culture bottles and washed in running tap water before transferring them to pots containing garden soil (Bioplug #1, Hungnong Seeds Co., Ltd., South Korea). Each pot was covered with plastic wrap to ensure high humidity for the first 3 days. After this adaptation period, the plants were transferred to greenhouse conditions. The percentage of survival was evaluated 2 weeks after transplantation.
Data Analysis
Data were statistically analyzed as a completely randomized design with 3 replicates. Analysis of paired sample t-test and variance (ANOVA) followed by Tukey’s multiple comparison test at 0.05 probability level was performed to compare the means between each experiment using OriginPro software version 8.5 (OriginLab Co., USA).
Results and Discussion
Effect of Explant Types and Growth Regulators on Plant Regeneration
A. hookeri has long been used as a vegetable and traditional medicine in Southeast Asia (Ayam, 2011; Pandey et al., 2008). Recently, the cultivation of A. hookeri has been rapidly growing due to its considerable value as a medicinal and edible crop. Since A. hookeri is a seed-sterile plant, multiplication of this plant has been accomplished primarily through division of underground rhizomes, which is called vegetative propagation. Thus, in vitro propagation through plant regeneration might offer an alternative approach for generating plant stocks for large-scale agriculture. However, there have been very little studies on the in vitro propagation of this species so far. The aim of this study is to develop an efficient in vitro propagation system of A. hookeri for producing high-quality plants in a relatively short period of time.
In general, the most important factors affecting plant regeneration systems are plant genotype, explant type, plant growth regulators (PGRs), and composition of medium (Gantait et al., 2010). As an initial step for establishing an in vitro plant regeneration system for A. hookeri, the frequency of shoot, root, and callus induction in various explant types was examined on B5 basal medium supple-mented with 30 g・L-1 sucrose and different concentrations and combinations of BA and NAA. After 3 weeks of incubation at 24°C under light, a high frequency of shoot, root, and callus formation was observed from the cultured flower bud receptacles (Table 1 and Fig. 1). However, the other explants, including the flower stalks, bulbils, roots and leaves, enlarged slightly, but did not show significant responses under any conditions during the incubation period (data not shown). This suggests that the receptacle is the most favorable explant source for the regeneration of A. hookeri.

Direct shoot regeneration began when receptacle explants became swollen after culturing for 10 days. Development of multiple shoots continued thereafter throughout the incubation period. The maximum percentage of shoot- generating receptacle explants was 93.33 ± 4.63%, when cultured on B5 medium supplemented with 10 μM BA and 1 μM NAA (Table 1 and Fig. 1G). This frequency was higher than those of other Allium species. In the case of A. sativum, the maximum frequency of shoot regeneration from root explants (Haque et al., 1997) or shoot tips (Nagakubo et al., 1993) was 75% or 41%, respectively, when cultured on MS medium (Murashige and Skoog, 1962) with a combination of 1 μM NAA and 10 μM BA, or modified LS medium (Linsmaier and Skoog, 1965) with a combination of 5 μM indole-3-acetic acid (IAA) and 10 μM BA, respectively. Silvertand et al. (1995) reported that the maximum fre-quencies of shoot induction from flower stalk explants of several A. ampeloprasum cultivars were from 84 to 100% when cultured on MS medium with a combination of 10 mg・L-1 BA and 10 mg・L-1 NAA. Like shoot induction, the highest number of shoots (18.72 ± 7.47) per explant was also observed on B5 medium containing 10 μM BA and 1 μM NAA, whereas B5 medium containing no growth regulators or relatively higher concentrations of NAA in combination with BA gave significantly (p < 0.05) lower numbers of shoots per explant (Table 2). In addition, direct root formation was induced when receptacle explants were cultured on B5 medium supplemented with 0.5 μM BA and 5 μM NAA, at a frequency of 76.67 ± 7.85% (Table 1 and Fig. 1C). Under these conditions, some explants (25.0 ± 7.86%) simultaneously produced both roots and shoots on the surface of explants. However, the number of roots per explant showed no significant difference (p > 0.05) between cultures, ranging from 2.17 ± 0.75 to 2.98 ± 1.55 roots per explant (Table 2). These results are in agreement with earlier studies showing that a lower auxin and higher cytokinin level promoted multiple shoot induction, whereas a lower cytokinin and higher auxin level resulted in enhance-ment of root formation in A. sativum (Bhojwanii, 1980; Haque et al., 1997), A. ampeloprasum (Silvertand, 1995), A. ascalonicum (Mohamed-Yasseen et al., 1994), and A. wallichii (Wawrosch et al., 2001).
Instead of direct plant regeneration, callus formation was induced on the surface of receptacle explants at a maximum frequency of 80.0 ± 7.43% when cultured on B5 medium supplemented with 1 μM BA and 10 μM NAA after 3 weeks of culture (Table 1 and Fig. 1D).
Effect of Liquid Culture and BA Concentration on Shoot Multiplication
It is known that liquid culture, with or without cytokines, is generally more efficient than solid support systems for shoot proliferation and multiplication for several species of plants, including Allium species (Escalona et al., 1999; Ilan et al., 1995; Kim et al., 2003). To examine the effect of culture type and BA concentration on shoot multiplication, explants with shoots were divided into 2 to 4 segments and further subcultured in solid or liquid B5 medium supplemented with different concentrations of BA (0, 2.5, 5, or 10 μM) alone, or 10 μM BA in combination with 1 μM NAA. After further subculturing for 3 weeks under light, a highly variable rate of shoot multiplication per explant was obtained (Table 3). Regardless of the BA concentration in the medium, the increase in shoot number per explant was significantly higher in liquid medium than in solid medium, when the mean total scores of the two culture types were compared by paired sample t-test (33.33 ± 10.75 vs. 14.93 ± 5.50; t = 6.60998, 4 d.f., p = 0.00272), suggesting that liquid medium is more promising than solid medium for shoot multiplication (Table 3, Figs. 2A, and 2B). Additionally, higher concentrations of BA, ranging from 5 to 10 μM, significantly enhanced shoot multiplication in both liquid and solid medium compared to BA-free medium. However, the addition of 1 μM NAA to the media did not significantly (p = 0.99813) influence the rate of shoot multiplication, suggesting that BA alone is sufficient for shoot multiplication. This result agrees with previous reports that initiation of multiple shoots is successfully promoted by a single cytokinin, such as 5 μM BA for A. cepa (Kahane et al., 1992), 0.5 mg・L-1 BA for A. tuberosum (Pandy et al., 1992), 4.4 μM BA for A. ampeloprasum (Mohamed-Yasseen et al., 1995), and 0.5 μM BA for A. sativum (Haque et al., 1998). In summary, liquid B5 medium containing 5 to 10 μM BA is suitable to induce a high rate of shoot multiplication from receptacles in A. hookeri.
Effect of Sucrose Concentration on Bulblet Formation
It has been demonstrated that high sucrose concentrations, ranging from 6-12% (w/v), promote in vitro bulblet formation and growth in many Allium species, including A. sativum (Haque et al., 1997; Kim et al., 2003; Nagakubu, 1993), A. cepa (Dunstan and Short, 1977; Matsubara and Hihara, 1978), A. porrum (Novák and Havel, 1981) and A. ampeloprasum (Silvertand et al., 1995). To estimate the optimum sucrose concentration for bulblet formation and growth, elongated shoot clusters (2-5 shoots) greater than 2 cm in length were dissected from proliferated shoot clumps and subcultured in B5 liquid medium containing 10, 30, 70, or 100 g・L-1 sucrose without PGR (Figs. 2C to 2D). After 3 weeks of subculture, over 95% of the shoots had a bulbous stem base and gave rise to adventitious roots in all of the media. This suggests that PGR-free medium is sufficient to induce bulblet formation and successful rhizogenesis, as observed in A. sativum (Haque et al., 1997; Ma et al., 1994). Otherwise, the sucrose concentration significantly (p < 0.05) affected the fresh weight and growth of bulblet. Shoot clumps growing in B5 medium with 1 to 7% (w/v) sucrose showed green-colored normal growth, whereas those growing in a medium with 10% (w/v) sucrose were light green, with a tendency to vitrification. Maximum fresh weight per bulblet was observed when cultured in B5 liquid medium with 7% (w/v) sucrose.

Transplantation of in Vitro Propagated Plantlets
After bulblet and root formation in liquid B5 medium containing 7% (w/v) sucrose without PGR, propagated plantlets were washed with tap water and transferred to garden soil. Some heavily clumped plantlets were separated from other plantlets for transplanting. Most of the trans-planted plantlets (greater than 95%) began to grow 5 to 7 days after ex vitro transplantation (Fig. 2E). These successfully developed into mature plants with no morphological abnor-malities under greenhouse conditions.
In this study, a new high-frequency regeneration system for mass clonal propagation of A. hookeri plant was established using receptacle explants. The results demonstrated that flower bud receptacles are the best explant for direct root and shoot induction or callus formation on B5 medium with particular concentrations and combinations of BA and NAA within 3 weeks. Moreover, a maximum of 62 shoots per explant was obtained through one round of liquid subculture with 10 μM BA for an additional 3 weeks. When small numbers of shoots dissected from the shoot clumps of the explants were cultured in B5 liquid medium containing different concentrations of sucrose for 3 weeks, most shoots formed bulblets with adventitious roots, with the highest rate observed in 7% (w/v) sucrose. Moreover, the survival rate of micropropagated plants was over 95% when trans-planted into potted soil, with no morphological abnormalities. Thus, a high-frequency in vitro regeneration system of A. hookeri established in this study might be beneficial for the mass production of high quality clones and further optimization of in vitro culture conditions.
