

서 언
토마토는 건강기능식품으로 인정받으면서 대중 수요 증가로 인해 2000년 4,746ha에서 2020년 5,521ha로 연평균 재배면적이 조금씩 증가하고 있다(Lee et al., 2019). 한편, 2003년 학교급식의 전면 실시는 토마토의 소비증가 요인이 되어 연중 안정된 생산이 요구되고 있어, 이를 위한 재배 및 환경관리 기술의 지원이 필요한 실정이다(RDA, 2018). 이러한 상황에 이상기온에 의한 여름철 고온이 작물 생산성을 감소시키고 연중 안정공급에 영향을 미치므로 고온에 대응하는 재배기술의 필요성이 대두되고 있다(Parry et al., 2004).
육묘의 경우 온실 내로 유입된 복사열 때문에 외부보다 온실 내부의 온도가 높아져 적정 온도를 유지하는데 어려움이 있어 환기, 차광, 증발냉각 및 지붕 살수를 이용하여 고온기를 극복하고 있으나 적정온도 유지에는 한계가 있다(Lee and Kim, 2011). 또한 소규모 온실을 운영하는 농가에서는 과도한 시설투자를 요구하는 냉난방 시스템을 갖추는 데 어려움이 있다(Choi et al., 2014; Kim et al., 2022).
토마토 육묘는 대부분 전문 육묘장에서 이루어지고 있으며, 일부 대형 육묘장은 시설투자를 통해 환경개선이 이루어지고 있으나 많은 중소 육묘장에서는 외부 환경에 영향을 받아 고온기 및 저온기에 건전한 묘를 생산하기에 어려운 상황이다(An et al., 2021). 연중 고품질 묘를 생산하기 위해 폐쇄형 육묘시스템이 개발되어 단기간 균일한 묘를 생산할 수 있게 되었으나(Kim et al., 2005; Kozai et al., 2000), 높은 시설 투자비용으로 인해 농가에 적용하기 어려움이 있을 것으로 생각된다. 특히 혹서기에 육묘장에서 작물 재배의 적정온도를 벗어날 경우 생리장해를 유발할 수 있다(Nam et al., 2015). 혹서기에 육묘장의 적정온도를 유지하기 위해 온실내부에 냉방설비를 할 경우 냉방부하가 매우 크며 이를 위한 초기비용과 에너지 소모량에 따른 운영비용이 높아 작물 생육에 적합한 온실 환경을 유지하는데 어려움이 많다(Nam et al., 2015: Choi et al., 2015). 이와 같은 문제를 해결하기 위해 진행되었던 선행연구 결과에 따르면 혹서기나 혹한기에 근권부 온도를 생육적온인 20–25°C로 유지하여 온도 스트레스에 의한 피해를 경감시킬 수 있다고 하였다(Chaisompongpan et al., 1990; Kwack et al., 2014). 고온으로 인한 대기온도가 40°C 이상 상승하는 온실에서 근권부 온도가 33°C 이상 상승하기 때문에 묘품질의 저하 없이 묘를 생산하는 것은 불가능하다(Kim et al., 2022). 이러한 문제 해결을 위해 양액온도 조절을 통하여 작물의 근권부 온도를 제어한다면 온실 전체를 냉방하는 것보다 냉난방에 소요되는 에너지 비용을 줄일 수 있을 뿐 아니라 작물의 양수분 흡수 저해를 완화시키고, 뿌리의 생장을 촉진시켜 생육 및 수량에 긍정적인 영향을 끼칠 수 있다(Moon et al., 2007; Morgan et al., 2011). 이러한 국부적 근권부 온도 조절 시스템을 이용하여 외부 환경에 대응한 최적 근권부 환경을 구명한다면 저온기, 고온기 및 이상기후에도 효율적인 대응이 가능해질 것이다.
따라서 본 연구는 고온기 및 저온기의 효율적인 대응을 위하여 근권부 온도 제어 방식으로서 공급 양액의 온도조절을 통해 토마토의 초기 생육반응을 관찰하고 지상부 온도에 대응한 근권부 온도 제어 범위를 구명하기 위하여 실험을 수행하였다.
재료 및 방법
지상부 및 지하부 온도 처리
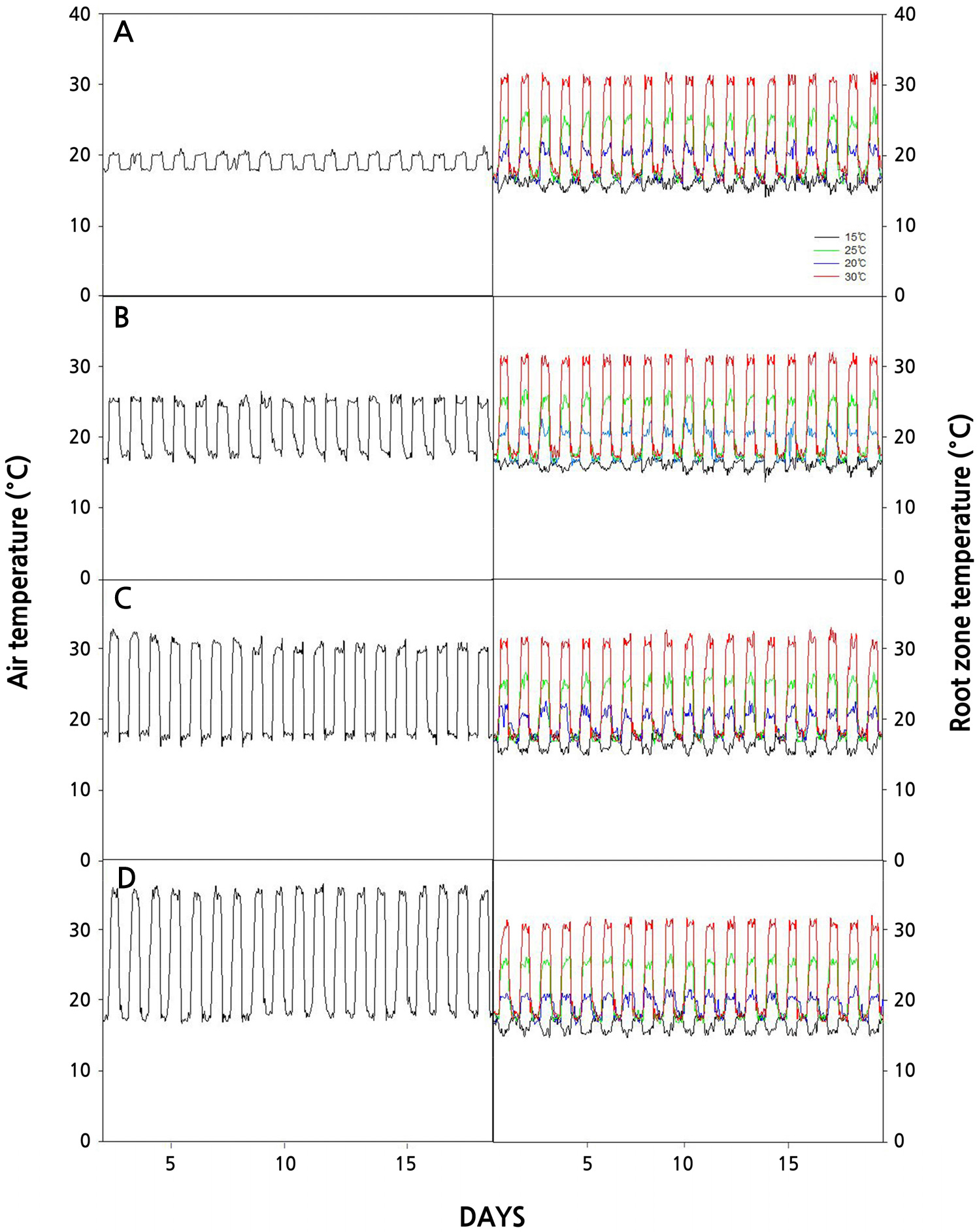
본 실험은 2022년 6월 5일부터 10월 11일까지 목포대학교 부속농장에 위치한 기후 조절용 23m2 규모의 소형 유리온실(7'3'2.3m)에서 진행하였다(Fig. 1). 온실의 대기온도는 공조기(DUS-100E, DONGHWA Co. Ltd, Korea)를 이용하여 주간 대기온도 20, 25, 30 및 35°C로 각각 조절하였고, 모든 대기온도 처리구의 야간 대기온도는 18°C로 조절하였다. 근권부 온도는 각각의 대기온도 처리구에서 주간 12시간 동안 15 ± 3, 20 ± 3, 25 ± 3 및 30 ± 3°C로 설정하여 실험을 수행하였다(Fig. 3). 따라서 야간의 근권부 온도는 별도로 조절하지 않아 18 ± 1°C로 유지되었다. 온실의 광량은 SMPS(Switched Mode Power Supply)형 LED램프(T5-15W-6500K-1500lm, NAMYUNG Lighting Co. Ltd., Korea)를 이용하여 원형포트 상단의 위치에서 200 ± 20µmol·m-2·s-1로 설정하였고, 명 12시간, 암 12시간으로 조절하였다. 또한 온실의 외부를 차광률 99%필름으로 차폐하여 외부 광량의 영향을 받지않도록 하였다(Fig. 1A). 근권부 온도조절은 양액탱크(200L)에 냉각기(DA-500B, DAEIL Co. Ltd., Korea), 전기온수기(KD-PD-S13K, KDheater Co. Ltd., Korea) 및 온도조절기(RMD-100-220-30A, Run electronics Co. Ltd., Korea)를 설치하여 양액의 온도를 조절하였다(Fig. 2). 온도가 조절된 양액은 펌프(PW350sm, Willo, Korea)와 점적드리퍼(2L/H Dripper set, Netafim, Israel)를 이용하여 배지에 공급해 근권부의 온도를 각각 15 ± 3, 20 ± 3, 25 ± 3 및 30 ± 3°C로 조절하였으며(Figs. 1B and 2), 조절된 대기온도와 근권부 온도는 측정기기(WatchDog 1000series Micro stations, Spectrum Technologies. Inc, USA)를 통해 모니터링 하였다(Fig. 3).


식물 재배 및 생장 분석
토마토(Solanum lycopersicum L., Dafnis, Syngenta Korea, Korea) 종자를 암면 셀트레이(Plantop Plug, Grodan, Netherlands)에 파종한 후 양액(EC 1.5, pH 6)으로 침지시켜 생장상(OS-SB2000, Onsol Co. Ltd., Korea)에서 7일 동안 발아시켰다. 생장상의 온도는 30°C, 광량 100 ± 10µmol·m-2·s-1 로 설정하였다. 파종 7일 후 떡잎 2매가 전개된 개체 중 균일한 묘를 선발하였고, 이를 0.5L의 상토(Eoullim, Shinsung Mineral Co., Korea)가 충진된 원형 플라스틱 포트(Ø100mm × H90mm)에 이식한 후 실험 온실로 옮겨 지상부와 근권부 온도 처리를 시작하였다. 근권부 온도 15 ± 3, 20 ± 3, 25 ± 3 및 30 ± 3°C로 설정된 모든 대기온도 실험은 다음과 같이 기간을 달리하여 실험을 수행하였다. 대기온도 20°C 처리구는 6월 5일부터 7월 3일까지 진행하였으며, 대기온도 25°C 처리구는 7월 6일부터 8월 2일, 대기온도 30°C처리구는 8월 9일부터 9월 6일, 대기온도 35°C처리구는 9월 12일부터 10월 11일까지 진행하였다.
근권부 온도 조절용 양액은 EC 2.5dS·m-1, pH 6으로 조정된 PBG표준양액(Table 1)을 사용하였으며, 주간(12시간)동안 6분 간격으로 1분동안 33mL씩 점적관주 하였고, 야간(12시간)에는 공급을 중단하였다. 생육 조사는 각각의 처리구당 5개체를 임의로 선발하여 조사하였고 파종 후 15일, 22일, 29일차에 파괴조사를 하였으며, 초장, 근장, 엽수, 엽면적, 지상부와 지하부의 생체중 및 건물중 측정하였다. 건물중은 생체중 측정 후 70°C로 설정된 건조기(JSOF-250T, JSR, Korea)에 168시간 동안 건조한 후 측정하였다. 엽면적은 엽면적 측정기(LI-3100 AREA METER, LI-COR Inc., USA)를 이용하여 측정하였다.
Table 1.
Nutrient solution for tomatoes (PBG nutrient solution from the Netherlands)
| Component | Macro elements (mM) | Micro elements (mM) | ||||||||||||
| NH4 | K | Ca | Mg | NO3 | SO4 | H2PO4 | B | Fe | Mn | Zn | Cu | Mo | ||
| 1.0 | 7.5 | 4.3 | 1.5 | 15.3 | 1.8 | 1.3 | 30.0 | 15.0 | 10.0 | 5.0 | 0.75 | 0.5 | ||
파종후 29일차 생육조사를 바탕으로 묘소질의 우수성을 확인하기 위해 농촌진흥청의 농업과학기술 연구조사 분석기준(RDA, 2012)과 기존 연구(Jeong et al., 2020)를 기초로 상대생장율(RGR, relative growth rate), 작물생장율(CGR, crop growth rate), T/R율(T/R ratio) 및 충실도(Compactness) 등을 아래와 같은 계산식으로 산출하여 비교하였다.
상대생장률(RGR, g-1·g-1·d-1) = (logew2–logew1)/(t2–t1)
작물생장율(CGR, g-1·m-2·d-1) = (w2–w1)/(t2–t1)
충실도(compactness, mg·cm-1) = 지상부의 건물중/식물체의 초장
T/R율(T/R ratio) = 지상부의 건물중/지하부의 건물중
․ w1, w2: 시작 및 종료시점의 식물체당 건물중
․ t1–, t2: 시작 및 종료시점의 시간
통계 분석
실험구 배치는 난괴법으로 처리구당 실험구 3반복으로 수행하였다. 지상부 및 지하부 온도에 따른 토마토의 초장, 근장, 엽수, 엽면적, 생체중, 건물중, CGR, 충실도, T/R율 및 RGR은 SPSS 통계 프로그램(IBM SPSS Statistics 26, IBM CO., Armonk, NY, USA)을 이용하여 분석하였으며, 2요인 분산 분석과 Duncan의 다중 검정(p < 0.05)으로 평균간의 차이에 대한 유의성을 검정하였다. 그래프는 SigmaPlot프로그램(Sigma Plot 12.0, Systat Software Inc., USA)을 이용하여 작성하였다.
결과 및 고찰
대기온도 20, 25, 30 및 35°C에서 근권부 온도를 각각 15, 20, 25 및 30°C로 조절하여 토마토 초장을 조사한 결과 대기온도 30, 35°C의 모든 근권부 온도 처리구에서 토마토의 초장은 다른 처리구에 비해 길었으며, 특히 대기온도 30°C에 근권부 온도를 25와 30°C로 조절한 조합처리구와 대기온도 35°C에 근권부 온도를 20, 25°C로 조절한 조합처리구의 초장이 가장 길었다(Tables 2, 3, 4 and Fig. 4). 월동무와 백침계 오이의 경우 대기 온도가 높을수록 초장이 증가한 것으로 보고된 바 있으며(Oh et al., 2017; Park et al., 2004), 주야간 온도차(DIF)가 높을수록 내성 호르몬인 GA의 생산이 많아져 초장이 길어진다는 연구결과가 보고된 바 있다(Son, 1995). 본 연구결과에서도 주야간 온도차가 높을수록 토마토 초장이 증가하여 선행연구 결과와 일치하는 경향을 보였다.
Table 2.
Growth characteristics of tomato seedlings as affected by different air temperatures and root zone temperatures 15 days after the sowing (n = 5, R = 3). Different letters indicate significant differences according to Duncan’s multiple range test at p < 0.05
|
Air temp. (°C) |
Root zone temp. (°C) |
Plant height (cm/plant) |
Root length (cm) |
No. of leaves (ea/plant) |
Leaf area (cm2/plant) |
Fresh weight (g/plant) |
Dry weight (g/plant) | |||
| Shoot | Root | Shoot | Root | |||||||
| 20 | 15 | 4.2 f-h | 3.5 b-e | 2.0 c-e | 1.3 f | 0.18 f-h | 0.102 a | 0.011 fg | 0.002 ab | |
| 20 | 3.8 gh | 4.7 a-c | 2.0 c-e | 1.8 ef | 0.18 f-h | 0.032 b | 0.010 fg | 0.003 ab | ||
| 25 | 4.5 e-h | 3.9 b-e | 2.0 c-e | 1.6 f | 0.22 d-h | 0.034 b | 0.011 fg | 0.006 a | ||
| 30 | 3.5 h | 3.3 c-e | 2.0 c-e | 0.2 f | 0.12 gh | 0.016 b | 0.008 g | 0.005 ab | ||
| 25 | 15 | 6.4 b-d | 2.6 de | 2.0 c-e | 2.3 ef | 0.22 d-h | 0.012 b | 0.015 d-h | 0.001 b | |
| 20 | 5.1 d-g | 2.3 e | 2.0 c-e | 3.0 d-f | 0.27 e-g | 0.025 b | 0.019 c-f | 0.001 b | ||
| 25 | 5.6 e-h | 2.4 e | 1.6 de | 2.3 ef | 0.18 f-h | 0.019 b | 0.012 e-g | 0.001 b | ||
| 30 | 5.6 c-f | 2.3 e | 1.4 e | 2.5 ef | 0.17 f-h | 0.017 b | 0.011 fg | 0.001 b | ||
| 30 | 15 | 6.8 bc | 3.1 c-e | 1.8 c-e | 4.5 c-e | 0.31 d-f | 0.029 b | 0.017 d-h | 0.003 ab | |
| 20 | 7.7 b | 3.9 b-e | 2.0 c-e | 5.5 b-d | 0.38 de | 0.045 ab | 0.021 c-e | 0.003 ab | ||
| 25 | 9.9 a | 4.0 b-e | 2.6 c | 8.7 a | 0.74 a | 0.062 ab | 0.038 a | 0.003 ab | ||
| 30 | 9.6 a | 4.3 a-d | 2.4 cd | 6.4 a-c | 0.46 cd | 0.034 b | 0.022 cd | 0.002 ab | ||
| 35 | 15 | 6.8 bc | 4.1 a-e | 5.2 a | 7.4 ab | 0.69 ab | 0.034 b | 0.033 ab | 0.002 ab | |
| 20 | 6.5 b-d | 5.9 a | 5.0 a | 8.4 a | 0.60 a-c | 0.065 ab | 0.036 ab | 0.003 ab | ||
| 25 | 6.3 b-d | 4.1 a-e | 4.4 ab | 7.2 a-c | 0.65 ab | 0.074 ab | 0.028 bc | 0.004 ab | ||
| 30 | 5.8 c-e | 5.0 ab | 4.0 b | 6.7 a-c | 0.57 bc | 0.036 b | 0.028 bc | 0.002 ab | ||
| Significance | ||||||||||
| A (Air temp.) | *** | *** | *** | *** | *** | *** | ns | * | ||
| B (Root zone temp.) | ns | ns | ns | ns | *** | ns | ns | ns | ||
| A × B | ** | ns | ns | ns | *** | ns | ** | ns | ||
Table 3.
Growth characteristics of tomato seedlings as affected by different air temperatures and root zone temperatures 22 days after the sowing (n = 5, R = 3). Different letters indicate significant differences according to Duncan’s multiple range test at p < 0.05
|
Air temp. (°C) |
Root zone temp. (°C) |
Plant height (cm/plant) |
Root length (cm) |
No. of leaves (ea/plant) |
Leaf area (cm2/plant) |
Fresh weight (g/plant) |
Dry weight (g/plant) | |||
| Shoot | Root | Shoot | Root | |||||||
| 20 | 15 | 5.7 e | 5.1 c-e | 2.6 f | 12.9 g | 0.50 g | 0.11 bc | 0.039 h | 0.006 h | |
| 20 | 5.9 e | 6.2 b-e | 3.6 de | 30.0 d-g | 0.95 fg | 0.09 bc | 0.074 gh | 0.007 gh | ||
| 25 | 6.6 e | 4.1 e | 3.4 d-f | 24.3 e-g | 1.11 e-g | 0.07 c | 0.080 f-h | 0.005 h | ||
| 30 | 6.3 e | 4.6 de | 3.4 d-f | 20.4 fg | 1.15 e-g | 0.07 c | 0.084 f-h | 0.005 h | ||
| 25 | 15 | 10.5 d | 5.3 c-e | 3.4 d-f | 43.2 b-e | 2.28 de | 0.20 bc | 0.139 d-g | 0.016 d-f | |
| 20 | 10.4 d | 10.1 b | 3.4 d-f | 44.5 b-e | 2.58 cd | 0.26 bc | 0.161 d-f | 0.015 d-g | ||
| 25 | 9.6 d | 8.0 b-e | 3.2 ef | 42.3 b-e | 2.33 de | 0.22 bc | 0.150 d-g | 0.010 e-h | ||
| 30 | 10.6 d | 7.6 b-e | 3.4 d-f | 35.6 c-f | 1.92 d-f | 0.15 bc | 0.124 e-h | 0.011 e-h | ||
| 30 | 15 | 14.1 ab | 4.9 c-e | 4.0 de | 40.6 b-e | 2.50 cd | 0.13 bc | 0.171 de | 0.008 f-h | |
| 20 | 12.6 c | 8.8 bc | 4.2 d | 46.4 b-d | 2.98 cd | 0.30 b | 0.218 d | 0.017 de | ||
| 25 | 14.4 a | 8.4 b-d | 4.2 d | 41.8 b-e | 3.11 cd | 0.27 bc | 0.178 de | 0.019 d | ||
| 30 | 14.8 a | 5.3 c-e | 3.8 de | 33.8 c-f | 2.59 cd | 0.14 bc | 0.135 d-g | 0.008 f-h | ||
| 35 | 15 | 14.0 a-c | 18.4 a | 6.0 b | 58.4 ab | 4.66 ab | 0.24 bc | 0.347 c | 0.031 c | |
| 20 | 14.9 a | 17.6 a | 6.8 a | 66.3 a | 5.05 a | 0.66 a | 0.690 a | 0.072 a | ||
| 25 | 12.8 bc | 18.3 a | 5.2 c | 51.3 a-c | 4.67 ab | 0.67 a | 0.691 a | 0.065 b | ||
| 30 | 15.0 a | 16.4 a | 4.2 d | 44.0 b-e | 3.65 bc | 0.60 a | 0.559 b | 0.035 c | ||
| Significance | ||||||||||
| A (Air temp.) | *** | *** | *** | *** | *** | *** | *** | *** | ||
| B (Root zone temp.) | ns | * | *** | * | ns | *** | ** | *** | ||
| A × B | * | ns | *** | ns | ns | * | *** | *** | ||
Table 4.
Growth characteristics of tomato seedlings as affected by different air temperatures and root zone temperatures 29 days after the sowing (n = 5, R = 3). Different letters indicate significant differences according to Duncan’s multiple range test at p < 0.05
|
Air temp. (°C) |
Root zone temp. (°C) |
Plant height (cm/plant) |
Root length (cm) |
No. of leaves (ea/plant) |
Leaf area (cm2/plant) |
Fresh weight (g/plant) |
Dry weight (g/plant) | |||
| Shoot | Root | Shoot | Root | |||||||
| 20 | 15 | 11.0 e | 13.3 hi | 6.2 de | 127 d-f | 5.3 d | 0.79 c-e | 0.38 h | 0.029 fg | |
| 20 | 10.3 e | 20.3 de | 6.6 d | 178 bc | 8.5 c | 1.25 cd | 0.61 e-g | 0.056 c-f | ||
| 25 | 11.6 e | 14.8 gh | 6.0 d-f | 157 c-e | 7.7 cd | 0.64 de | 0.53 e-h | 0.034 e-g | ||
| 30 | 9.6 e | 9.8 i | 6.2 de | 115 ef | 6.4 cd | 0.44 e | 0.40 gh | 0.024 g | ||
| 25 | 15 | 17.5 d | 15.1 gh | 5.6 e-g | 157 c-e | 14.0 ab | 0.82 c-e | 0.46 f-h | 0.068 cd | |
| 20 | 16.9 d | 20.6 de | 5.8 d-g | 188 b | 17.0 a | 1.29 cd | 0.65 d-f | 0.100 b | ||
| 25 | 16.3 d | 19.2 ef | 5.2 f-g | 169 b-d | 14.2 ab | 0.93 c-e | 0.55 e-h | 0.082 bc | ||
| 30 | 16.6 d | 18.2 e-g | 5.2 f-g | 158 b-d | 12.4 b | 0.92 c-e | 0.41 gh | 0.072 b-d | ||
| 30 | 15 | 22.2 c | 12.6 hi | 5.0 g | 108 f | 5.9 cd | 0.32 e | 0.46 f-h | 0.023 g | |
| 20 | 22.3 c | 23.8 b-d | 6.0 d-f | 140 c-f | 8.9 c | 1.39 c | 0.74 c-e | 0.061 c-e | ||
| 25 | 23.8 a-c | 22.2 c-e | 6.0 d-f | 132 d-f | 8.5 c | 1.17 cd | 0.61 e-g | 0.071 b-d | ||
| 30 | 25.5 a | 16.0 f-h | 5.8 d-g | 139 c-f | 8.7 c | 0.59 de | 0.53 e-h | 0.042 d-g | ||
| 35 | 15 | 21.7 c | 24.1 b-d | 7.6 c | 159 b-d | 12.5 b | 1.00 c-e | 0.84 b-d | 0.073 bc | |
| 20 | 25.2 ab | 26.6 ab | 8.0 bc | 195 b | 14.9 ab | 2.31 ab | 1.15 a | 0.193 a | ||
| 25 | 24.9 ab | 25.6 ac | 9.0 a | 258 a | 15.9 a | 2.20 b | 1.02 ab | 0.174 a | ||
| 30 | 23.1 bc | 28.5 a | 8.6 ab | 235 a | 16.1 a | 2.92 a | 0.93 bc | 0.171 a | ||
| Significance | ||||||||||
| A (Air temp.) | *** | *** | *** | *** | *** | *** | *** | *** | ||
| B (Root zone temp.) | ns | *** | * | *** | *** | *** | *** | *** | ||
| A × B | ** | *** | * | ** | ns | *** | ns | *** | ||
근장의 경우 파종 후 15일부터 29일까지 대기온도와 근권부 온도 두가지 요인 모두 유의적으로 근장에 영향을 미치는 것으로 확인되었다(Tables 2, 3 and 4). 특히 대기온도 35°C 처리구는 다른 처리구에 비해 근장이 길었고 파종 후 29일에는 대기온도와 상관없이 근권부 온도 20°C 처리구에서 근장이 가장 길었다(Table 4). 오이의 경우 16°C의 근권부 온도에 노출될 경우 질산의 흡수율이 유의적으로 감소하여 뿌리의 생육이 억제되었으며 토마토의 경우에도 근권부 온도 13°C에서 양분의 흡수가 억제되어 생육이 불량해진다는 연구 결과가 보고된 바 있다(Bai et al., 2016; Lee et al., 2002). 본 연구에서도 근권부 온도를 15°C로 처리하였을 때 양분의 흡수가 억제되어 근장의 생육이 억제된 것으로 사료된다.
엽수와 엽면적의 경우 대기 온도는 파종 후 15일부터 29일까지 영향을 미쳤으며 대기온도가 증가할수록 엽수가 증가하는 경향을 보였다. 대기온도 35°C의 경우 근권부 온도를 20°C와 25°C로 조절하였을 때 토마토 묘의 엽수와 엽면적의 생육이 가장 좋았으나 근권부 온도를 15°C로 조절하였을 때 다른 처리구에 비해 생육이 억제되는 결과를 보였다(Tables 2, 3, 4 and Fig. 4). 토마토의 경우 근권부 온도는 뿌리 생육과 양수분 흡수에 영향을 미치며 대기온도가 증가함에 따라 엽수가 증가한다는 연구 결과가 보고된 바 있어 본 연구의 결과와 일치하였다(Mizuno et al., 2022).
지상부 생체중과 건물중은 대기온도 35°C가 다른 대기온도 처리구에 비해 가장 높았으나 파종 후 15일로 접어들수록 대기온도의 영향보다는 근권부 온도의 영향을 많이 받았다(Tables 2, 3 and 4). 파종 후 29일에는 모든 대기온도 처리구에 근권부 온도를 20°C와 25°C로 처리하였을 때 생체중과 건물중이 높은 경향을 보였으며, 특히 대기온도 25°C와 근권부 온도 20°C의 조합 처리구가 다른 처리구에 비해 가장 높았다(Table 4). Noh(1997)의 토마토 연구 결과에 따르면 근권부 온도가 21°C일 때 지상부 생체중과 건물중이 가장 높았고, 엽의 생체중과 건물중은 근권부 20°C에서 가장 높은 수치를 나타냈다고 보고된 바 있다(Lee at al., 2001). 본 연구의 결과도 Noh(1997)의 연구 결과와 비슷한 경향을 보였다.
지하부 생체중과 건물중의 경우 파종 후 15일부터 29일까지 대기온도의 영향을 많이 받았으며(Tables 2, 3 and 4), 대기온도 35°C 처리구에서 가장 높았다(Tables 2, 3 and 4). 전체적으로 근권부 온도는 생육이 후반기로 접어들수록 지하부 생체중과 건물중에 영향을 크게 미쳤다(Tables 2, 3 and 4). 특히 파종 후 29일에는 대기온도 30, 35°C 처리구에 근권부 온도를 20, 25°C로 처리하였을 때 지하부 생체중이 가장 높았다(Table 4). 토마토의 경우 근권부 온도 상승이 토마토 증산량과 호흡량의 증가로 인해 근권부 온도 40°C에서 지하부 생체중과 건물중이 가장 낮은 값을 보였으며, 근권부 25°C에서 가장 높은 값을 나타냈다고 하였다(Jang et al., 2015). 본 연구 결과에서도 근권부 온도 20, 25°C로 처리하였을 때 지하부 가장 높은 수치를 나타냈으며, 근권부 온도 30°C에서 가장 낮은 수치를 나타내 고온에서 증산량과 호흡량이 증가된 것으로 생각된다.
지상부 생체중과 건물중을 제외한 나머지 항목들은 대기 온도와 근권부 온도의 영향을 받는 것으로 나타났다. 특히 근장의 생육은 파종 후 15일 이후부터 대기온도와 근권부 온도의 영향을 많이 받았으며 대기온도 25, 30, 35°C에 근권부 온도를 20, 25°C로 설정하였을 때 토마토 초기 생육에 효과적이었다(Tables 2, 3 and 4). 그러나 대기온도 35°C의 경우 생육 후반기로 접어들수록 약간의 위조 증상이 관찰되어 이후 건전한 묘로 사용하기에 어려움이 있을 것으로 생각된다(Fig. 4). 높은 대기온도의 경우 과실의 수량을 감소시키고, 소형과 비율을 높였다는 결과가 보고된 바 있어(Lee et al., 2008), 대기 온도가 25, 30°C일 경우 근권부 온도를 20, 25°C로 처리하는 것이 가장 효과적일 것으로 판단된다.

대기온도와 근권부 온도가 생육에 미치는 영향을 ANOVA 다요인 분석한 결과 초장, 근장, 엽수, 엽면적, 생체중, 그리고 건물중 모두 대기온도의 영향을 가장 많이 받은 것으로 분석되었다(Tables 2, 3 and 4). 근권부 온도의 경우 초장을 제외한 생육특성은 처리 후 15일과 22일까지 근권부 온도의 영향을 받지 않거나 그 영향이 적었으나 처리 후 29일에는 근권부 온도의 영향을 많이 받은 것으로 분석되었다. 그리고 대기온도와 근권부 온도의 상호작용을 분석한 결과 지상부 생체중과 건물중을 제외한 모든 항목에서 두 요인의 상호작용이 고도로 유의하였다. 이러한 결과는 생육초기에 대기온도의 영향을 크게 받지만 혹서기와 같이 높은 대기온도 조건에서는 대기온도에 대응할 수 있는 요인이 근권부 온도 조절임을 시사하고 있다.
파종 후 29일째의 T/R율과 묘의 충실도는 Fig. 5에 그래프로 나타내었다. 과채류 육묘에 있어 묘의 품질을 나타내는 지표중의 하나로 T/R율은 낮은 값일수록 지하부의 건물중이 높아 지상부보다 지하부의 생육이 증가하여 근권의 발달이 우수한 묘라고 판단할 수 있다(Jeong et al., 2020). 충실도는 전체 건물중에 지상부의 초장을 나눈 값으로 그 값이 높을수록 묘의 품질이 우수하다고 보고되고 있다(Zhang et al., 2003; Lee et al., 2016). 본 연구에서는 대기온도 25°C의 처리구가 전체적으로 낮은 T/R율을 보였으며, 대기온도 30°C와 35°C에 근권부 온도를 15°C로 처리하였을 때 다른 근권부 온도 처리에 비해 높은 T/R율을 나타내었다. 오이 재배에서 양액온도를 16°C로 처리하였을 때 T/R율이 가장 높았으며 양액온도가 높아짐에 따라 감소하였다는 결과가 보고되었다(Lee, 1999). 본 연구에서도 모든 대기온도에 근권부 온도를 15°C로 처리하였을 때 다른 근권부 온도 처리구보다 T/R율이 높은 경향을 보여 근권부 온도를 15°C이하로 처리한다면 뿌리의 생장이 억제될 것으로 생각된다. 토마토 묘의 묘소질 지표로서 충실도를 보면 대기온도 20°C와 근권부 온도 20°C 처리구에서 가장 높은 충실도를 나타냈고, 근권부 온도가 낮은 15°C의 처리구에서는 모든 대기온도 처리구에서 가장 낮은 충실도를 나타냈다(Fig. 5). 이는 근권부 온도 15°C가 토마토 묘소질을 향상시키는데 한계 이하의 온도인 것으로 판단되며, 특히 근권부 온도 20–25°C로 조절한다면 묘의 품질향상에 도움이 될 것으로 생각된다.

Fig. 5.
Comparisons of the T/R (top/root) ratio and compactness of tomato seedlings as affected by different air temperatures (A.T.) and root zone temperatures 29 days after sowing (n = 5). Different letters and bars indicate significant differences according to Duncan’s multiple range test at p < 0.05.
토마토의 생장속도를 비교하기 위하여 CGR과 RGR을 Fig. 6에 그래프로 나타내었다. 대기온도 35°C의 모든 근권부 온도 처리구에서 CGR과 RGR 값이 모두 높게 나타났는데 이는 누적온도가 작물생육의 속도를 촉진하는 것으로 생각된다. 특히 모든 대기온도에 근권부 온도를 20°C로 처리하였을 때 CGR과 RGR값이 가장 높았다. 오이의 경우 육묘 20–30일 후에 근권부 온도를 25°C로 처리하였을 때 30°C 처리구에 비해 CGR과 RGR 값이 높은 값을 나타냈다(Lee et al., 2003). 본 연구 결과에서도 모든 대기온도에 근권부 온도를 15°C와 30°C로 처리하였을 때 CGR과 RGR 값이 낮아 과도한 고온이나 저온은 생장 속도를 감소시키는 것으로 생각된다. 토마토는 온대성 작물로서 생육기간 생육적온은 18.3–32.2°C 범위이고 최고온도는 30–35°C 이상 올라가지 않아야 하며, 토마토 육묘기에는 주간 24°C, 야간 18°C 범위로 보고되었다(Shamshiri et al., 2018). 본 실험에서도 선행연구와 비슷한 결과를 보였으며, 모든 대기온도에 근권부를 20–25°C로 처리하였을 때 생장율이 우수하였다.
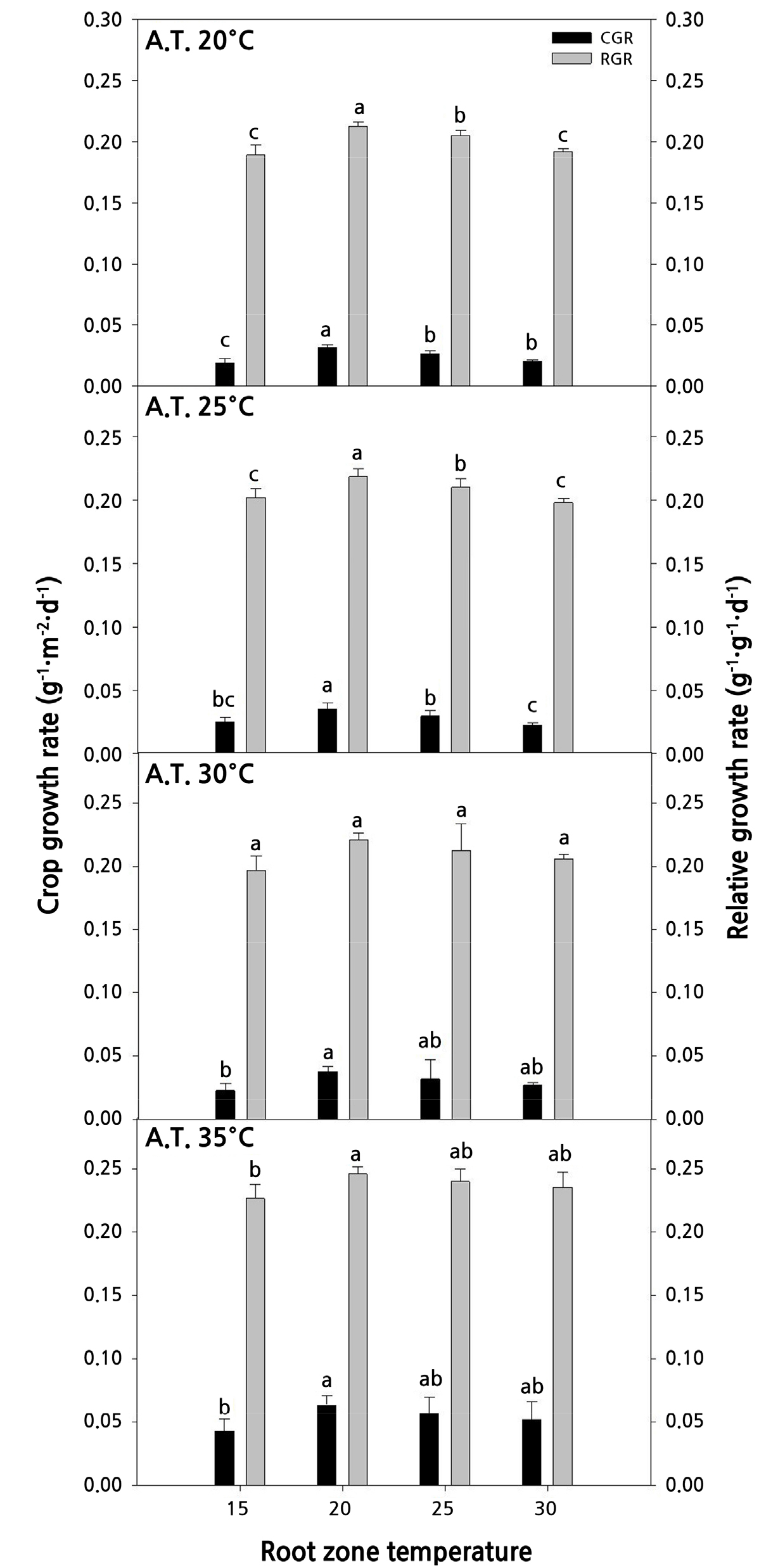
Fig. 6.
Comparisons of the CGR (crop growth rate) and RGR (relative growth rate) of tomato seedlings as affected by different air temperatures (A.T.) and root zone temperatures 29 days after sowing (n = 5). Different letters and bars indicate significant differences according to Duncan’s multiple range test at p < 0.05.
이상의 모든 결과를 종합해 보면 대기온도 25, 30°C에서 근권부 온도 20–25°C로 설정하였을 때 초기 생육에 효과적이었으며 대기온도가 35°C인 고온기에는 근권부 온도를 20–25°C로 설정한다면 고온에 의한 생리장해를 감소시킬 수 있을 것으로 판단된다. 또한 재배가 어려운 혹서기 및 혹한기에 양액 온도 조절을 통해 생육 촉진을 가능하게 할 뿐 아니라 근권부의 과도한 온도상승 및 하강을 방지하여 토마토의 건전묘 생산에 도움을 줄 것으로 생각된다.
