

Introduction
Materials and Methods
Seed preparation
Experimental design and growth conditions
Evaluation of growth characteristics
Leaf gas exchange measurements
Statistical analyses
Results and Discussion
Shoot and root characteristics
Growth analysis and leaf gas exchange
Principal component analysis
Conclusions
Introduction
The genus Angelica, known as a medicinal crop, belongs to the Umbelliferae (Apiaceae) family. It is widely used as a medicinal herb owing to its proven beneficial effects on the human body, including blood replenishment, blood circulation, and antioxidant effects (He et al. 2023). Three different Angelica species are mainly cultivated in northeast Asia: Korean angelica (Angelica gigas Nakai), Japanese angelica (A. acutiloba Kitagawa), and Chinese angelica [A. sinensis (Oliv.) Diels]. These three angelica species have been reported to be genetically and chemically distinct (Jeong et al. 2015; Kim et al. 2024b). In Korea, Korean and Japanese angelica are mainly cultivated, and among them, Korean angelica accounts for approximately 83% of the total angelica production in Korea as of 2023 (MAFRA 2024). Korean angelica is usually sown around April, and the seedlings are harvested in October-November and stored under cold storage conditions (RDA 2018). Cold-stored seedlings are transplanted to the field in April of the following year and harvested after the leaves fall (RDA 2018). Korean angelica is a cool-season crop mainly cultivated in mountainous areas at altitudes of 300–500 m above sea level and has been found to be sensitive to temperature increases (Yun et al. 2024).
Recently, potential crop yields have declined owing to increased temperatures and heat stress caused by climate change (Hatfield et al. 2011; Hasanuzzaman et al. 2013; Hatfield and Prueger 2015; Kim et al. 2024a; Yun et al. 2024). A common feature of heat stress responses is the inhibition of growth and development through the induction of defense mechanisms that consume the photosynthetic products necessary for survival (Hasanuzzaman et al. 2013; Zhang et al. 2022; Yeom and Oh 2023; Park et al. 2025). Within an appropriate temperature range, the plant growth speed is proportional to the increase in temperature; however, when the temperature rises beyond a critical point, growth is inhibited (Giri et al. 2017). Environments outside the optimal temperature range reduce the photosynthetic capacity of the plants, making it difficult to produce healthy, high-quality crops in the long term (Kim et al. 2024a; Kwak et al. 2024). High temperatures impair chlorophyll accumulation and increase respiration, reducing net photosynthesis and inhibiting the accumulation of primary metabolites (Dutta et al. 2009). Previous studies have reported that Korean angelica shows decreased growth rates and yields when grown at high temperatures above 28°C (Jeong et al. 2021; Park et al. 2025). However, mild heat stress at temperatures exceeding the optimal growth temperature by 1 to 4°C does not cause severe damage (Hatfield and Prueger 2015; Jang et al. 2024). Accordingly, it is necessary to determine the optimal temperature range that does not cause severe stress in each crop to prevent crop yield losses and achieve stable production.
A plant factory with an artificial lighting system (PFAL) is a cultivation space that minimizes the impact of external environmental conditions and artificially controls environmental factors such as the temperature (Lee et al. 2020), light conditions (Ju et al. 2023), and CO2 concentration (Yamaura et al. 2023) to produce uniform and high-quality crops (Kozai 2013). As climate change gradually worsens worldwide, the use of PFALs to control environmental factors is becoming increasingly important. Numerous studies have evaluated the environmental conditions suitable for plants so as to produce high-quality seedlings using PFALs. The transplantation cultivation of Angelica species using plug seedlings has been reported to be a useful strategy for shortening the cultivation period and preparing for abnormal climates (Lee et al. 2024). High-quality seedlings exhibiting vigorous root development show high rooting rates and environmental adaptability following transplantation, both of which are linked to increased production (Choi et al. 2023; Yamaura et al. 2023; Yun et al. 2025). If uniform and high-quality Korean angelica seedlings are produced in PFALs and supplied to farmers, it is expected that adaptability to climate change will increase and stable production will be realized. Information regarding the optimal temperature and temperature range is necessary to produce high-quality seedlings and avoid severe stress. In this study, we aimed to evaluate the growth characteristics of Korean angelica seedlings grown at different temperatures in PFALs to determine the appropriate temperature range for producing high-quality Korean angelica seedlings and to provide a temperature management method during the nursery period of these plants.
Materials and Methods
Seed preparation
Korean angelica (Angelica gigas Nakai) seeds were collected at Pyeongchang-gun (37° 52' N, 128° 48' E), Gangwon-do, Korea, in September of 2020. The collected seeds were stored in a cold storage facility maintained at 4°C until their use in the experiment. Korean angelica is a morphophysiological dormant seed with both ‘physiological dormancy’, where germination inhibitors exist within the seed, and ‘morphological dormancy’, where embryos are immature (Baskin and Baskin 2014; Zhang et al. 2023). Before the experiment, all seeds were washed under running tap water for 48 hours to remove any germination inhibitors (RDA 2018). Subsequently, to break morphological dormancy, the seeds were wrapped in wet gauze, placed in a plastic zipper bag, and stored in low-temperature storage maintained at 4°C for two weeks (Park et al. 2023). Before sowing, the seeds were subjected to a hot water treatment for 15 min at 45°C in a constant-temperature water bath (MSB-1511D, MonoTech Co. Ltd., Daegu, Korea) as a means of seed disinfection. After this treatment, the seeds were cooled under running tap water for 10 min.
Experimental design and growth conditions
The experiment was conducted at three PFALs (C1200H3, FC Poibe Co. Ltd., Seoul, Korea). All seeds were sown in a commercial medium (Plantworld, Nongwoobio Co. Ltd., Suwon, Korea) in 72-cell trays (35 mL/cell, Bumnong Co. Ltd., Jeongeup, Korea). For uniform germination, the internal temperature of all PFALs was maintained at 22/19°C (light/dark) for two weeks after sowing. Two weeks after sowing, the Korean angelica had fully developed cotyledons and the first true leaves had begun to grow. The plants were cultivated for 60 days, which included the two-week germination period.
The temperature treatment used here was based on previous research reporting that the optimal temperature range for Korean angelica seedlings was 20–24°C (Park et al. 2025). Temperatures inside PFALs were set at three different levels: 22/19°C (non-stress temperature condition), 25/22°C (suboptimal-temperature condition), and 28/25°C (high-temperature condition), to determine the range that did not cause severe high-temperature stress. The average light/dark temperature and relative humidity of each PFAL were maintained as follows: for the non-stress temperature condition 22.0 ± 0.1/19.1 ± 0.2°C, 59.1 ± 8.8%, for the suboptimal-temperature condition 24.9 ± 0.2/22.1 ± 0.1°C, 42.0 ± 12.7%, and for the high-temperature condition 28.0 ± 0.2/25.0 ± 0.1°C, 22.1 ± 6.8%. The experiment was designed to treat each plug tray as a replicate (72 plants per tray), and three trays were used per treatment. Temperatures inside each PFAL were measured using temperature data loggers (TR-76Ui, T&D Co. Ltd., Matsumoto, Japan) during the experimental period (Fig. 1).
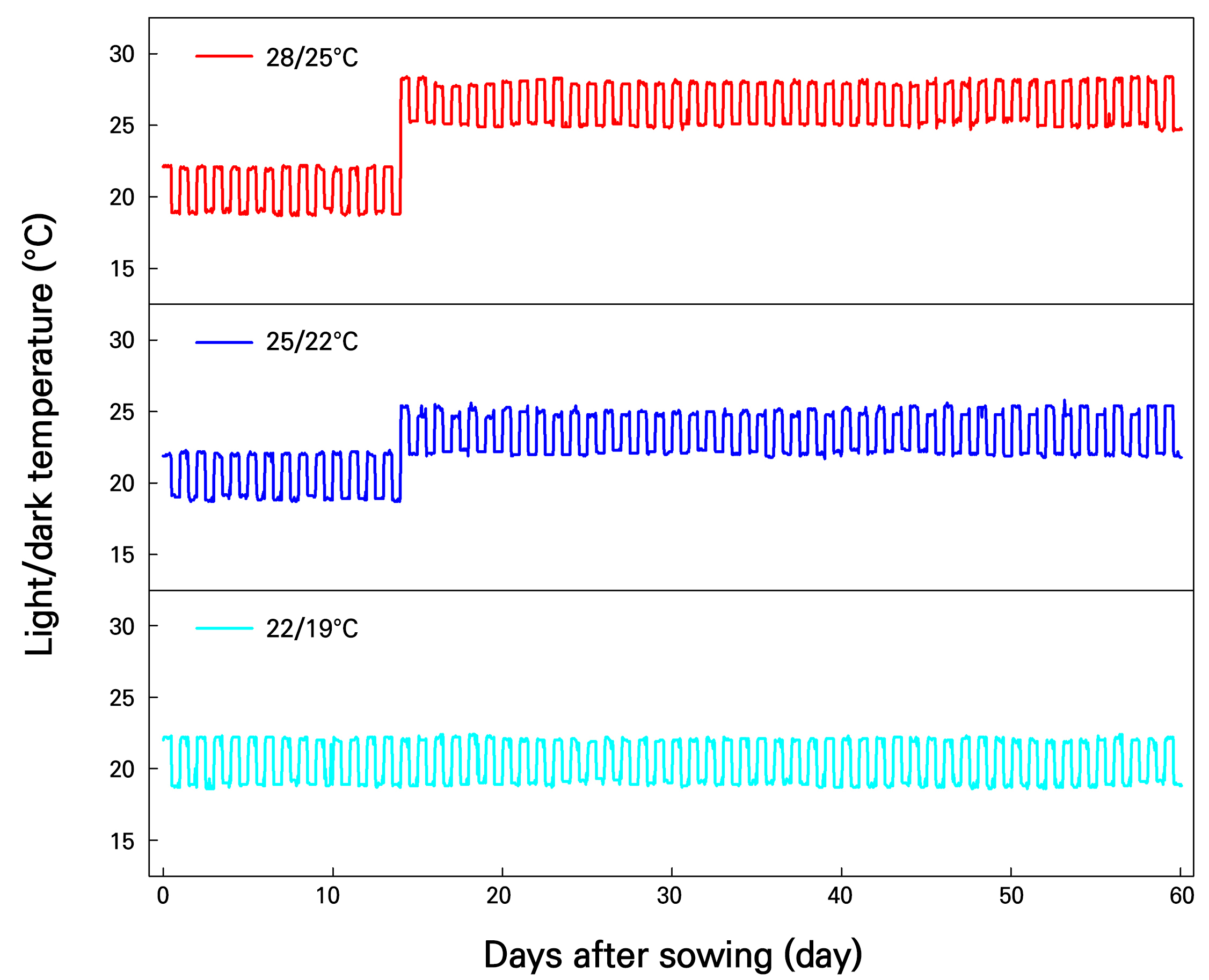
The light sources used were light-emitting diodes (Bissol LEDs Co. Ltd., Seoul, Korea), and the light quality was red : green : blue = 7 : 1 : 2. The light intensity was measured repeatedly ten times at the height of the growing point using a spectrometer (HD2102.2, Delta Ohm S.r.L., Caselle, Italy), and it was held at 255 ± 13 µmol·m-2·s-1 during the experiment period. The daily light intensity in the PFALs was set to 10.8 mol·m-2·d-1. The plug seedlings were sub-irrigated every two days with a multipurpose nutrient solution [(CaNO3)2·4H2O 472.0 mg·L-1, KNO3 202.0 mg·L-1, KH2PO4 272.0 mg·L-1, NH4NO3 80.0 mg·L-1, MgSO4·7H2O 246.0 mg·L-1, Fe-EDTA 15.0 mg·L-1, H3BO3 1.4 mg·L-1, MnSO4·4H2O 2.1 mg·L-1, ZnSO4·7H2O 0.8 mg·L-1, CuSO4·5H2O 0.2 mg·L-1, and Na2MoO4·2H2O 0.1 mg·L-1]. The electrical conductivity and pH were set to 1.0 dS·m-1 and 6.5, respectively.
Evaluation of growth characteristics
Growth characteristics, in this case the petiole length, leaf length and width, number of leaves, crown and root diameter, SPAD value, leaf area, and fresh and dry weights (leaf, stem, and root), were measured in triplicate for three individuals per tray. Each parameter was measured 40, 50, and 60 days after sowing (DAS). The petiole length was measured instead of plant height according to the morphological characteristics of Korean angelica. Leaves with a width greater than 1 cm were counted, except for the cotyledons. The crown and root diameters were measured at the thickest part using Vernier calipers (CD-20CPX, Mitutoyo Co. Ltd., Kawasaki, Japan). SPAD values of the leaves were measured using a chlorophyll meter (SPAD-502, Konica Minolta Inc., Osaka, Japan) with three replicates, and the average value was used for the analysis. Leaf area was measured using a leaf area meter (LI-3000; LI-COR Inc., Lincoln, NE, USA). Fresh and dry weights were measured using an electronic scale (WBA-220, Witeg Labortechnik GmbH, Wertheim, Germany). The dry weight was measured after drying the divided leaf, stem, and root samples for 72 hours in a drying oven (Venticell-222, MMM Medcenter Einrichtungen GmbH, Munich, Germany) at 70°C. The root was washed with tap water and scanned using a scanner (Epson Expression 12000XL, Epson Co. Ltd., Nagano, Japan) to evaluate the root morphology characteristics, which were in this case the total root length, root volume, and number of root tips. The images were analyzed using a root image analysis system (WinRhizo Pro 2020, Regent Instruments, Sainte-Foy, QC, Canada). To confirm differences in growth speed and dry matter accumulation based on the temperature conditions, the relative growth rate (RGR) and crop growth rate (CGR) were analyzed (Beadle 1993). RGR and CGR were calculated using the following equations:
where and represent the time as DAS, whereas and are the total dry weights of the plant at times and , respectively, under the condition of .
Leaf gas exchange measurements
The stomatal conductance (Gs), intercellular CO2 concentration (Ci), and net photosynthesis rate (Pn) were measured using a portable photosynthesis analyzer (CIRAS-3, PP Systems, Boston, MA, USA) at 60 DAS. Measurements were obtained from the second leaf of the plants with three replicates of two individuals per treatment. The leaf cuvette had a 4.5 cm2 window, and the light was provided by light-emitting diodes, set to 20% red, 50% green, and 30% blue. The cuvette conditions for photosynthesis measurements were set as follows: light intensity of 800 µmol·m-2·s-1, a CO2 concentration of 500 µmol·mol-1, and relative humidity of 55%. The temperatures in the cuvette were set to 22, 25, and 28°C, the daytime temperatures at which photosynthesis occurs.
Statistical analyses
The statistical analysis in this study was carried out using the statistical analysis software SAS 9.4 (SAS Institute Inc., Cary, NC, USA). One-way analysis of variance and Tukey’s range test were conducted on the data. Graphs were plotted using SigmaPlot 12.5 (Systat Software Inc., San Jose, CA, USA). Principal component analysis (PCA) in R (v. 4.2.1, R Foundation for Statistical Computing, Vienna, Austria) was used to examine the relationship between shoot and root growth characteristics. The PCA was carried out using the packages ggplot2, graphics, factoextra, and ellipse.
Results and Discussion
Shoot and root characteristics
The morphology of the Korean angelica plug seedlings was shaped by different temperature conditions within the PFALs (Fig. 2). At 40 DAS, no statistically significant differences were observed in any of the shoot growth parameters, except for the petiole length, between the treatments (Table 1). The petiole length did not show a clear trend throughout the nursery period. At 50 and 60 DAS, crown diameters, leaf areas, and dry weights were lowest under the 28/25°C condition, with no significant differences between the 22/19°C and 25/22°C conditions. The SPAD values tended to decrease with an increase in the temperature at 60 DAS.

Table 1.
Shoot growth characteristics of Korean angelica affected by different temperature conditions at 40, 50, 60 days after sowing (n = 3)
|
Days after sowing (days) |
Temperature (light/dark) |
Petiole length (cm) |
Leaf length (cm) |
Leaf width (cm) |
No. of leaves (/plant) |
Crown diameter (mm) |
SPAD value | Leaf area (cm2) | Dry weight (mg) | |
| Leaf | Stem | |||||||||
| 40 | 22/19°C | 2.8 abz | 1.6 a | 1.6 a | 2.0 a | 2.24 a | 32.6 a | 6.5 a | 24.0 a | 10.5 a |
| 25/22°C | 3.5 a | 1.6 a | 1.7 a | 2.2 a | 2.45 a | 31.4 a | 6.6 a | 26.5 a | 9.2 a | |
| 28/25°C | 2.6 b | 1.4 a | 1.4 a | 1.9 a | 2.21 a | 29.2 a | 5.1 a | 19.8 a | 8.9 a | |
| 50 | 22/19°C | 7.0 az | 2.8 a | 2.8 a | 3.0 a | 4.58 a | 35.2 a | 23.1 a | 91.6 a | 32.3 a |
| 25/22°C | 5.7 b | 2.8 a | 2.8 a | 3.0 a | 5.03 a | 34.0 a | 21.4 a | 88.9 a | 35.8 a | |
| 28/25°C | 5.2 b | 2.4 b | 2.3 b | 2.9 a | 3.55 b | 32.1 a | 13.7 b | 60.3 b | 20.9 b | |
| 60 | 22/19°C | 9.3 az | 4.7 a | 4.2 a | 3.2 a | 7.18 a | 35.8 a | 56.8 a | 200.4 a | 85.1 a |
| 25/22°C | 7.8 a | 4.4 a | 3.8 a | 3.2 a | 6.79 a | 34.2 ab | 52.3 a | 194.3 a | 68.5 ab | |
| 28/25°C | 7.6 a | 3.4 b | 3.0 b | 3.1 a | 5.67 b | 32.0 b | 31.3 b | 121.9 b | 48.7 b | |
With regard to the root characteristics, the total root length at 40 DAS was not different between 22/19°C and 25/22°C and was lower in the 28/25°C condition (Fig. 3A). However, there were no significant differences between the treatments at 50 and 60 DAS. The root diameter, volume, dry weight, and number of root tips consistently tended to decrease as the temperature was increased (Fig. 3B, 3C, 3D, and 3E). The root : shoot ratios, representing the dry weight proportions of roots and shoots, tended to be higher at low temperatures relative to those at high temperatures (Fig. 3F).

Fig. 3.
Changes in the root morphology characteristics and root dry weight of Korean angelica as affected by different temperature conditions at 40, 50, and 60 days after sowing. Scatter points represent the means, and error bars indicate the standard errors (n = 3). Different lowercase letters near the data points indicate significant differences according to Tukey’s range test at p < 0.05.
Growth analysis and leaf gas exchange
The RGR and CGR, which correspondingly indicate the growth rate and dry matter accumulation per unit time of Korean angelica based on the temperature conditions, were analyzed, as shown in Fig. 4. RGR did not show statistically significant differences across any of the measurement time points or treatments (Fig. 4A). Regarding the CGR outcomes, there was a tendency for dry matter accumulation to decrease as the temperature was increased in the 40–50 DAS interval. In the 50–60 DAS interval, dry weight accumulation was lowest under the 28/25°C, with no difference between the 22/19°C and 25/22°C conditions (Fig. 4B).
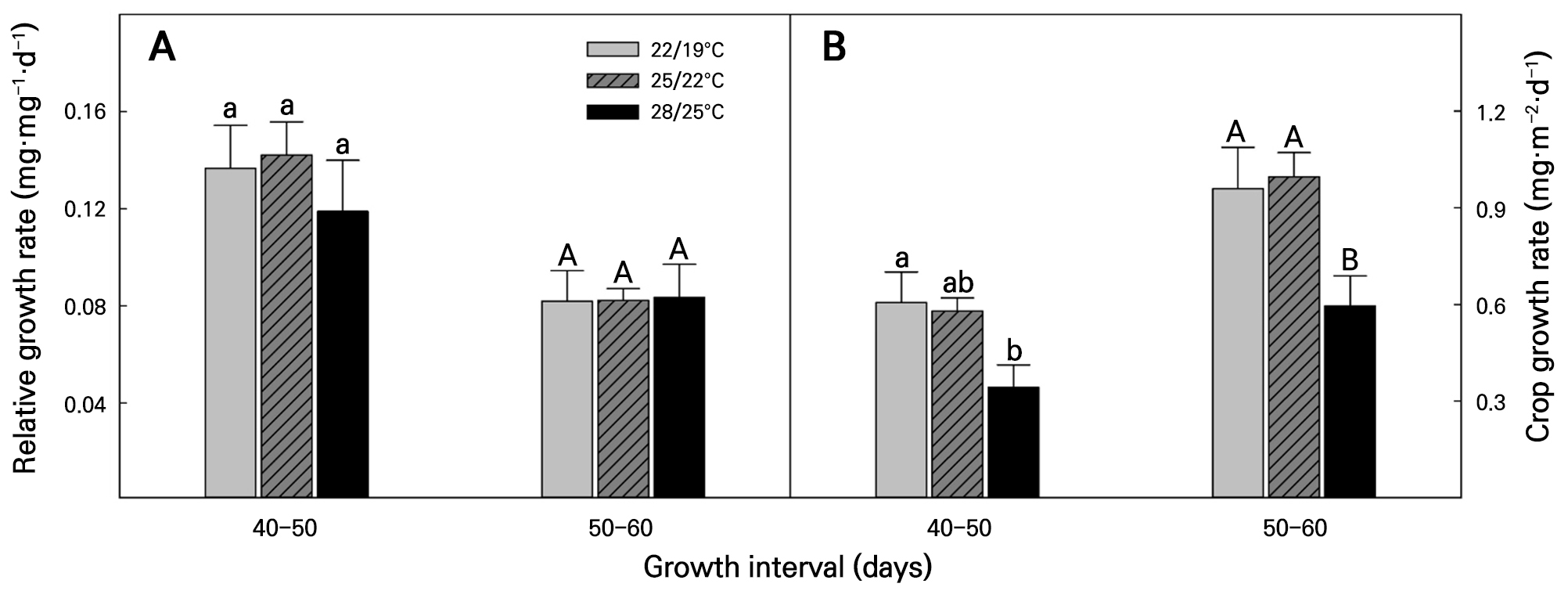
Fig. 4.
Relative growth rate (A) and crop growth rate (B) of Korean angelica as affected by different temperature conditions. Vertical bars represent the means and error bars indicate the standard errors (n = 3). Different lowercase and uppercase letters above the error bars indicate significant differences based on Tukey’s range test at p < 0.05. Lowercase letters indicate significant differences in means across temperature conditions between 40 and 50 days after sowing. Uppercase letters indicate significant differences in means across temperature conditions between 50 and 60 days after sowing.
A comparison of the leaf gas exchange in Korean angelica leaves in response to the different temperature conditions is shown in Fig. 5. Gs and Ci did not significantly differ at any temperature (Fig. 5A and 5B). However, Pn tended to decrease as the temperature was increased, and the Pn value of Korean angelica seedlings grown at 22/19°C was significantly higher than the corresponding value for those grown at 28/25°C (Fig. 5C).
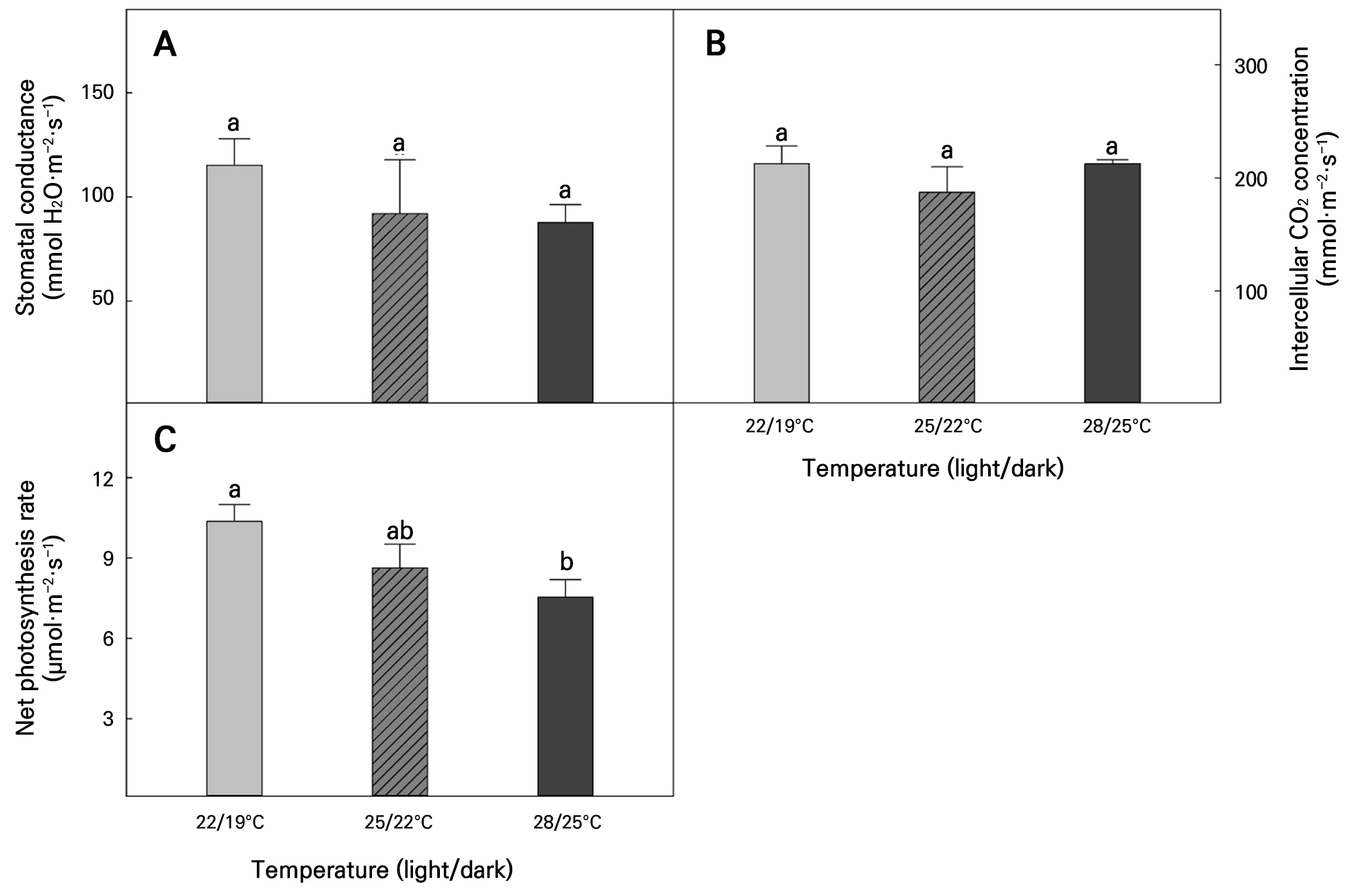
Fig. 5.
Stomatal conductance (A), intercellular CO2 concentration (B), and net photosynthesis rate (C) in Korean angelica as affected by different temperature conditions at 60 days after sowing. Vertical bars represent the means, and error bars indicate the standard errors (n = 3). Different lowercase letters above the error bars indicate significant differences based on Tukey’s range test at p < 0.05.
The level of leaf gas exchange is closely related to stomatal opening and closure, which is a typical physiological response that occurs when plants experience drought or high-temperature stress (Hatfield and Prueger 2015; Wang and Dang 2023). Plants close their stomata to suppress water loss by inhibiting transpiration under drought and high-temperature stress conditions (Zhang et al. 2022). Stomatal closure is among the survival strategies of plants; however, it inhibits CO2 inflow through the stomata. In addition, transpiration is minimized, which inhibits nutrient absorption through differences in the water potential (Giri et al. 2017; Onwuka and Mang 2018). Ultimately, these stress symptoms inhibit plant growth and development. However, in our study, the Gs and Ci values of Korean angelica did not show significant differences with an increase in the temperature or a decrease in the relative humidity (Fig. 5A and 5B), implying that they were not subjected to stress that threatened their survival within the temperature range in this case. The inference that ‘no serious stress was experienced in the temperature condition in this case’ is also suggested in the results showing that RGR was maintained without significant differences among the temperature treatments (Fig. 4A). Plants often exhibit differences in optimal temperature conditions within species or varieties (Hwang et al. 2024). In addition, although not fatal, they display a consistent physiological response of slow growth when exposed to inappropriate environments (Hasanuzzaman et al. 2013; Jeong et al. 2021; Zhang et al. 2022; Park et al. 2025). In particular, under high-temperature conditions, this can be understood as a mechanism for decreasing net photosynthesis because respiration increases more rapidly than photosynthesis (Hatfield and Prueger 2015). This is consistent with the result showing that CGR, which represents absolute dry matter accumulation, was significantly lower under the high-temperature conditions. However, given that no severe growth stress, such as organ damage, was experienced, a constant growth rate was maintained, and there was no significant difference in the RGR outcomes.
Principal component analysis
PCA, a multivariate statistical technique, was used to identify factors highly correlated with shoot and root development (Fig. 6). PCA reduces the dimensionality of the data by transforming existing variables into principal components (PCs) and preserves the variation of the entire dataset in only a few PCs (Liu et al. 2020). According to the PCA results, the first two PCs account for 89.1% of the total variations (PC 1 + PC 2). This method can efficiently and visually verify correlations between variables. Strong correlations were observed between the petiole length, crown diameter, leaf area, dry weight (leaf, stem, root, and total), root diameter, and root volume. Among these, root development parameters, i.e., the root volume, diameter, and dry weight, showed strong correlations with the crown diameter, leaf area, and leaf and stem dry weights. A strong correlation between the crown diameter and dry weight is speculated because the crown is the growth point where assimilation products accumulate (Macías-Rodríguez et al. 2002). However, non-destructive measurements of the crown diameter and petiole length are challenging as both the crown and petiole are covered by leaves or hidden by surrounding plants in tray units. The leaf is the plant organ commonly most utilized for non-destructive analyses of physiological changes, including the leaf area index (Carlson and Ripley 1997), leaf water content (Jackson et al. 2004), nitrogen content (Ali et al. 2017), and for disease diagnosis (Zhao et al. 2022). The strong correlation between leaf area and root development as identified in the PCA results suggests that using the leaf area to predict future root development in Korean angelica is feasible.
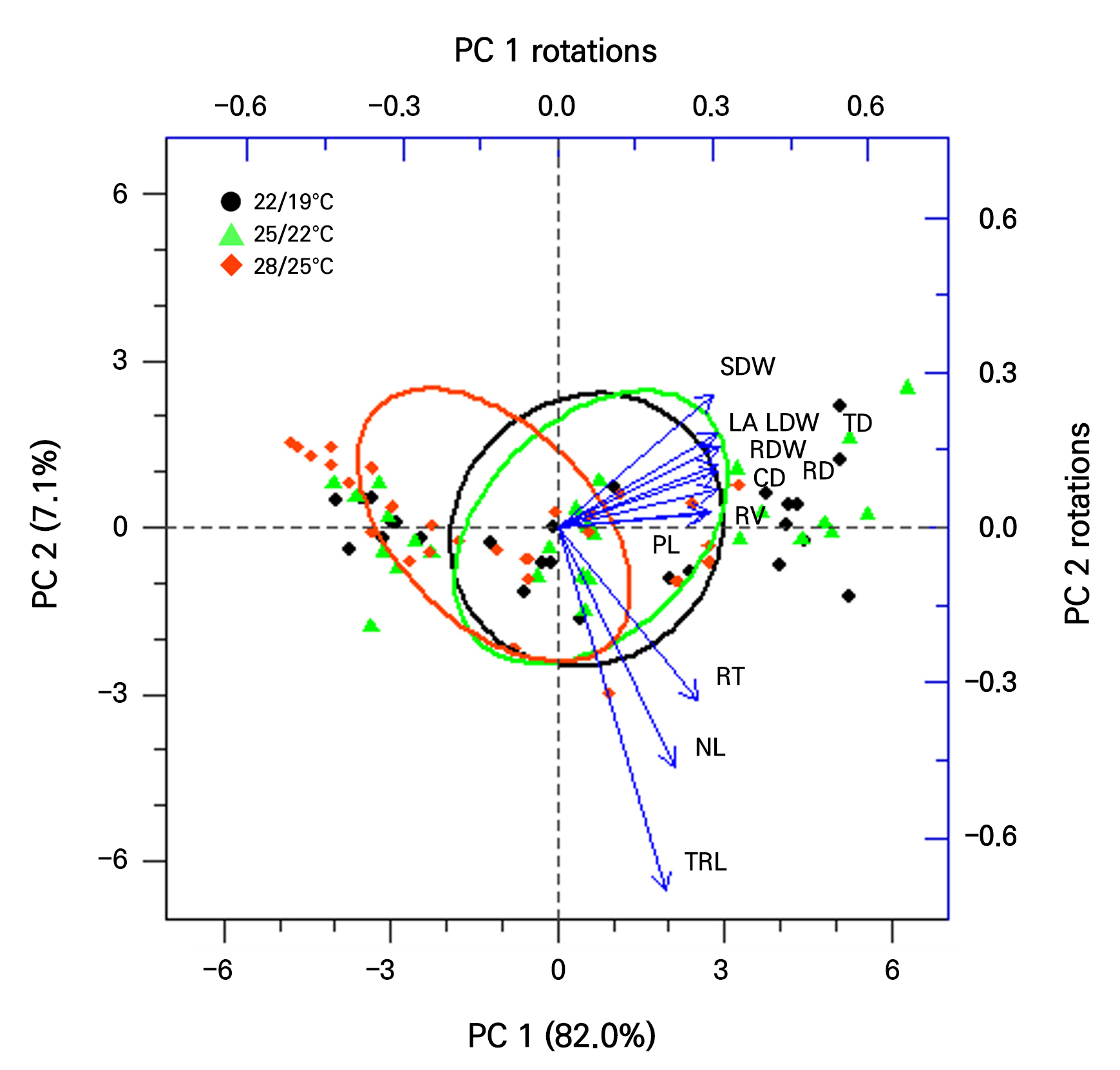
Fig. 6.
Principal component analysis of the petiole length (PL), number of leaves (NL), crown diameter (CD), leaf area (LA), leaf dry weight (LDW), stem dry weight (SDW), root dry weight (RDW), total dry weight (TD), and root morphology indicators, where TRL represents the total root length, RD represents the root diameter, RV represents the root volume, and RT represents the number of root tips. The length of the vectors indicates the relative proportion of variability in each variable. The angle between the vectors indicates a positive or negative correlation between each variable. The location of each point represents the principal component score calculated through the principal component analysis.
Conclusions
When the results of this study were summarized, 22/19°C was determined to be the most appropriate temperature for Korean angelica seedlings. However, there was no significant difference from the 25/22°C condition, and considering economic temperature management in the nursery facility, 19–25°C was determined to be the appropriate temperature range. PCA results confirmed the correlation between shoot and root development, which suggests an area for further study in this field. Additionally, it suggests the need for future studies focusing on environmental control, including appropriate humidity levels and carbon dioxide concentrations according to temperature changes, as well as nutrient movement. These results can be utilized by farms or nurseries that cultivate Korean angelica and can contribute to the production of Korean angelica in the future.
