

Introduction
Materials and Methods
Plant material and experimental sites
PGR application
Visual assessment of phytotoxic symptoms
Classification and characteristics of fruiting shoots
Fruiting shoot number and morphological measurements
Assessment of fruit set across fruiting shoot types
Yield and fruit/stone characteristics
Experimental design and statistical analysis
Results
Baseline morphological characteristics of fruiting shoots according to the positional category in control trees
Final fruit set and fruit-set ratio per floral cluster
Fruit set and fruit-set ratio based on total fruiting shoots per tree
Yield per tree and marketable yield
Fruit and stone characteristics
Phytotoxic symptoms
Discussion
Introduction
Jujube (Ziziphus jujuba Mill.) is a widely cultivated deciduous fruit tree in Korea and China, valued for fresh consumption, bioactive compounds, and medicinal applications (Liu et al. 2020; Kim et al. 2021). In recent decades, both cultivation and research have expanded globally, reflecting increasing interest in jujube production (Crawford et al. 2013; Yao 2013; Stănică 2019; Acarsoy Bilgin 2020; Ghouth 2022; Khadivi and Beigi 2022).
In Korea, jujube has long been recognized as a functional component of traditional medicine (Ji et al. 2014; Agrawal et al. 2023). Recent increases in demand for fresh fruit and product diversification have further expanded its cultivation and consumption (Kim et al. 2021; Park et al. 2022a; Park et al. 2022b; Park et al. 2023; Lee et al. 2025). According to the Korea Forest Service (KFS 2025), national production reached 7,666 tons in 2024, with a production value of 52.53 million USD. Production is concentrated in Boeun and Gyeongsan. Gyeongsan recorded higher output (2,379 tons), primarily from dried jujube, whereas Boeun produced less fruit (1,239 tons) but generated higher fresh-fruit revenue (14.60 vs. 6.46 million USD). The number of jujube-growing households (52,145) and the total cultivation area (6,487 ha) have continued to increase in recent years (MAFRA 2024).
Jujube exhibits profuse flowering and strong parthenocarpy (Ackerman 1961; Liu et al. 2008), with a prolonged blooming period of approximately ten weeks (Sun et al. 2009; Hernández et al. 2015; Nam et al. 2015; Yao et al. 2015). In Korea, jujube generally flowers from mid-June to early August, although the onset and duration of flowering can vary depending on the cultivar, tree age, pruning intensity, and local orchard conditions. However, achieving stable fruit set remains challenging because this flowering period overlaps with the summer monsoon season (KMA 2022), when frequent rainfall and reduced solar radiation often result in physiological fruit drop and yield losses (KREI 2023; KREI 2024).
Various approaches have been explored to mitigate poor fruit set under these conditions, including pollinator release, nutrient supplementation, root pruning, girdling, and the application of plant growth regulators (PGR) (Paik 1972; CARES 2017b; Lee et al. 2017; Oh et al. 2017; Fan et al. 2022; Ran et al. 2022; Tao et al. 2024). Despite partial success, most approaches have seen limited commercial adoption due to the intense labor required or the lack of well-defined application protocols.
Among PGRs, gibberellic acid (GA3) and auxin-like compounds such as 4-chlorophenoxyacetic acid (4-CPA) have been widely used to regulate fruit set and early fruit development in several horticultural crops. For example, gibberellins are associated with ovary growth and parthenocarpic fruit development in seedless mandarin (Talon et al. 1992), and GA3 is commonly used in grape production to induce seedlessness, regulate seed development, and increase berry growth and/or cluster elongation (Cheng et al. 2013; Acheampong et al. 2017; Jung and Hur 2020). In contrast, 4-CPA and related auxin-like compounds have been used to promote fruit set under unfavorable conditions; in eggplant, the PGR 4-CPA induced fruit set and fruit development under low-light and low-temperature winter conditions (Lee et al. 2004), while in tomato, combined 4-CPA and gibberellin treatments reduced high-temperature inhibition of fruit set and increased the number of normal fruits (Sasaki et al. 2005). Thus, GA3 and 4-CPA were selected in the present study to compare gibberellin- and auxin-related approaches for improving fruit retention during flowering.
In jujube, GA3 is linked to enhanced pollen performance and fruit-set-related hormonal regulation (Ayimaiti et al. 2020; Fan et al. 2022). However, little is known about whether fruiting shoots with different positional origins respond differently to a PGR application under open-field conditions. Jujube fruiting shoots are deciduous annual shoots that develop on current-season shoots, and their developmental timing and flowering sequence can differ according to positional origin (Liu et al. 2006; Yao 2013). In woody perennial fruit crops, shoot origin and position can influence vascular connectivity, local assimilate supply, and the source–sink balance (Costes et al. 2006). Therefore, we hypothesized that Fs1–Fs3 would differ in terms of the flowering sequence, early fruit-retention capacity, and responsiveness to exogenous PGRs during flowering.
Most previous studies evaluated PGR responses at the canopy or on the whole-tree scale without adequately accounting for structural variations among fruiting shoots (CARES 2017b; Ayimaiti et al. 2020; Mishra et al. 2020; Fan et al. 2022). Fruiting shoots in jujube originate from different positional locations and exhibit distinct flowering intensity levels, growth patterns, and vegetative–reproductive balances (Yao 2013; Liu et al. 2020; Song et al. 2021; Bao et al. 2022). However, these differences have rarely been incorporated into fruit-set evaluations.
In this study, jujube fruiting shoots were classified into three positional types based on their origin on current-season shoots. GA3, 4-CPA, and combined GA3 + 4-CPA treatments were applied three times at two-week intervals beginning at approximately 10% bloom, and positional differences in fruit-set responses and fruit quality were evaluated under open-field conditions at two major production sites. This study aimed to evaluate whether flowering-stage applications of GA3, 4-CPA, and a combination of these acids can improve fruit set and yield outcomes in open-field jujube and to determine whether these responses differ according to the fruiting-shoot positional origin.
Materials and Methods
Plant material and experimental sites
The experiment was conducted in 2024 using ‘Bokjo’, a major fresh-market jujube cultivar in Korea. Field trials were established at two open-field sites in Chungcheongbuk-do, Korea: the experimental orchard of the Jujube Research Institute, Chungcheongbuk-do Agricultural Research and Extension Services, Boeun, and a commercial orchard in Goesan. The Boeun site was located at 36°34'37.4" N, 127°44'53.7" E, whereas the Goesan site was located at 36°40'25.5" N, 127°46'04.3" E. Trees were planted at 3 × 4 m in Boeun and 2 × 4 m in Goesan, expressed as row spacing × tree spacing. Trees at both sites were grown under open-field conditions and trained to a modified central leader system with heavy winter pruning for fresh-fruit production. In Boeun, 13–15-year-old trees with six to seven primary scaffold branches were used, whereas in Goesan, 8–10-year-old trees with five to six scaffold branches were used. Fertilization, irrigation, and pest and disease management followed regional jujube cultivation practices and were applied uniformly within each site. No other PGRs were used during the experimental period. At each site, 35 trees were used for the PGR experiment, consisting of seven treatments with five replicate trees per treatment.
PGR application
Seven treatments were evaluated in this study: an untreated control treatment, GA3 at 25 ppm (GA1), GA3 at 50 ppm (GA2), 4-CPA at 10 ppm (CPA1), 4-CPA at 20 ppm (CPA2), GA3 + 4-CPA at 20 + 10 ppm (GCPA1), and GA3 + 4-CPA at 40 + 20 ppm (GCPA2). GA3 was applied using a 2.5% commercial formulation (Illidine; Jangyu Industry, Korea), whereas 4-CPA was applied using a 0.15% commercial formulation (Tomadoton; FarmHannong, Korea). The combined GA3 + 4-CPA treatments were applied using a premixed commercial formulation containing 2.5% GA3 and 1.25% 4-CPA (Chakgwal-Ring; Jangyu Industry, Korea). All concentrations are expressed as ppm in the final spray solution. Because the GA3 concentrations differed between the GA3-alone and combined commercial formulations, the combined treatments were evaluated as commercially available premixed PGR formulations rather than as factorial combinations designed to isolate the additive or synergistic effect of 4-CPA at identical GA3 concentrations.
Treatments were applied three times during the flowering period, beginning at approximately 10% bloom at each site and repeated at two-week intervals. Because jujube has a prolonged and asynchronous flowering habit, the spray schedule was determined based on the flowering stage of each orchard rather than on identical calendar dates. Accordingly, applications were conducted on June 12, June 25, and July 12 in Boeun, and on June 24, July 12, and July 26, 2024, in Goesan. A motorized sprayer (DHP20; Daewon Co., Japan) was used to thoroughly wet canopies at 1,600 L ha-1, equivalent to approximately 1.8 L tree-1. Applications were conducted between 09:00 and 10:00 on clear mornings. Daily precipitation, the daily maximum air temperature, the daily minimum air temperature, and the daily mean air temperature during the flowering and PGR application period were obtained from the Korea Meteorological Administration weather station nearest to each experimental site (KMA 2024). These data are provided in Supplementary Table S1.
Visual assessment of phytotoxic symptoms
Visual phytotoxic symptoms were assessed 14 days after each PGR application, corresponding to DAT14-1, DAT14-2, and DAT14-3. Symptoms were evaluated based on leaf tip burn, inward leaf curling, calyx browning or necrosis, and reduced flower opening. Injury severity was rated for each experimental tree on a 0–5 ordinal scale according to the approximate proportion of the whole tree showing visible symptoms. The raw scores, median, and range are provided in Supplementary Table S2.
Classification and characteristics of fruiting shoots
Fruiting shoots were classified into three positional types (Fs1–Fs3, Fruiting shoot 1–Fruiting shoot 3) based on their developmental origin of current-season shoots (Fig. 1A and 1B). Fs1 shoots arise from the basal portion of each bud, Fs2 shoots arise from the main axis (stem), and Fs3 shoots are from lateral branches.
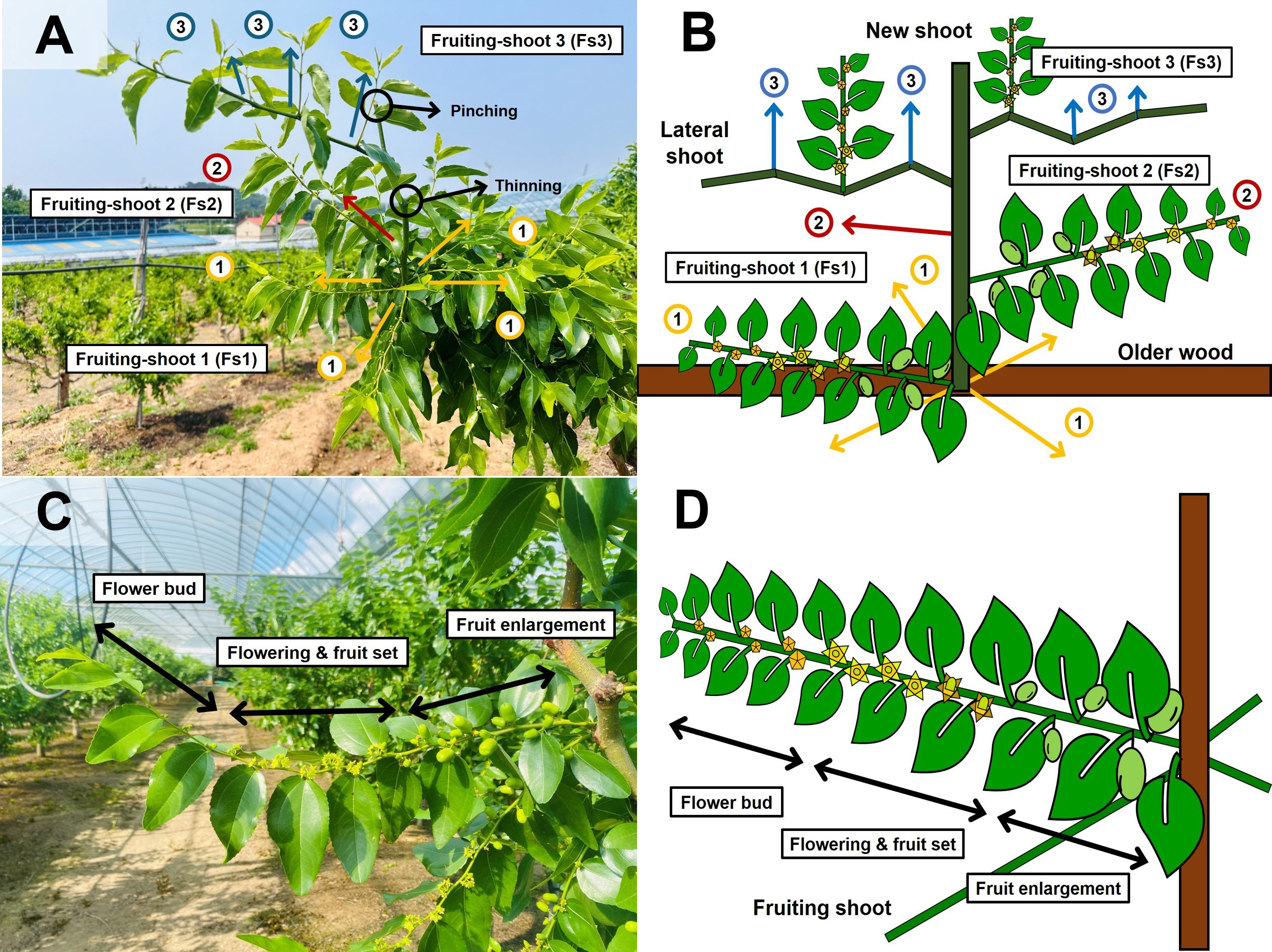
Fig. 1.
Classification and morphological characteristics of fruiting-shoot types according to positional origin in jujube: (A) Positional arrangement of fruiting shoots on current season shoots. (B) Schematic illustration of the three positional types: Fs1 (basal bud origin), Fs2 (main axis origin), and Fs3 (lateral branch origin). (C) Fruiting shoots during the flowering and fruit set stages. (D) Structural schematic illustrating key components of fruiting shoots.
Fruiting shoot number and morphological measurements
Measurements were conducted on trees that had not undergone shoot thinning. For fruiting shoot number (Table 1), five control trees per site were evaluated by complete census, counting Fs1–Fs3 shoots at the whole-tree level. For flowering phenology and morphological traits, stratified random subsamples were collected from the same trees across canopy layers (upper, middle, lower) and aspects. Ten shoots per positional type (30 tree-1) were tagged for monitoring. Flowering phenology was recorded weekly at the whole-tree level from the first open flower to flowering end. Morphological traits were measured at the fruit-coloring stage using uniform maturity criteria across sites. Subsample data were averaged according to the tree prior to analysis (n = 5 trees site-1).
Table 1.
Flowering and morphological characteristics of fruiting shoots according to positional type in control trees
Measurements were conducted in untreated control trees and were not used to assess PGR treatment effects. Fs1–Fs3 denote the three positional types of fruiting shoots (basal bud origin, main axis origin, and lateral branch origin of current-season shoots), respectively. Values are means ± SD (n = 5 control trees per site). Days indicate the number of calendar days from the first to the last flowering date (inclusive). The number of fruiting shoots was obtained by whole tree census and was not subjected to statistical comparisons. Within each site, different letters indicate significant differences among the fruiting shoot positions for length, number of leaves, and number of floral clusters according to Tukey’s HSD test (p < 0.05).
Assessment of fruit set across fruiting shoot types
Fruit set was assessed at five time points: pre-treatment (approximately 10% bloom), 14 days after the first spray (DAT14-1), 14 days after the second spray (DAT14-2), 14 days after the third spray (DAT14-3), and final harvest (fruit-coloring stage). At pre-treatment, fruit set was defined as visibly retained fruitlets (ovary-inflated stage) per tagged shoot; nonzero values could occur due to phenological variations at tagging. Observations followed the spray schedule (Boeun: June 12, June 25, July 12; Goesan: June 24, July 12, July 26, 2024), with final assessment at fruit coloring (Boeun: September 3; Goesan: September 6, 2024). Five trees per treatment were monitored (experimental unit: tree; n = 5), with ten shoots per positional type (Fs1–Fs3) randomly selected and tagged per tree (30 shoots tree-1; 150 shoots treatment-1 site-1).
Due to jujube's asynchronous flowering habit, fruit counts represent retained fruitlets at each stage (fruit set minus abscission). Measurements were stage-specific, not longitudinal tracking of fixed cohorts (Supplementary Table S3). Statistical comparisons used tree-means ANOVA at each stage within sites.
Fruit set was quantified by the following two indices:
Fruit set A (Fruiting shoot-level, %) = number of fruits set / number of floral clusters × 100
Fruit set B (Tree-level, %) = number of fruits set / number of fruiting shoots × 100
Fruit set was evaluated using two complementary indices to account for between-tree variation in fruiting shoot number and composition. Fruit set A (fruits per floral cluster) captures positional responses (Fs1–Fs3), whereas Fruit set B (fruits per fruiting shoot) provides whole-tree validation. These indices distinguish PGR effects from orchard-structure variations.
Yield and fruit/stone characteristics
Fruits were harvested in early October from five trees per treatment at each site. All fruits from each experimental tree were collected and weighed using an electronic balance (CAS FS-250; CAS Corp., Seoul, Korea) to determine the total yield. After total yield measurements, the harvested fruits were separated into marketable and non-marketable categories, and each category was weighed separately. Non-marketable fruits were defined as fruits affected by cracking, shriveling, disease, or insect damage. Fruit and stone quality assessments were conducted using only marketable fruits according to methods set by the Research Standards for Agricultural Science and Technology Investigation (RDA 2012).
For each treatment, 150 marketable fruits were randomly sampled for fruit and stone evaluations, with 30 fruits collected from each of five trees. Fruit and stone dimensions, in this case the length and width, were measured using digital calipers (CD-15 CPX; Mitutoyo, Kawasaki, Japan). Pedicel length was also measured using digital calipers. Fruit and stone weights were measured using an electronic balance (XT 4200C; Precisa, Dietikon, Switzerland). Post-treatment fruiting-shoot length and number of leaves were measured from the same tagged shoots at the fruit-coloring stage and averaged at the tree level before the statistical analysis. The soluble solids content (SSC, °Brix) was measured using a portable digital refractometer (PAL-1; ATAGO Co., Ltd., Tokyo, Japan), and fruit firmness was determined using a penetrometer (KM-5; Fujiwara, Tokyo, Japan). Measurements from the 30 fruits per tree were averaged to obtain one tree-level mean. Thus, five tree-level means per treatment at each site were used for the statistical analysis.
Experimental design and statistical analysis
At each site, treatments were arranged in a randomized complete block design (RCBD), with orchard rows used as blocks. Individual trees served as experimental units, with five trees per treatment at each site (Supplementary Fig. S1). Within each tree, ten fruiting shoots per positional type (Fs1–Fs3) were randomly selected and monitored, resulting in 30 tagged fruiting shoots per tree and 150 tagged fruiting shoots per treatment at each site. For fruit and stone quality assessments, 30 marketable fruits were sampled from each tree. Subsample measurements at the shoot or fruit level were initially averaged within each tree. The resulting five tree-level means per treatment at each site were used in the statistical analysis to avoid pseudoreplication. Analyses were conducted separately for each site. For time-course fruit-set data, statistical comparisons were performed independently at each observation stage within each site. Data were analyzed by analysis of variance (ANOVA) after checking normality and homogeneity of the variances. Mean separation was performed using Tukey’s honestly significant difference (HSD) test at p < 0.05. All statistical analyses were conducted using IBM SPSS Statistics 21 (IBM Corp., Armonk, NY, USA).
Results
Baseline morphological characteristics of fruiting shoots according to the positional category in control trees
Baseline characteristics of fruiting-shoot positional types were evaluated in control trees (Table 1). The flowering period was influenced by the positional origin of fruiting shoots. At both sites, anthesis in Fs1 and Fs2 began approximately ten days earlier than in Fs3, whereas the overall flowering duration was similar among positional types, ranging from 47 to 51 days.
Fruiting-shoot abundance differed between the two sites. In Boeun, Fs1 was the most abundant type, with 417.5 shoots tree-1, followed by Fs3 (193.6 shoots tree-1) and Fs2 (29.1 shoots tree-1). In contrast, Goesan had fewer Fs1 shoots (133.0 shoots tree-1), whereas Fs3 was more abundant than Fs1 (175.5 shoots tree-1). Morphological traits also showed site-specific patterns. In Boeun, Fs1 had significantly more leaves (22.0) and floral clusters (21.2) than Fs3 (17.9 leaves and 16.7 floral clusters), whereas shoot length did not differ significantly among positional types. In Goesan, Fs2 shoots were significantly longer (41.8 cm) than Fs3 shoots (32.1 cm), whereas the number of leaves and floral clusters did not differ significantly among positional types. These results indicate that earlier flowering of Fs1 and Fs2 as compared to Fs3 was common to both sites, whereas fruiting-shoot abundance and morphological characteristics differed between the sites.
Final fruit set and fruit-set ratio per floral cluster
Final fruit set per fruiting shoot and fruit-set ratio A varied according to the PGR treatment, fruiting-shoot position, and site (Tables 2 and 3). Across both sites, GA3-alone treatments generally produced higher fruit set than 4-CPA-containing treatments, although the magnitude and statistical significance of the response differed among fruiting-shoot positions.
Table 2.
Number of floral clusters, fruit set, and fruit-set ratio A by treatment and fruiting-shoot type in Boeun
Fs1–Fs3 denote the three positional types of fruiting shoots (basal bud origin, main axis origin, and lateral branch origin of current-season shoots), respectively. GA1 and GA2 indicate GA3 treatments at 25 and 50 ppm; CPA1 and CPA2 indicate 4-CPA treatments at 10 and 20 ppm; and GCPA1 and GCPA2 indicate combined GA3 + 4-CPA treatments at 20 + 10 and 40 + 20 ppm, respectively. All variables are expressed on a per fruiting shoot basis. Fruit-set ratio A was calculated as the number of fruit sets divided by the number of floral clusters. Values are means ± SD (n = 5 trees per treatment). Within Boeun and for each fruiting shoot type, different letters indicate significant differences among the treatments according to Tukey’s HSD test (p < 0.05).
Table 3.
Number of floral clusters, fruit set, and fruit-set ratio A according to the treatment and fruiting-shoot type in Goesan
Fs1–Fs3 denote the three positional types of fruiting shoots (basal bud origin, main axis origin, and lateral branch origin of current season shoots), respectively. GA1 and GA2 indicate GA3 treatments at 25 and 50 ppm; CPA1 and CPA2 indicate 4-CPA treatments at 10 and 20 ppm; and GCPA1 and GCPA2 indicate combined GA3 + 4-CPA treatments at 20 + 10 and 40 + 20 ppm, respectively. All variables are expressed on a per fruiting shoot basis. Fruit-set ratio A was calculated as the number of fruit sets divided by the number of floral clusters. Values are means ± SD (n = 5 trees per treatment). Within Goesan and for each fruiting shoot type, different letters indicate significant differences among the treatments according to Tukey’s HSD test (p < 0.05).
In Boeun, treatment effects were evident in all three fruiting-shoot types (Table 2). GA3 at 25 ppm (GA1) produced the highest final fruit number in Fs1 and Fs2, with 2.26 and 2.33 fruits shoot-1, respectively. The corresponding fruit-set ratios A were also highest under GA1 in Fs1 and Fs2, reaching 10.5% and 12.2%, respectively. In contrast, Fs3 responded most strongly to GA3 at 50 ppm (GA2), showing the highest fruit number per shoot (2.46 fruits shoot-1) and fruit-set ratio A (13.9%). In general, 4-CPA-alone and GA3 + 4-CPA treatments showed lower fruit set levels than the GA3-alone treatments in Boeun.
In Goesan, treatment responses were less pronounced in Fs1 and Fs2 than in Fs3 (Table 3). In Fs1, GA1 showed the largest values for both fruit number per shoot (2.07 fruits shoot-1) and fruit-set ratio A (11.8%), although its fruit number partially overlapped with those of GA2, CPA1, and CPA2. In Fs2, GA1 showed a similar pattern, with the largest mean values for fruit set (3.27 fruits shoot-1) and fruit-set ratio A (14.2%), with GA2 showing intermediate values. In contrast, Fs3 showed a clear treatment response: GA2 produced the largest values for both fruit number per shoot (5.40 fruits shoot-1) and fruit-set ratio A (33.9%), whereas 4-CPA-containing treatments resulted in markedly lower values.
Overall, the positional response pattern was generally consistent between the two sites. GA1 was more favorable for Fs1 and Fs2, whereas GA2 was more effective for Fs3. However, the statistical strength of this pattern differed by site, with Boeun showing clearer treatment effects across all fruiting-shoot types and Goesan showing the most distinct response in Fs3. These results indicate that the optimal GA3 concentration for improving fruit set may differ according to the positional origin of fruiting shoots.
Fruit set and fruit-set ratio based on total fruiting shoots per tree
Although the composition of fruiting-shoot types differed between Boeun and Goesan, the total number of fruiting shoots did not differ significantly among the treatments within each site (Table 4). In Boeun, the number of fruiting shoots ranged from 620 to 662 shoots tree-1 among the treatments, whereas in Goesan it ranged from 324 to 368 shoots tree-1. Therefore, treatment comparisons within each site were not confounded by differences in total fruiting-shoot abundance among the treatment trees.
Table 4.
Number of fruiting shoots, fruit set, fruit-set ratio B, and yield per tree in response to PGR treatments at two sites
GA1 and GA2 indicate GA3 treatments at 25 and 50 ppm; CPA1 and CPA2 indicate 4-CPA treatments at 10 and 20 ppm; and GCPA1 and GCPA2 indicate combined GA3 + 4-CPA treatments at 20 + 10 and 40 + 20 ppm, respectively. Non marketable fruits refer to fruits affected by cracking, shriveling, or damage caused by diseases or insect pests. Values are means ± SD (n = 5 trees per treatment per site). Within each site, different letters indicate significant differences among the treatments according to Tukey’s HSD test (p < 0.05), and statistical analyses were conducted independently for each site.
At the tree level, GA3 at 25 ppm (GA1) consistently produced the highest total fruit set and fruit-set ratio B at both sites. In Boeun, GA1 increased the fruit-set rate from 358 fruits tree-1 in the control to 1,263 fruits tree-1, representing a 3.53-fold increase. Fruit-set ratio B also increased from 54.7% in the control to 190.7% under GA1. In Goesan, GA1 increased the number of fruits set from 162 to 537 fruits tree-1, corresponding to a 3.31-fold increase, with fruit-set ratio B increasing from 46.5% to 149.3%.
The absolute number of fruits set and total fruiting shoots were higher in Boeun than in Goesan, reflecting the different baseline orchard structures between the two sites. However, the proportional increase in fruit set under GA1 was similar between the sites. These results indicate that GA3 at 25 ppm consistently improved tree-level fruit retention under different orchard conditions, whereas the 4-CPA-alone and GA3 + 4-CPA treatments generally showed lower fruit-set ratios B than the GA3-alone treatment.
Yield per tree and marketable yield
Yield responses were generally consistent with tree-level fruit-set responses (Table 4). At both sites, GA3 at 25 ppm (GA1) produced the highest total yield and marketable yield. In Boeun, GA1 increased the total yield from 4.15 kg tree-1 in the control to 9.44 kg tree-1, representing a 2.27-fold increase. The marketable yield also increased from 2.66 to 6.01 kg tree-1 under GA1. In Goesan, GA1 increased the total yield from 2.02 to 5.14 kg tree-1, corresponding to a 2.54-fold increase, with the marketable yield increased from 1.75 to 4.44 kg tree-1.
Although the absolute yield was higher in Boeun than in Goesan, the relative increase in the total yield under GA1 was comparable between the two sites. This pattern indicates that GA3 at 25 ppm consistently improved the yield by increasing fruit retention, despite differences in the baseline orchard structure and fruiting-shoot abundance. The non-marketable yield showed a site-specific response. In Boeun, the non-marketable yield increased after the high-yielding GA3 treatments, whereas in Goesan, no significant differences in the non-marketable yield were detected among the treatments.
Fruit and stone characteristics
Fruit and stone characteristics varied by treatment and site (Table 5). In Boeun, fruit length was greatest in the control and CPA1 treatments, whereas the GA3 and GA3 + 4-CPA treatments produced relatively short fruits. Fruit width was highest in CPA1 and CPA2, while fruits from GA3- and GA3 + 4-CPA-treated trees were generally narrower. Fruit firmness and SSC did not differ significantly among the treatments.
Table 5.
Fruit and stone characteristics of marketable jujube fruits by treatment and site
Fruit and stone morphological indices were calculated as the ratios of the length to width (length/width). GA1 and GA2 indicate GA3 treatments at 25 and 50 ppm; CPA1 and CPA2 indicate 4-CPA treatments at 10 and 20 ppm; and GCPA1 and GCPA2 indicate combined GA3 + 4-CPA treatments at 20 + 10 and 40 + 20 ppm, respectively. Quality and morphological measurements were performed only on marketable fruits. Values are means ± SD (n = 5 trees per treatment per site), with 30 fruits per tree treated as subsamples and summarized as tree means before the analysis. Within each site, different letters indicate significant differences among the treatments according to Tukey’s honest significant difference (HSD) test (p < 0.05).
In Goesan, fruit length and width were highest in CPA1, whereas GCPA2 produced the smallest fruits. The fruit morphological index, fruit firmness, and SSC were not significantly affected by the treatments. Stone traits also varied according to the treatment. In Boeun, stone size and weight were generally greater in the control or CPA2 treatments, whereas GA2 produced the lowest stone weight and smaller stone dimensions. In Goesan, stone length and width were higher in the control or CPA2 treatments than in the GA3 + 4-CPA treatments, and stone weight was lowest under GA2. Overall, treatment effects on fruit and stone traits were less consistent between the two sites than treatment effects on fruit set and yield.
Phytotoxic symptoms
Visual phytotoxic symptoms were not observed at DAT14-1 or DAT14-2 but were detected at DAT14-3 in the 4-CPA-containing treatments at both sites. No visible injuries were observed in the control or GA3-only treatments throughout the observation period (Fig. 2K and 2L; Supplementary Table S2). The main symptoms included leaf tip burn, inward leaf curling, and browning or necrosis of calyx tissues, which were accompanied by reduced flower opening. At DAT14-3, visual injury scores ranged from 1 to 3 in the 4-CPA-containing treatments, whereas the control and GA3-only treatments had scores of 0 at both sites. The highest median injury scores were observed in CPA2 and GCPA2. Fruiting-shoot and pedicel morphological traits after PGR treatment are summarized in Supplementary Table S4. PGR effects on these traits were site- and shoot-type dependent, whereas the pedicel length was generally longer in PGR-treated trees than in the control.

Fig. 2.
Experimental overview and representative treatment responses after PGR application in jujube. (A, B) Experimental sites in Boeun and Goesan (May 15, 2024): (C–F) Basal fruiting shoots (Fs1) in Boeun under the Control, GA3 25 ppm (GA1), 4-CPA 10 ppm (CPA1), and GA3 + 4-CPA (20 + 10 ppm, GCPA1) treatments (July 21, 2024). (G–J) Lateral fruiting shoots (Fs3) in Goesan under the same treatments (August 7, 2024). (K) Leaf tip burn and curling. (L) Calyx scorching and inhibited flower opening due to floral deformation (July 21, 2024).
Discussion
This study evaluated fruit-set responses to GA3, 4-CPA, and combined GA3 + 4-CPA treatments according to the positional origin of fruiting shoots in open-field jujube orchards. Previous studies examined PGR effects on jujube fruit set at the flower, canopy, or whole-tree level (Oh et al. 1990; CARES 2017b; Mishra et al. 2020; Fan et al. 2022), and other studies characterized differences in branch or canopy traits, including nutrient compositions and leaf responses (Song et al. 2021; Bao et al. 2022). However, relatively little information is available on how fruiting shoots with different developmental origins respond to a PGR application. By classifying fruiting shoots into Fs1, Fs2, and Fs3 and combining shoot-level and tree-level fruit-set indices, this study provides a framework for evaluating position-specific PGR responses in jujube.
Jujube fruiting shoots are deciduous annual organs that develop on current-season shoots and bear flowers and fruits within the same growing season (Liu et al. 2006; Yao 2013; Liu et al. 2020; Song et al. 2021; Lan et al. 2025). In this study, Fs1 and Fs2 began flowering earlier than Fs3 at both sites, indicating that fruiting-shoot positional origin was closely associated with flowering phenology. This pattern is consistent with the prolonged and asynchronous flowering habit of jujube reported in previous studies (Sun et al. 2009; Hernández et al. 2015; Nam et al. 2015; Yao et al. 2015; Oh 2019). The overall flowering duration was similar among fruiting-shoot types, suggesting that positional origin mainly affect flowering onset rather than the total flowering duration. This difference in reproductive phenology may influence the developmental stage at which each fruiting-shoot type responds to a PGR application.
The difference in fruiting-shoot abundance between Boeun and Goesan should be interpreted as a baseline orchard-structure difference rather than a treatment effect. In Boeun, the trees were older, had more primary scaffold branches, and contained a greater amount of older perennial wood. These conditions likely increased the number of basal bud-origin shoots and resulted in a higher abundance of Fs1. In contrast, the younger Goesan trees had fewer scaffold branches and relatively more current-season shoot growth, which may have reduced the relative abundance of Fs1 and increased the proportion of lateral branch-origin shoots such as Fs3. Similar effects of tree age, the scaffold structure, and pruning on jujube growth and fruiting have also been reported previously (CARES 2017a; Lee et al. 2017; Kim et al. 2021). Therefore, the contrasting fruiting-shoot compositions between the sites support the need to consider the orchard structure when evaluating fruit set and PGR responses in jujube.
The use of two complementary fruit-set indices helped distinguish positional responses from whole-tree responses. Fruit set relative to floral clusters was used to evaluate shoot-level responses among Fs1, Fs2, and Fs3. This approach is useful because jujube produces numerous flowers, and the flower number can vary widely among floral clusters and fruiting shoots (Liu et al. 2006; Yao et al. 2015; Zou et al. 2017; Meng et al. 2020). In contrast, fruit set relative to total fruiting shoots per tree was used to validate whether shoot-level responses were reflected at the tree level. Similar approaches are useful when total flower counts are impractical or when fruiting potential differs among trees due to differences in pruning intensity, branch density, or canopy structure characteristics (Alburquerque et al. 2004; Majumder et al. 2013; Lee and Chang 2019; Yoo et al. 2022). Thus, fruit sets A and B allowed treatment effects to be compared while accounting for between-site differences in the fruiting-shoot composition, as the floral cluster number per fruiting shoot and total fruiting-shoot number per tree outcomes did not differ significantly among the treatments within each site.
The application of GA3 during flowering consistently improved fruit set and yield, particularly at 25 ppm. At both sites, GA1 produced the highest tree-level fruit set, fruit-set ratio B, total yield, and marketable yield. These findings are consistent with previous reports that gibberellins enhance fruit set in jujube by improving pollen germination, pollen-tube growth, and fruit-set-related hormonal regulation (Ayimaiti et al. 2020; Fan et al. 2022). The positive response to GA3 may also be associated with the parthenocarpic tendency of jujube given that gibberellins are known to participate in ovary growth and parthenocarpic fruit development in several fruit crops (Talon et al. 1992; Cheng et al. 2013; Jung and Hur 2020). However, the objective of the present study was not to induce seedless fruit in jujube. Rather, the aim was to improve fruit retention and stabilize fruit set during the flowering period in a species that already exhibits parthenocarpic tendencies (Ackerman 1961; Liu et al. 2008; Yao et al. 2015).
The response to GA3 differed according to the fruiting-shoot position. Fs1 and Fs2 generally responded more favorably to GA3 at 25 ppm, whereas Fs3 showed a stronger response to GA3 at 50 ppm. Because jujube fruiting shoots bear flowers and fruit within the same growing season and have a prolonged flowering period (Liu et al. 2006; Yao et al. 2015), fruitlets on earlier-flowering Fs1 and Fs2 were likely already undergoing early enlargement when later-developing Fs3 shoots reached the active flowering or early fruit-retention stages. In woody fruit trees, the fruit distribution and crop load can influence dry matter partitioning within branch units, supporting the possibility of localized source–sink competition (Marsal et al. 2003). Thus, flowers on Fs3 may have competed with previously retained fruitlets on Fs1 and Fs2, as well as newly opened flowers on the terminal portions of earlier-positioned shoots, for assimilates and endogenous growth-regulatory signals. Under these conditions, Fs3 may have required a stronger exogenous GA3 stimulus to maintain flower or young-fruit retention, consistent with the role of GA3 in jujube fruit-set regulation (Fan et al. 2022). However, endogenous GA levels, the carbohydrate distribution, sink strength, and pollen-tube growth were not directly measured; therefore, this interpretation should be regarded as a physiological hypothesis requiring further validation.
The temporal pattern of fruit set also requires careful interpretation. The number of fruits did not increase cumulatively after successive applications but declined during the observation period. This decline likely reflects the combined effects of asynchronous flowering, fruitlet abscission, and unfavorable weather conditions during the flowering and early fruit-retention period. In Korea, the flowering period of jujube overlaps with the East Asian monsoon season, when frequent rainfall and reduced solar radiation can contribute to physiological fruit drop (KMA 2022; KREI 2023; KREI 2024). The daily weather data summarized in Supplementary Table S1 provide the site-specific meteorological context for interpreting fruitlet abscission during flowering and early fruit retention under open-field conditions (KMA 2024). Although this study was conducted in a single growing season, the two-site design partially mitigated this limitation by including orchards that differed in terms of their tree age, planting distance, scaffold structure, and fruiting-shoot composition. Nevertheless, because jujube flowering and early fruit retention overlap with the East Asian monsoon season, the response to GA3 can vary depending on the seasonal rainfall distribution and the onset of the monsoon. Therefore, the superior response to 25 ppm GA3 should be interpreted as the optimal response under the 2024 field conditions tested here. Multi-year trials, preferably encompassing at least two or three growing seasons that include early- and late-monsoon years, are needed to determine whether this concentration remains optimal across a broader range of weather conditions.
The comparison between the GA3-alone treatments and the combined GA3 + 4-CPA treatments requires cautious interpretation. In this study, commercially available PGR products were selected to improve practical applicability under field conditions. However, the GA3 concentrations in the combined formulations were 20 and 40 ppm, whereas those in the GA3-alone treatments were 25 and 50 ppm. Therefore, the combined treatments were not factorial combinations designed to isolate the additive or synergistic effect of 4-CPA under identical GA3 concentrations. Rather, they should be interpreted as evaluations of commercially available premixed formulations under open-field orchard conditions, not as tests of chemical synergy.
Fruit and stone characteristics were less consistently affected by the PGR treatment than fruit set and yield. Fruit size tended to be larger in the control or 4-CPA-alone treatments, particularly CPA1, whereas high-fruit-set GA3 treatments generally produced smaller fruits. This pattern suggests that differences in fruit size were more closely associated with the crop load than with a direct negative effect of GA3 on fruit growth. In fruit crops, reducing the crop load by thinning can increase the fruit size or the proportion of fruit in larger marketable size classes (Schupp et al. 2008; Wang et al. 2023). Therefore, the relatively small fruits observed in the high-yielding GA3 treatments may reflect assimilate distribution among a larger number of retained fruits. Fruit firmness and SSC were not significantly affected by PGR treatments at either site, indicating that the increased fruit retention under GA3 treatment did not substantially alter these quality traits. From a commercial perspective, GA3 at 25 ppm may be advantageous for yield stabilization, but its practical value should be evaluated together with the fruit size distribution and marketable grade composition.
Stone traits also varied among the treatments, but the patterns were not consistent between the sites. Gibberellins are commonly used to induce seedlessness or alter seed development in other crops, such as grape (Talon et al. 1992; Cheng et al. 2013; Jung and Hur 2020). Because jujube has parthenocarpic tendencies (Ackerman 1961; Liu et al. 2008; Yao et al. 2015), we examined whether PGR treatments were associated with changes in the stone size. Although some treatments showed smaller stone dimensions or lower stone weights, these reductions generally corresponded to a reduced overall fruit size rather than providing clear evidence of direct PGR-induced stone shrinkage. Previous studies demonstrated that stone traits are closely associated with the overall fruit morphology in jujube (Oh 2019; Oh et al. 2025). Therefore, the present results do not support a strong conclusion that GA3 or 4-CPA directly hindered stone development.
PGR treatments produced limited and site-dependent effects on fruiting-shoot vegetative traits. Although gibberellins have been reported to promote elongation growth in several horticultural crops (Othman et al. 2021; Zhang et al. 2023), the fruiting-shoot length and number of leaves did not show consistent treatment responses across both sites or across PGR classes in the present study. Therefore, the improvement in fruit set under the GA3 treatment should not be interpreted primarily as a simple consequence of enhanced vegetative growth. The pedicel length was generally greater in PGR-treated trees than in the controls at both sites; however, because longer pedicels were observed not only in GA3 treatments but also in the 4-CPA-containing treatments, the physiological significance of pedicel elongation in jujube remains unclear.
In addition to their lower fruit-set responses, the 4-CPA-containing treatments showed visible phytotoxic symptoms, unlike the GA3-only treatments. The main symptoms included leaf tip burn, inward leaf curling, calyx browning or necrosis, and reduced flower opening. In the present study, no visible injuries were detected after the first or second PGR application, whereas symptoms appeared 14 days after the third application of the 4-CPA-containing treatments (Supplementary Table S2). These symptoms were most evident in terminal or young expanding tissues of fruiting shoots, suggesting that repeated exposure to 4-CPA-containing sprays may have disturbed the growth-regulatory balance required for normal vegetative and floral development during flowering. This response is broadly consistent with previous reports which found that excessive applications of 4-CPA can adversely affect plant growth and fruit quality outcomes (Gemici et al. 2006). However, because 4-CPA residue, the degradation rate, and tissue-level accumulation were not measured, we cannot conclude that the delayed symptoms resulted from chemical accumulation in leaf tips. Future studies should include residue analysis and quantitative measurements of injured leaves, flowers, and fruitlets to clarify the mechanism of 4-CPA-induced phytotoxicity in jujube.
Overall, GA3 at 25 ppm was the most effective treatment for improving the tree-level fruit set and yield in open-field jujube orchards. The response was consistent at both Boeun and Goesan, despite differences in the tree age, scaffold structure, fruiting-shoot composition, and baseline yield potential. Nevertheless, the position-specific response pattern, particularly the stronger Fs3 response to GA3 at 50 ppm, indicates that fruiting-shoot developmental origin and the phenological sequence can influence the optimal PGR response. This finding supports the value of evaluating fruit set not only at the whole-tree level but also according to the fruiting-shoot position.
Several limitations should be considered when interpreting the results. First, as discussed above, the experiment was conducted in a single growing season; therefore, the stability of the 25 ppm GA3 response should be validated through multi-year trials under contrasting monsoon patterns. Second, direct physiological measurements, such as endogenous hormone levels, carbohydrate distribution, pollen-tube growth, and source–sink relationships, were not conducted. Third, the combined GA3 + 4-CPA treatments were based on commercial premixed formulations rather than factorial combinations with identical GA3 concentrations. Fourth, phytotoxic symptoms were evaluated using ordinal visual injury scores; however, future studies should include more rigorous quantitative measurements, such as the proportion of injured leaves, flowers, or fruitlets, to allow a statistical comparison among 4-CPA concentrations. Finally, the long-term effects of increased fruit retention should be examined, including possible impacts on reserve accumulation, fruiting-shoot formation in the following year, tree vigor, and alternate bearing. Considering the labor requirements and potential for tree injuries associated with girdling, which is commonly used by growers to overcome poor fruit set, the application of GA3 may represent a practical alternative for stabilizing jujube production. However, the optimal concentration and timing should be further refined for different cultivars, orchard structures, and production systems, including rain-shelter and greenhouse cultivation (Acheampong et al. 2017; El-Masri et al. 2018; Senthilkumar et al. 2018).
Supplementary Material
Supplementary materials are available at Horticultural Science and Technology website (https://www.hst-j.org).
- HORT_20260020_Fig_S1.docx
Supplementary Fig. S1. Schematic representation of the randomized complete block design (RCBD) used at both the Boeun and Goesan sites.
- HORT_20260020_Table_S1.docx
Supplementary Table S1. Daily precipitation and air temperature during the flowering and PGR application period in Boeun and Goesan in 2024.
- HORT_20260020_Table_S2.docx
Supplementary Table S2. Visual phytotoxic injury scores recorded at DAT14-3 in Boeun and Goesan in 2024.
- HORT_20260020_Table_S3.docx
Supplementary Table S3. Temporal changes in fruit set per fruiting shoot by treatment, fruiting-shoot type, and site.
- HORT_20260020_Table_S4.docx
Supplementary Table S4. Fruiting-shoot lengths, number of leaves, and pedicel length by treatment, fruiting-shoot type, and site.
