

Introduction
Materials and Methods
Experimental materials
Characteristics of ‘Hongro’ and ‘Greenball’
Protein extraction and two-dimensional electrophoresis (2-DE)
MALDI-TOF/TOF MS
Results
Characteristics of ‘Hongro’ and ‘Greenball’
Differences in proteomes expressed in the peel specimens of ‘Hongro’ and ‘Greenball’
Identification and functional analysis of proteomes
Discussion
‘Hongro’
‘Greenball’
Introduction
Apples (Malus domestica Borkh.) are widely cultivated worldwide with more than 10,000 cultivars differing in many aspects, including fruit characteristics, yield, and stress tolerance (Janick et al. 1996; Christopher et al. 2009). Proteomics, the study of the proteome expressed within organisms, can be applied to various practical agricultural factors, such as breeding, stress tolerance, growth, storage, and quality. Annurca (Malus domestica Borkh. cv. Annurca), an apple cultivar grown in southern Italy, is known for its long shelf life and crunchy texture. An analysis of the flesh proteome of three Annurca apple lines revealed that 203 proteins were commonly expressed, 39 of which belonged to functional categories such as metabolism, energy, stress, ripening, and allergens (Guarino et al. 2007). Apples are climacteric fruits, and as they mature various physiological changes occur, such as rapid increases in respiration and ethylene production, decreases in firmness and acidity, and increases in volatile compounds (Mattheis 1991; Song and Bangerth 1996; Peng and Lu 2007). Among apple cultivars, those with high hardness and storage properties are known to have low expression rates of proteins related to ethylene biosynthesis and high expression rates of proteins related to the maintenance of the cytoskeleton compared to those with low hardness and storage properties (Marondedze and Thomas 2012). Apples have been associated with several health benefits, including a reduced risk of chronic diseases such as heart disease, stroke, and certain cancers (Van‘t Veer et al. 2000; Boyer and Liu 2004). However, due to the presence of allergens, apples may induce allergic reactions in susceptible individuals. The major allergens in apples include Mal d1, Mal d2, Mal d3, and Mal d4 (Herndl et al. 2007). Recently, a study of the large-scale proteome in the peel of ‘Hongro’, an apple cultivar bred and cultivated in Korea, reported that many proteins related to carbohydrate and protein metabolism, defense and stress responses, antioxidants, secondary metabolism, lipid, transport, the cytoskeleton, nucleotides, and allergens are expressed (Kim et al. 2020). In addition, proteins related to ethylene and secondary metabolite biosynthesis, photosynthesis, defense and stress responses, antioxidants, and allergens were expressed at a higher rate in the peel than in the flesh, and only a few proteins related to carbohydrate metabolism were expressed at a higher rate in the flesh than in the peel of ‘Hongro’ (Chung et al. 2021).
Apples are the most widely cultivated fruit crop in Korea and rank second among all fruits in terms of cultivation area, production volume, and production amount (KOSIS 2022). In 2018, the cultivation area rankings in Korea according to apple cultivar were ‘Fuji’ at 76.0%, ‘Hongro’ at 11.8%, ‘Yoko’ at 0.5%, and ‘Tsugaru’ at 0.3% (KOSIS 2020). Among these cultivars, the ‘Hongro’ cultivar used in this study was bred domestically. The optimal harvest time for this cultivar is early to mid-September. It has weak resistance to pests and diseases and a deep red skin color. The shelf life at room temperature is about 30 days. Meanwhile, ‘Greenball’ was also bred in Korea and is a niche market cultivar with a much smaller cultivation area compared to ‘Hongro’. The optimal harvest time for this cultivar is early September, similar to ‘Hongro’. The peel is greenish-yellow and the shelf life is very short, about a week at room temperature. This study compared the proteomes expressed in the peel of the Korean apple cultivars ‘Hongro’ and ‘Greenball’, which have distinctly different characteristics, such as color and storability. Using two-dimensional electrophoresis, proteins with different expression rates were identified by a MALDI-TOF/TOF MS analysis. The aim of this study is to identify protein profiles with varying expression rates in apple cultivars with different mature skin colors and to determine the physiological and biochemical roles associated with the characteristics of each cultivar. These results are expected to serve as indicators in a wide range of future research related to the breeding, storage, physiological characteristics, and stress tolerance of apple cultivars.
Materials and Methods
Experimental materials
On September 3, 2021, nine ‘Hongro’ (deep red) and ‘Greenball’ (greenish yellow) at harvest time were sampled in three repetitions from three trees (three samples per tree) at the Apple Utilization Research Institute of Gyeongnam Agricultural Research and Extension Services. On the day of harvest, the apples were washed with tap water and then peeled, ensuring that the peel thickness was kept to less than 1 mm. The peel was frozen, ground in liquid nitrogen, and stored at ‒75°C until protein extraction.
Characteristics of ‘Hongro’ and ‘Greenball’
To determine the fruit characteristics of each cultivar, the color, browning degree, hardness, sugar content, and acidity were measured. Chromaticity was measured using a portable colorimeter (Spectrophotometer CM-700d, Minolta Co., Tokyo, Japan), and the L (lightness) value, A value (+: redness, ‒: greenness), and B value (+: yellowness, ‒: blueness) for each cultivar were measured six times after standardization on a white board (L = 99.54, a = +0.05, b = ‒0.32). The degree of browning was assessed according to the L value, A value, and B value of the cut surface at room temperature 30 minutes after the apples were cut. Hardness was measured using a device capable of measuring this physical property (Texture Analyzer TMS-Touch, FTC, Virginia, USA) by laying the apples on their left and right sides, with the results for both sides of each apple added and multiplied by 9.807 (Newton index) to express the hardness sum (N, Newton). Acidity was measured using an acidity meter (G20 compact Titrator, METTLER TOLEDO, Schwerzenbach, Switzerland) by converting the appropriate value of 0.1 N NaOH to malic acid until the pH level reached 8.3%, with the titration amount indicating the amount of 0.1N NaOH added.
Protein extraction and two-dimensional electrophoresis (2-DE)
Proteins from the peel specimens of ‘Hongro’ and ‘Greenball’ were extracted using the phenol method (Hurkman and Tanaka 1986; Kim et al. 2020), dissolved in lysis buffer, and quantified using Coomassie Brilliant blue G (CBBG)-250 (Bradford 1976). A total amount of 400 µg of protein was separated by isoelectronic focusing (IEF) electrophoresis and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (Kim et al. 2020). The SDS-PAGE gel was then stained with CBBG-250 (Bio-Rad, CA) (Matsui et al. 1999). Two-dimensional electrophoresis (2-DE) was conducted six times using protein samples extracted twice each from three replicate samples. Gel images were scanned with a GS-800 calibrated densitometer (Bio-Rad, Hercules, CA, USA) and then analyzed using Quantity One version 4.6.9 software (Bio-Rad, Hercules, CA, USA). For protein spots, the abundance value (AV) was measured by comparing the adjusted volume with the gel background. Protein spots that showed more than a two-fold difference in the abundance value between the two apple cultivars were selected at the 5% significance level using Student's t-test. The selected spots were assigned unique numbers in order from a low pI value and high molecular weight to a high pI value and low molecular weight and were used for the protein analysis.
MALDI-TOF/TOF MS
Protein spots were decolorized with acetonitrile and ammonium bicarbonate, reduced, alkylated, and then digested with trypsin (Kim et al. 2020). Peptides were extracted with an extraction solution, frozen, dried, and then dissolved with the sample solution and mixed with a matrix. The peptide and matrix mixture was loaded onto a dedicated auto flex max MALDI-TOF/TOF (Bruker Daltonics, Bremen, Germany) plate and subjected to MS and MS/MS analyses. The TOF-TOF MS analysis was conducted using 500 laser shots with external calibration under the following conditions: a Smartbeam Ⅱ laser (355 nm), 2 kHz in MS, 200 Hz in MS/MS, positive ion mode and reflectron mode. The mass peak was processed using Flex Analysis (ver 3.4). MS and MS/MS spectra data were analyzed using the UniProt database (20210916 version) at a mass tolerance level of 100 ppm. Single miss cleaved trypsin digestion, oxidation of methionine, and carbamidomethylation of cysteine were allowed for MS/MS spectrum searches in the database. After the MS analysis, MS/MS was performed using laser-induced decomposition, and a search was conducted for each peptide ion score using a significance threshold of p = 0.05 in the Mascot protein database.
Results
Characteristics of ‘Hongro’ and ‘Greenball’
The peel of ‘Hongro’ used in the experiment was deep red, while the peel of ‘Greenball’ was greenish yellow (Fig. 1.Fig. 1). Analyzing the characteristics of each cultivar, the redness of the peel was significantly higher in ‘Hongro’ (30.88) than in ‘Greenball’ (‒9.11), and the yellowness was significantly higher in ‘Greenball’ (40.67) than in ‘Hongro’ (18.87) (Table 1.Table 1). The redness and browning degree of the flesh were higher in ‘Hongro’ than in ‘Greenball’. Additionally, the sugar content was higher in ‘Hongro’, while the acidity level was higher in ‘Greenball’.

Table 1.
Comparison of the characteristics of ‘Greenball’ and ‘Hongro’ apple cultivars
| Variety | Chromaticity of peel | Hardness | Sum of hardness (N) |
Chromaticity of flesh | Degree of browning |
Soluble solids (ºBrix) | Acidity (%) | ||||
| A*y | B*x | Left | Right | A*y | B*x | A*y | B*x | ||||
| Greenball |
‒9.11 (±0.22) |
40.67 (±0.70) |
3.95 (±0.15) |
3.90 (±0.27) |
38.49 (±1.71) |
‒1.62 (±0.35) |
17.79 (±0.74) |
‒0.18 (±0.14) |
22.49 (±0.66) |
10.5 (±0.54) |
0.29 (±0.01) |
| Hongro |
30.88 (±0.62) |
18.87 (±0.33) |
4.43 (±0.19) |
4.33 (±0.26) |
42.95 (±2.11) |
‒0.57 (±0.13) |
17.55 (±0.46) |
1.12 (±0.35) |
21.92 (±1.12) |
13.1 (±0.31) |
0.16 (±0.01) |
| p valuez | 0.000 | 0.000 | 0.103 | 0.321 | 0.166 | 0.029 | 0.810 | 0.011 | 0.695 | 0.003 | 0.000 |
Differences in proteomes expressed in the peel specimens of ‘Hongro’ and ‘Greenball’
To analyze the proteomes expressed in ‘Hongro’ and ‘Greenball’ comparatively, 2-DE was utilized, and protein spots with expression rates that differed by more than two-fold were selected. There was no significant difference in the protein spots expressed in the pI range of 4–5 (results not shown), but in the pI range of 5–8, 29 protein spots showed more than a two-fold difference in the expression rate (Figs. 2 and 3). In ‘Hongro’, 17 proteins, consisting of spots 1, 2, 3, 6, 7, 8, 9, 13, 17, 19, 20, 21, 22, 23, 24, 25, and 28, were more highly expressed. In ‘Greenball’, 12 proteins consisting of spots 4, 5, 10, 11, 12, 14, 15, 16, 18, 26, 27, and 29 were more highly expressed.
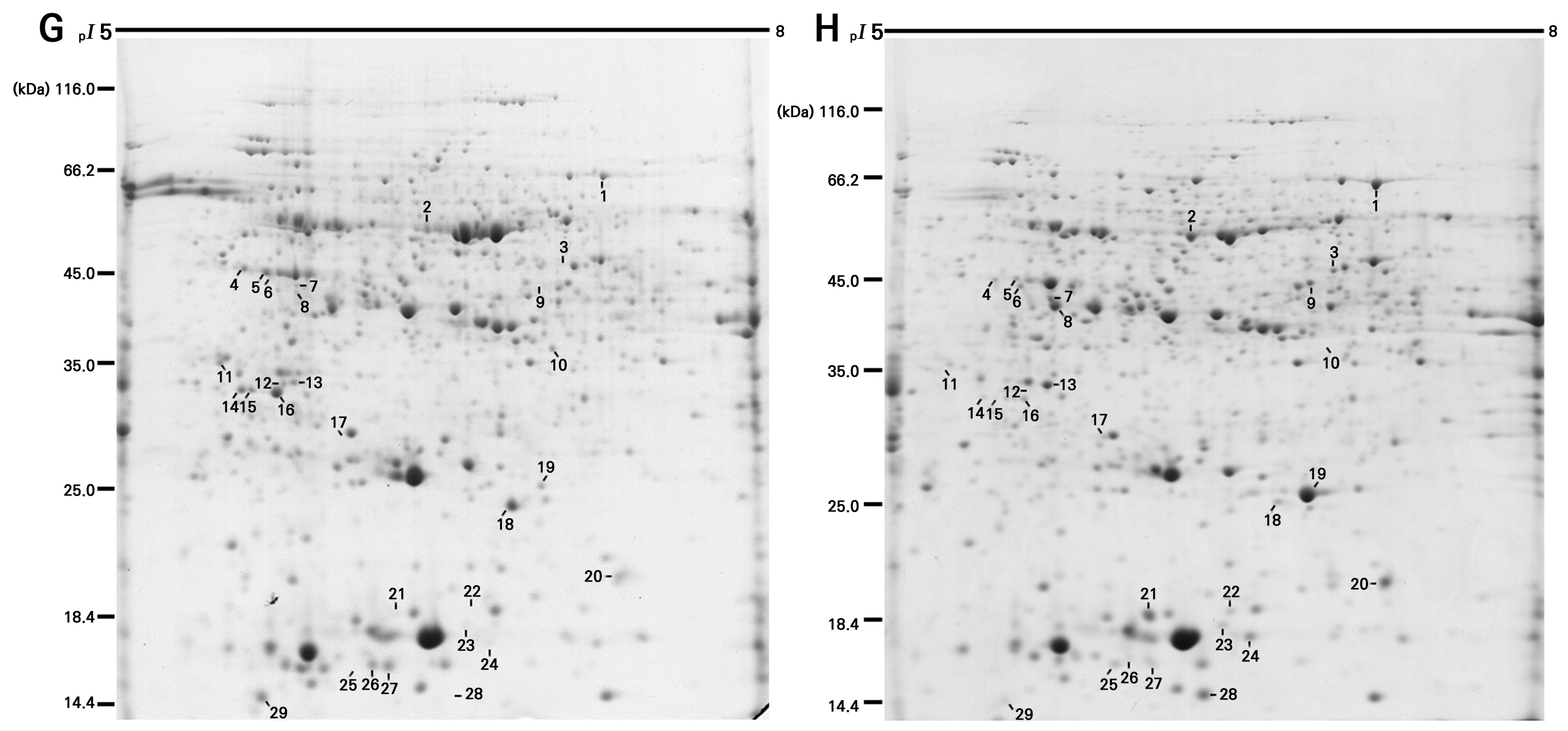
Fig. 2.
Comparison of representative 2-DE gel images of the peel of ‘Greenball (G)’ and ‘Hongro (H)’: in total, an amount of 400 ug of proteins was separated on IPG strips (17 cm, pH5-8) and 11.5% polyacrylamide SDS-PAGE. The gels were stained with colloidal CBB and differently expressed protein spots were measured by Quantity One version 4.6.9 software (Bio-Rad, Hercules, CA, USA).

Identification and functional analysis of proteomes
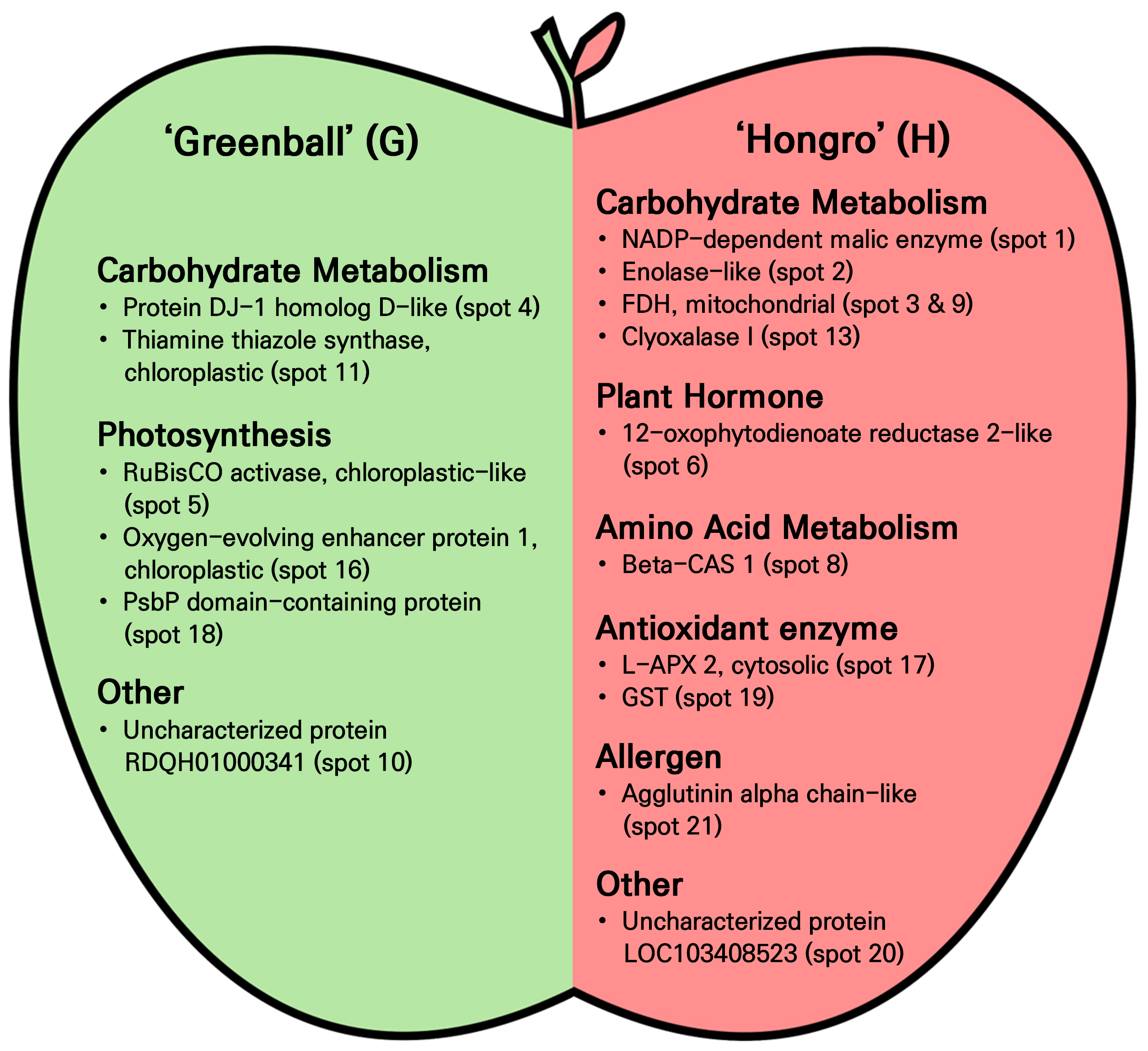
Twenty-nine protein spots with expression rates that differed by more than two-fold in ‘Hongro’ and ‘Greenball’ were preprocessed by coring and were subsequently analyzed by MALDI-TOF/TOF. As a result, eleven spots (spots 1, 2, 3, 6, 8, 9, 13, 17, 19, 20, and 21) highly expressed in ‘Hongro’ and six spots (spots 4, 5, 10, 11, 16, and 18) highly expressed in ‘Greenball’ were identified as proteins with known physiological functions (Table 2). In ‘Hongro’, proteins related to carbohydrate metabolism (spots 1, 2, 3, 9, and 13), plant hormones (spot 6), amino acid metabolism (spot 8), antioxidant enzymes (spots 17 and 19), allergens (spot 21), and other factors (spot 20) were identified, while in ‘Greenball’, proteins related to carbohydrate metabolism (spots 4 and 11), photosynthesis (spots 5, 016 and 18) , and other factors (spot 10) were identified (Fig. 4). As shown below, the proteins identified in each variety were organized by function to determine their roles in relation to the characteristics of ‘Hongro’ and ‘Greenball’.
Table 2.
Identification of differently expressed proteins in the peels of ‘Greenball’ and ‘Hongro’ apple cultivars by MALDI-TOF/TOF MS
|
Spot Number | Protein name | Organism |
Accession number | Abundance value |
Mww/pIv (Theoretical) |
Mww/pIv (Observed) | Scoreu | |||
| G | H | G/H ratio | p valuex | |||||||
| 1 | NADP+-dependent malic enzyme | Malus domestica | gi|658004919 | 327.8 (±50.2) | 995.1 (±40.3) | 1.0 : 3.0 | 0 | 64920/5.72 | 64.3/7.3 | 264 |
| 2 | Enolase-like | Malus domestica | gi|657998659 | 360.6 (±17.0) | 981.5 (±35.6) | 1.0 : 2.7 | 0 | 47981/5.92 | 53.9/6.4 | 392 |
| 3 | Formate dehydrogenase, mitochondrial | Malus domestica | gi|658012400 | 34.5 (±3.6) | 255.3 (±8.0) | 1.0 : 7.4 | 0 | 42384/6.50 | 46.6/7.1 | 139 |
| 4z | Protein DJ-1 homolog D-like | Malus domestica | gi|657978922 | 263.3 (±18.7) | 123.4 (±2.7) | 2.1 : 1.0 | 0.008 | 42459/5.14 | 45.4/5.5 | 68 |
| 5 | RuBisCO activase, chloroplastic-like | Malus domestica | gi|658028348 | 564.7 (±38.1) | 170.2 (±4.8) | 3.3 : 1.0 | 0.003 | 52088/6.49 | 45.1/5.5 | 246 |
| 6z | 12-Oxophytodienoate reductase 2-like | Malus domestica | gi|658058901 | 76.1 (±8.4) | 234.0 (±7.7) | 1.0 : 3.1 | 0 | 41456/5.21 | 44.3/5.5 | 119 |
| 7 | Unidentified | - | - | 89.6 (±5.9) | 276.8 (±14.9) | 1.0 : 3.1 | 0 | - | 43.2/5.8 | - |
| 8z | β-CAS 1 | Malus domestica | gi|93359259 | 72.6 (±4.8) | 1080.9 (±31.2) | 1.0 : 14.9 | 0 | 40081/7.60 | 42.4/5.7 | 133 |
| 9 | Formate dehydrogenase, mitochondrial | Malus domestica | gi|658012400 | 31.9 (±3.8) | 294.3 (±6.6) | 1.0 : 9.2 | 0 | 42384/6.50 | 44.2/7.0 | 443 |
| 10 | Uncharacterized protein RDQH01000341 | Malus domestica | Uniprot A0A498HPZ2 | 250.1 (±19.2) | 13.9 (±0.9) | 18.0 : 1.0 | 0.002 | 65304/7.11 | 37.1/7.1 | 92 |
| 11z | Thiamine thiazole synthase, chloroplastic | Malus domestica | gi|657954088 | 204.6 (±15.7) | 18.5 (±2.5) | 11.1 : 1.0 | 0.002 | 39915/5.71 | 35.3/5.4 | 175 |
| 12 | Unidentified | - | - | 208.5 (±26.9) | 52.5 (±5.6) | 4.0 : 1.0 | 0.016 | - | 33.3/5.7 | - |
| 13 | Glyoxalase Ⅰ | Gossypium hirsutum | gi|211906514 | 263.1 (±35.8) | 897.8 (±33.2) | 1.0 : 3.4 | 0 | 33609/5.69 | 33.9/5.7 | 119 |
| 14 | Unidentified | - | - | 358.7 (±18.8) | 71.4 (±3.2) | 5.0 : 1.0 | 0.001 | - | 32.9/5.4 | - |
| 15 | Unidentified | - | - | 336.4 (±31.0) | 46.4 (±3.7) | 7.3 : 1.0 | 0.004 | - | 32.9/5.4 | - |
| 16z | Oxygen-evolving enhancer protein 1, chloroplastic | Malus domestica | gi|658006067 | 1075.3 (±48.8) | 152.8 (±6.7) | 7.0 : 1.0 | 0.001 | 35263/5.76 | 32.8/5.6 | 87 |
| 17z | AP 2, cytosolic | Malus domestica | gi|658029900 | 25.7 (±4.9) | 138.8 (±6.7) | 1.0 : 5.4 | 0 | 27887/5.39 | 29.7/6.0 | 86 |
| 18 | PPD-containing protein | Malus domestica | Uniprot A0A498J6A3 | 1056.5 (±45.7) | 216.9 (±4.8) | 4.9 : 1.0 | 0.001 | 28842/8.67 | 24.9/6.9 | 77 |
| 19 | GST-like | Malus domestica | gi|658055911 | 156.8 (±2.6) | 3097.1 (±100.4) | 1.0 : 19.8 | 0 | 23979/6.17 | 24.2/6.8 | 168 |
| 20 | Uncharacterized protein LOC103408523 | Malus domestica | gi|658020415 | 100.9 (±12.3) | 933.4 (±15.8) | 1.0 : 9.3 | 0 | 18560/6.75 | 20.8/7.30 | 145 |
| 21z | Agglutinin alpha chain-like (2020) | Malus domestica | gi|658026136 | 95.1 (±7.3) | 1142.6 (±38.4) | 1.0 : 12.0 | 0 | 20818/9.43 | 18.6/6.3 | 68 |
| 22 | Unidentified | - | - | 32.2 (±2.7) | 157.9 (±3.9) | 1.0 : 4.9 | 0 | - | 18.8/6.7 | - |
| 23 | Unidentified | - | - | 56.9 (±5.9) | 164.7 (±6.7) | 1.0 : 2.9 | 0 | - | 17.6/6.6 | - |
| 24 | Unidentified | - | - | 82.6 (±9.3) | 617.7 (±6.7) | 1.0 : 7.5 | 0 | - | 17.2/6.7 | - |
| 25 | Unidentified | - | - | 50.9 (±10.7) | 283.9 (±12.6) | 1.0 : 5.6 | 0 | - | 16.2/6.1 | - |
| 26 | Unidentified | - | - | 404.2 (±34.1) | 112.7 (±9.2) | 3.6 : 1.0 | 0 | - | 16.4/6.2 | - |
| 27 | Unidentified | - | - | 1128.8 (±65.3) | 136.1 (±10.5) | 8.3 : 1.0 | 0.001 | - | 16.3/6.3 | - |
| 28 | Unidentified | - | - | 8.8 (±2.4) | 982.0 (±52.5) | 1.0 : 111.6 | 0.001 | - | 14.8/6.6 | - |
| 29 | Unidentified | - | - | 686.9 (±36.0) | 119.8 (±7.0) | 5.7 : 1.0 | 0.001 | - | 14.6/5.6 | - |
zIdentification of certain selected proteins cites results as revealed by proteomic analyses in previous studies (Jae Ho Kim et al. 2020. A Large-Scale Proteome Analysis of Proteins Expressed in the Peel of ‘Hongro’).
Discussion
‘Hongro’
Carbohydrate metabolism
Spot No. 1 was expressed more strongly by 3.0 times in ‘Hongro’ than in ‘Greenball’ and was identified as an NADP+- dependent malic enzyme (EC 1.1.1.40). Malate accounts for most of the organic acids in apples (Wu et al. 2007) and significantly impacts several fruit characteristics, including taste, quality, coloration, storage, and ripening (Tijero et al. 2021). The NADP+-dependent malic enzyme catalyzes the production of pyruvate, CO2, and NADPH using malate and NADP+ as substrates (Saigo et al. 2013; Selinski et al. 2014). Pyruvate is oxidized in the mitochondria to generate ATP, and NADPH acts as a reducing agent during anabolic processes such as the synthesis of nucleic acids, glucose, lipids, and proteins in vivo (Spaans et al. 2015; Tijero et al. 2021). Therefore, it was expected that ‘Hongro’ would have higher production rates of ATP and NADPH compared to those of ‘Greenball’. Spot No. 2 was more strongly expressed by 2.7 times in ‘Hongro’ than in ‘Greenball’ and was identified as enolase-like. Enolase (EC 4.2.1.11), also known as phosphopyruvate hydratase, is an enzyme that catalyzes the conversion of 2-phosphoglycerate (PGA) into phosphoenolpyruvate (PEP) during glycolysis and belongs to the family of hydrolases that cleave carbon-oxygen bonds (Prabhakar et al. 2009). PEP generated from PGA is converted to pyruvic acid and synthesizes ATP. Pyruvic acid is continuously delivered to the Krebs cycle to generate NADH and FADH2, and ultimately synthesizes ATP through oxidative phosphorylation in the mitochondria. Therefore, it was expected that ‘Hongro’ would have a higher ATP production rate than ‘Greenball’. Spots No. 3 and 9 were more strongly expressed by 7.4 and 9.2 times, respectively, in ‘Hongro’ than in ‘Greenball’ and were identified as FDH (EC 1.2.1.2), mitochondrial. In higher plants, FDH is an NAD-dependent enzyme mainly localized in the mitochondria, catalyzing the oxidation of formate to CO2 and producing NADH, a respiratory substrate (Hourton-Cabassa et al. 1998). Formate is produced through various pathways and is used as a major precursor for C1 metabolism or is oxidized by FDH (Iganberdiev et al. 1999; Ambard-Bretteville et al. 2003). In most plant species, this enzyme is highly abundant in the mitochondria of non-green tissues, such as tubers and storage roots, and is less abundant in photosynthetic tissues (Colas des Francs-Small et al. 1992). With regard to ‘Hongro’, given that the expression rate of FDH is higher than that of ‘Greenball’, the synthesis rate of ATP from NADH was also expected to be higher. Additionally, FDH and formate increase under various stress conditions, such as cold, dryness, and wounding (Hourton-Cabassa et al. 1998). ATP is required for responses that increase tolerance to various types of stress. Therefore, it was expected that ‘Hongro’ would have a higher rate of ATP production from NADH and higher tolerance to various types of stress compared to ‘Greenball’. Spot No. 13 was identified as glyoxalase I (EC 4.4.1.5) and was more strongly expressed by 3.4 times in ‘Hongro’ than in ‘Greenball’. Methylglyoxal (MG), a substrate of glyoxalase I, is mainly produced as a by-product of glycolysis involving dihydroxyacetone phosphate (DHAP) and glyceraldehyde 3-phosphate (G3P). Some of it is also produced through the catabolism of acetone and threonine, the peroxidation of lipids, and the decomposition of glycated proteins (Bellier et al. 2019; He et al. 2020). MG can react with various cellular components, including proteins and nucleic acids, in organisms, inducing degradation and causing various types of cytotoxicity, such as aging and oxidative stress and giving rise to mutations as well (Thornalley 2003; Bierhaus et al. 2012). Both bacteria and eukaryotes have a glyoxalase detoxification system consisting of two enzymes that work sequentially to detoxify reactive aldehydes, including toxic MG. Glyoxalase I is the enzyme that acts during the first step of this detoxification system and catalyzes the conversion of MG to S-d-lactoylglutathione using reduced GSH (Kwon et al. 2013; Farrera and Galligan 2022). Therefore, it was expected that ‘Hongro’ would have greater detoxification ability against reactive aldehydes, including MG, which is cytotoxic, compared to ‘Greenball’.
Plant hormones
Spot No. 6 was expressed more strongly by 3.1 times in ‘Hongro’ than in ‘Greenball’ and was identified as 12-OPR (EC 1.3.1.42) 2-like. In plants, OPRs are a subgroup of flavin-dependent oxidoreductases that catalyze the NADPH-dependent reduction of 12-oxo-cis-10,15-phytodienoate to 3-oxo-2-(cis-2’-pentenyl)-cyclopentane-1-octanoate in the biosynthetic pathway of JA (Schaller et al. 2000). JA and its derivatives act as signaling molecules for the regulation of various developmental processes, such as plant defense processes against biotic agents including herbivores and pathogens, senescence, tuber formation, and seed germination (Engelberth et al. 2007; Tani et al. 2008). Therefore, it was expected that ‘Hongro’ would have a higher JA biosynthesis rate and greater gene regulation ability in various developmental processes than ‘Greenball’.
Amino Acid Metabolism
Spot No. 8 was expressed more strongly by 14.9 times in ‘Hongro’ than in ‘Greenball’ and was identified as β-CAS 1. In plants, cyanide is produced during the metabolism of glyoxylate, nitrile, cyanogenic glycosides, and cyanolipids, and during ethylene biosynthesis (Machingura et al. 2016). Cyanide irreversibly binds to mitochondrial cytochrome C oxidase and interferes with the electron transfer process and thus must be quickly converted to a non-toxic product (Mano and Soman 2020). The main enzyme that detoxifies cyanide is β-CAS, which catalyzes the conversion of toxic cyanide to non-toxic β-cyanoalanine by binding to cysteine (Lai et al. 2009; Yu et al. 2012). In apples, β-CAS activity is known to be mainly present in both the fruit and seeds (Han et al. 2007; Machingura et al. 2016). In climacteric fruits such as apples, a large amount of ethylene is biosynthesized and released during maturation (Alexander and Grierson 2002). Therefore, it was expected that ‘Hongro’ would have stronger detoxification ability for toxic cyanide than ‘Greenball’.
Antioxidant Enzyme
Spot No. 17 was expressed more strongly by 5.4 times in ‘Hongro’ than in ‘Greenball’ and was identified as APX ( EC 1.11.1.11) 2, cytosolic. Plants produce various types of reactive oxygen species (ROS), such as hydrogen peroxide (H2O2), not only under abiotic forms of stress such as drought, heat, and in the presence of heavy metals, but also during normal metabolic processes in the chloroplasts and mitochondria (Caverzan et al. 2012; Foyer and Noctor 2013). These ROS can damage various cellular components, including proteins, lipids, and nucleic acids, necessitating diverse detoxification systems (Mittler 2002). APX is a plant-specific heme-containing peroxidase that catalyzes the conversion of H2O2 into H2O using ascorbate as an electron donor (Foyer and Noctor 2013). APX can be categorized into four types, cytosolic APX (cAPX), mitochondrial APX (mitAPX), chloroplastic APX (chlAPX), and peroxisomal APX (pAPX), based on localization within the cell. Among these, cAPX has the APX1 and APX2 isoforms (Pandey et al. 2017). Therefore, it was expected that ‘Hongro’ would be more able to remove toxic H2O2 than ‘Greenball’. Spot No. 19 was identified as GST (EC 2.5.1.18)-like and was expressed more strongly by 19.8 times in ‘Hongro’ than in ‘Greenball’. GST catalyzes the conjugation reaction between the thiol group of glutathione (GSH) and endogenous and exogenous toxic electrophilic compounds (Cummins et al. 2013; Estévez and Hernández 2020). Additionally, GST can scavenge ROS produced under biotic and abiotic types of stress (Dixon et al. 2010; Estévez and Hernández 2020) and can act directly to scavenge toxic endogenous products, functioning as a glutathione peroxidase (Marrs 1996). Therefore, it was expected that ‘Hongro’ would have greater antioxidant ability to scavenge ROS than ‘Greenball’, as well as the ability to detoxify various types of xenobiotics.
Allergens
Spot No. 21 was expressed more strongly by 12 times in ‘Hongro’ than in ‘Greenball’ and was identified as an agglutinin alpha chain-like compound. Agglutinin is a type of antibody that has properties similar to those of lectin, a glycoprotein widely found in foods (Vojdani 2015). Lectin in plants can act as a defense mechanism against insects, bacteria, and fungi (Sauvion et al. 2004). Lectins can enter the circulatory systems of animals (Hamid and Masood 2009) and may cause autoimmune diseases by binding to glycoproteins and glycolipids present on the surfaces of epithelial cells, lymphocytes, and red blood cells (Cordain et al. 2000; Vasconcelos and Oliveira 2004). Dietary lectins and agglutinins are known to be present in several foods, such as wheat, soybeans, peanuts, tomatoes, and potatoes (Vojdani 2015). Therefore, it was predicted that ‘Hongro’ may have a stronger defensive effect against biological factors such as insects compared to ‘Greenball’, but it was also expected to be more likely to cause autoimmune diseases harmful to the human body.
‘Greenball’
Carbohydrate metabolism
Spot No. 4 was expressed more strongly by 2.1 times in ‘Greenball’ than in ‘Hongro’ and was identified as protein DJ-1 homolog D-like. DJ-1 homologs expressed in Arabidopsis include DJ-1a (Xu et al. 2010), DJ-1c and DJ-1b (Lin et al. 2011), and DJ-1 d (Kwon et al. 2013). In most organisms, DJ-1 homolog proteins are involved in the response to oxidative stress and are known to have glyoxalase activity (Farrera and Galligan 2022). Glyoxalase 1 (spot 13) was expressed at a higher level in ‘Hongro’ than in ‘Greenball’. Therefore, it was expected that ‘Greenball’, unlike ‘Hongro’, would have a higher expression level of DJ-1 homolog D, which has a similar function to glyoxalase, and thus would scavenge reactive aldehydes such as MG. Spot No. 11 was expressed more strongly by 11.1 times in ‘Greenball’ than in ‘Hongro’ and was identified as thiamine thiazole synthase (EC 2.8.1.10), chloroplastic. Thiamine (vitamin B1) is an essential compound for organisms, biosynthesized in plants, fungi, bacteria, and some protozoa, and contains pyrimidine and thiazole moieties within the molecule (Webb et al. 2007; Begley et al. 2008). In plants, these rings are biosynthesized and combined in plastids, and thiamine thiazole synthase catalyzes the synthesis of the thiazole moiety (Goyer 2010; Gerdes et al. 2012). This enzyme is also associated with increased tolerance to several types of stress, including drought and salt (Tunc-Ozdemir et al. 2009; Li et al. 2016). Biosynthesized thiamine is phosphorylated and acts as a coenzyme in metabolic pathways such as sugar and amino acid catabolism (Ciszak et al. 2003; Fitzpatrick and Chapman 2020). In humans, thiamine deficiency can cause various diseases, including beriberi, with symptoms such as weight loss, malaise, and confusion (McCandless 2010; Smith et al. 2021). Therefore, it was expected that ‘Greenball’ would have a higher biosynthetic rate of thiamine, which acts as a coenzyme and is beneficial to humans, than ‘Hongro’.
Photosynthesis
Spot No. 5 was expressed more strongly by 3.3 times in ‘Greenball’ than in ‘Hongro’ and was identified as RuBisCO (EC 4.1.1.39) activase, chloroplastic-like. For RuBisCO, the lysine at the active site undergoes carboxylation for carbon assimilation, is converted to the active form through binding with Mg2+ ions, and is subsequently combined with the substrates RuBP and CO2 (Andersson 2008). Meanwhile, when phosphate sugars such as xylulose-1,5-bisphosphate, 2,3-pentodiulose-1,5-bisphosphate or 2-carboxy-D-arabinitol-1-phosphate, which are produced at low concentrations, bind to apo-type or active-type RuBisCO, the enzyme becomes inactive (Bhat et al. 2017). RuBisCO activase, belonging to the ATPase superfamily, is a type of molecular chaperone that promotes the release of inhibitory sugar phosphate from inactive RuBisCO by hydrolyzing ATP. This action remodels RuBisCO into its active form (Portis et al. 2008). RuBisCO activase was found to have increased expression levels under various forms of abiotic stress, such as drought, high temperatures, and salt (Rollins et al. 2013; Chen et al. 2015). Therefore, it was expected that ‘Greenball’ would have stronger RuBisCO activity for CO2 fixation than ‘Hongro’. Spot No. 16 was expressed more strongly by seven times in ‘Greenball’ than in ‘Hongro’ and was identified as oxygen-evolving enhancer protein 1, chloroplastic. This protein is a 33 kDa subunit, known as one of the components of the oxygen-evolving complex (OEC) of photosystem II (PS II) (Lydakis-Simantiris et al. 1999). OEC consists of the three exogenous proteins of 17, 24, and 33 kDa and other endogenous thylakoid membrane proteins that photocleave water molecules and generate oxygen (Yi et al. 2005). Therefore, it was expected that ‘Greenball’ would have more distinct OEC development in chloroplasts than ‘Hongro’ and that oxygen release and electron transfer processes through water photolysis would be more active. Spot No. 18 was expressed more strongly by 4.9 times in ‘Greenball’ than in ‘Hongro’ and was identified as a PPD-containing protein. PPD protein is one of the PS I assembly factors in the thylakoid membranes of green algae and land plants and is homologous to the PsbP protein, a constituent protein of PS II (Ishihara et al. 2007; Liu et al. 2012). On the stromal side of PS I, PsaC, PsaD, and PsaE proteins, which promote binding with ferredoxin and ferredoxin-NADP reductase, are located adjacent to each other, and on the luminal side, PsaA, PsaB, PsaF, and PsaN proteins, which form a docking site for plastocyanin, are adjacent to each other (Roose et al. 2014). Here, PPD proteins interact with the luminal loops of PsaA and PsaB, facilitating their proper folding and integration into the thylakoid membrane (Liu et al. 2012; Roose et al. 2014). Therefore, it was expected that ‘Greenball’ would facilitate smoother PS I assembly compared to ‘Hongro’ and that the subsequent electron transfer and NADP+ reduction activities would also be greater.
In summary, among the proteins with higher expression rates in ‘Hongro’ than in ‘Greenball’, NADP+-dependent malic enzyme, enolase-like, and FDH are enzymes that act in the ATP biosynthesis system. Accordingly, ‘Hongro’ was expected to have a higher ATP biosynthesis rate than ‘Greenball’. Given that NADP+-dependent malic enzyme also catalyzes the biosynthesis of NADPH, which acts as a reducing agent within the cell, it was expected that the synthesis of various biomolecules, including glucose, would increase in ‘Hongro’ compared to ‘Greenball’. In addition, because the expression rates of glyoxalase I, 12-OPR 2-like, β-CAS 1, APX 2, GST-like, and agglutinin alpha chain-like were higher in Hongro than in Greenball, the scavenging of toxic aldehydes including MG (glyoxalase I), the biosynthesis of JA (12-OPR 2-like), the detoxification of toxic cyanide (β-CAS 1), the antioxidant ability (APX 2, GST-like), and allergenicity (agglutinin alpha chain-like) were also expected to be higher in ‘Hongro’.
Among the proteins with higher expression rates in ‘Greenball’ than in ‘Hongro’, DJ-1 homolog D-like is expected to have glyoxalase activity. For this reason, it was expected that ‘Greenball’ would have the ability to scavenge toxic aldehydes, including MG, similar to those in red beans. In particular, ‘Greenball’ was expected to have a higher photosynthetic rate than ‘Hongro’ because it has a green-yellow peel and has high expression rates of RuBisCO activase, PS I (oxygen-evolving enhancer protein 1, chloroplastic) and Ⅱ (PPD-containing protein) assembly proteins. Meanwhile, among the proteins whose expression rates differed between ‘Hongro’ and ‘Greenball’, no proteins directly related to the development of skin color were identified. Spot 20, which had an expression rate approximately 9.3 times higher in ‘Hongro’, and spot 10, which had an expression rate approximately 18.0 times higher in ‘Greenball’, were identified as uncharacterized protein LOC103408523 and uncharacterized protein RDQH01000341, respectively. In addition, spots 7, 22, 23, 24, 25, and 28, which showed higher expression levels in ‘Hongro’, and spots 12, 14, 15, 26, 27, and 29, which showed higher expression levels in ‘Greenball’, were not identified. If these proteins are identified through additional research, it is expected that the types and functions of proteins that play important roles in the varietal characteristics of ‘Hongro’ and ‘Greenball’ will be more clearly elucidated.
