

서 언
재료 및 방법
실험재료
생장조정제 및 과립밀도 처리
과실 생육 및 품질 조사
과피의 해부학적 특성 및 경도 측정
열과 발생 특성 조사
통계분석
결과 및 고찰
과립밀도에 따른 과립 비대와 과실품질
과립밀도에 따른 열과 발생
과립 부위별 과피 특성
서 언
국내 포도 산업은 과거 ‘캠벨얼리’와 ‘거봉’ 등 구미교잡종(Vitis labruscana)에서 ‘샤인머스캣’을 중심으로 한 유럽계(V. vinifera) 품종 재배가 급격히 확산되고 있다. 한국농촌경제연구원 농업관측정보(KREI 2023)에 따르면 2024년 ‘샤인머스캣’은 6,182ha로 전체 재배면적의 42.6%를 차지하고, 기존 재배 품종 외 다양한 품종들이 3.4%(약 500ha)로 전년 대비 8.8% 증가할 것으로 추정되고 있다.
‘홍주씨들리스’는 농촌진흥청에서 ‘Italia’와 ‘Perlon’의 교배조합을 통해 2013년 최종 선발한 품종으로 평균 당도 18.4°Brix, 산 함량 0.57%, 과피색이 적색인 무핵 품종이다(Roh et al. 2018). 반면 ‘홍주씨들리스’는 무핵 품종이나 농업 현장에서는 대과 생산 및 식감 향상을 위해 gibberellin과 cytokinin류 생장조절제 처리가 관행적으로 이루어지고 있으며, 정립되지 않은 재배체계는 과다 착과에 따른 열과, 착색 불량 등 다양한 생리장해가 발생되고 있는 상황이다.
일반적으로 포도의 열과 발생에는 과피의 구조(Zhang et al. 2021; Zhang et al. 2024), 과다 착과(Choi et al. 2024), 온도(Kim et al. 2021), 토양수분(Considine and Brown 1981; Kim et al. 2016) 등 유전 ‧ 재배 ‧ 환경적으로 다양한 요인들이 영향을 미치는 것으로 알려져 있으며(Choi and Choi 2023), 직접적인 원인으로는 과실 내부에서 증가된 팽압에 의해 나타난다(Andersen and Richardson 1982).
재배적 요인에 해당하는 과다 착과는 당도 증가와 산 함량 감소를 늦춰 성숙(착색 불량 등)을 지연시킨다(Ozer et al. 2012). 과방의 제한된 면적 내 과립간 생장 압박(Considine and Brown 1981)은 과분과 과피 세포의 정상적 발달을 저해하여(Choi et al. 2024), 열과 발생을 증가시키며(Shim et al. 2007), 저장양분 불균형에 따른 화진현상(Shim et al. 2007)과 저온피해(Stergios and Howell 1977)를 유발할 수도 있다. 또한 송이 내 공기 유동을 방해하여 병 발생을 증가시키는 원인이 되기도 한다(R’Houma et al. 1998).
최근 젊은 소비계층을 중심으로 고당도, 무핵, 특징적인 외관과 색을 가진 포도 수요가 증가하고 있으며, ‘홍주씨들리스’도 재배가 확대되고 있는 품종 중 하나이다. 본 연구는 생장조정제를 활용한 ‘홍주씨들리스’의 대과 생산 시 착립밀도에 따른 과실품질을 비교하고, 과피특성을 조사하여 열과 발생 양상에 대해 구명함으로써 생리장해 경감을 위한 최적 착립밀도 제시 및 기초적인 재배정보 획득을 위해 수행되었다.
재료 및 방법
실험재료
실험은 전라북도 익산시 왕궁면 소재(35°57’05”N, 127°04’19”E, 해발고도 18m) 포도 과원(비가림 시설)에서 2년간(2021–2022년) 수행하였다. 재식거리 3×3m로 식재된 3년생 포도 ‘홍주씨들리스’를 대상으로 병해충 피해가 없고, 수세가 균일한 시험수 5–8주를 선정하여 조사하였으며 농촌진흥청 농업기술길잡이 “포도(RDA 2021)”에 준하여 재배관리 하였다.
생장조정제 및 과립밀도 처리
관행재배와 동일한 조건에서 과립밀도를 달리하기 위해 생장조정제 처리는 만개 후 1–2일과 만개 후 14일에 2회 처리하였다. 생장조정제 처리 전 첫번째 꽃이 개화한 시기에 화방(flower cluster)의 크기에 따라 하단 약 3–5cm 정도를 제외한 상단부 지경을 모두 제거하였으며, 생장조정제 처리는 GA3(IAP Gibberellin SG, Jahngryu Industries Co., Ltd., Cheongju, Korea) 25mg·L-1, forchlorfenuron(Fulmet, Arysta LifeScience Inc., Tokyo, Japan) 2.5mg·L-1가 혼용 희석된 용액에 화방을 침지하였다.
생장조정제 처리가 완료된 과방은 만개 후 20일경 송이당 과립 수를 35, 45, 55, 65립으로 적립하였고, 수확기의 과립밀도는 각각 1.7, 2.2, 2.7, 3.1개·cm-1이었다. 시험수 1주에 모든 처리구의 7개 이상 과방을 임의배치 하였으며, 신초당 과방의 수는 평균 1.0개로 적방하였다.
과실 생육 및 품질 조사
과립 종경, 횡경, 과립지수(L/D ratio), hunter a 값의 변화를 조사하고자 2021(만개 후 15, 25, 35, 47, 57, 67, 76, 86, 100, 114, 127일)–2022(만개 후 20, 31, 43, 52, 60, 68, 79, 89, 101, 116, 125일)년 8–15일 간격으로 총 11회 조사하였으며, 처리별 10개 과방의 최하단에 위치한 2개의 과립을 대상으로 종경과 횡경을 측정하였다. hunter a 값은 디지털 색차계(CR-10plus, Konica Minolta Inc., Osaka, Japan)를 이용하여 과립 적도부를 2회 측정한 후 평균값을 산출하였다.
과실품질은 2021–2022년 만개 후 130일에 과방을 채취하여 5개의 과방을 4반복으로 조사하였다. 과방중은 디지털 전자저울(PB602-S, Mettler Toledo Inc., Zwingenberg, Germany)을 이용하여 처리된 모든 과방을 측정하였고, 각 과방별 과립 수를 계수하여 과방중을 과립 수로 나눈 값을 평균 과립중으로 산출하였다. 가용성 고형물 함량은 과방당 5개의 과립을 무작위로 채취하여 착즙액을 혼합한 후 디지털 굴절당도계(PR-101α, Atago Co. Ltd., Tokyo, Japan)로 측정하였다. 착즙한 과즙은 증류수로 1:10의 비율로 희석하여 전위차적정기(Easyplus titration, Mettler Toledo Inc., Zwingenberg, Germany)로 0.1N NaOH를 pH 8.1이 되는 시점까지 적정한 후 tartaric acid 상당량으로 환산하였다.
과피의 해부학적 특성 및 경도 측정
과립 부위별 과피 세포를 검경하고자, 만개 후 120일 경 과실을 채취하여 고정(fixation), 탈수(dehydration), 포매(embedding) 과정을 거쳐 epon block을 제작하였다(Park 1995). 제작이 완료된 epon block은 ultramicrotome(Ultracut UCT, Leica Co., Wetzlar, Germany)으로 두께 1µm 절편을 채취하였고, 염색(periodic acid-schiff)한 후 광학현미경(BX51, Olympus Co., Tokyo, Japan)으로 검경하였다. 대상은 과립 부위별 외표피(outer epidermis), 아표피(hypodermis) 층 수와 두께, 1mm2당 아표피 세포 수, 표피 두께(외표피와 아표피 포함)를 조사하였다.
과피 표면의 과분(epicuticular wax)과 주두흔(stigma-stain)을 검경하기 위해 과피 세포 검경과 동일한 시기 과실을 채취하였으며, ‘홍주씨들리스’와의 상대적 비교를 위해 열과 발생이 적은 것으로 알려져 있는 ‘샤인머스캣’을 대조 품종으로 사용하였다. 현미경 검경은 FE-SEM(Field emission scanning electron microscope)을 사용하여 Choi et al.(2024)의 방법을 적용하였고, 과분은 2,000배율, 주두흔은 50배율로 검경하였다.
동일한 시기에 채취한 과립의 경도도 과경부, 적도부, 과정부로 구분한 후 과실 물성측정기(SD-700, Sun Scientific Co., Ltd., Tokyo, Japan)로 측정한 평균값을 이용하였다.
열과 발생 특성 조사
열과 발생은 과립비대와 동일한 대상과 시기에 조사하였다. 열과 발생률은 착립수준에 따라 열과가 발생한 과립이 있는 과방과 과방당 열과된 과립 수를 계수하여 백분율로 산출하였다. 또한 과립 부위를 과립경이 있는 과경부(stem-end), 과립 중앙 적도부(equatorial), 주두흔이 위치한 과정부(stylar-end)로 구분하였으며, 열과가 발생한 위치를 전체 열과립수의 백분율로 표현하였다.
통계분석
조사 데이터는 통계프로그램(Sigmaplot 14.0, Systat Inc., San Jose, CA, USA)을 이용하여 Duncan의 다중검정, 요인분석과 그래프를 작성하였다.
결과 및 고찰
과립밀도에 따른 과립 비대와 과실품질
2년간(2021–2022) 포도 ‘홍주씨들리스’의 생육시기별 과립 종경과 횡경, hunter a값의 변화 결과는 Fig. 1과 같다. 2년간 과립 종경과 횡경은 초기(만개 후 30일경) 급격한 비대 이후 수확기까지 완만하게 증가하는 경향을 보였다. 과립밀도 처리간 비대 양상은 유사하였으며, 수확기에 이르러 과립밀도가 높을수록 비대량은 적은 경향을 보였다(Fig. 1A, 1B, 1D, and 1E). 적색계 품종의 적색 정도를 나타내는 hunter a값은 만개 후 약 60–80일까지 급격한 증가를 나타낸 후 완만해지는 경향을 보였는데, 만개 후 60–80일이 ‘홍주씨들리스’의 착색기로 판단되었다(Fig. 2). 과립밀도에 따른 hunter a 값은 과립밀도 3.1개·cm-1에서 1.7, 2.2, 2.7개·cm-1 처리보다 낮은 경향이었다(Fig. 1C and 1F).
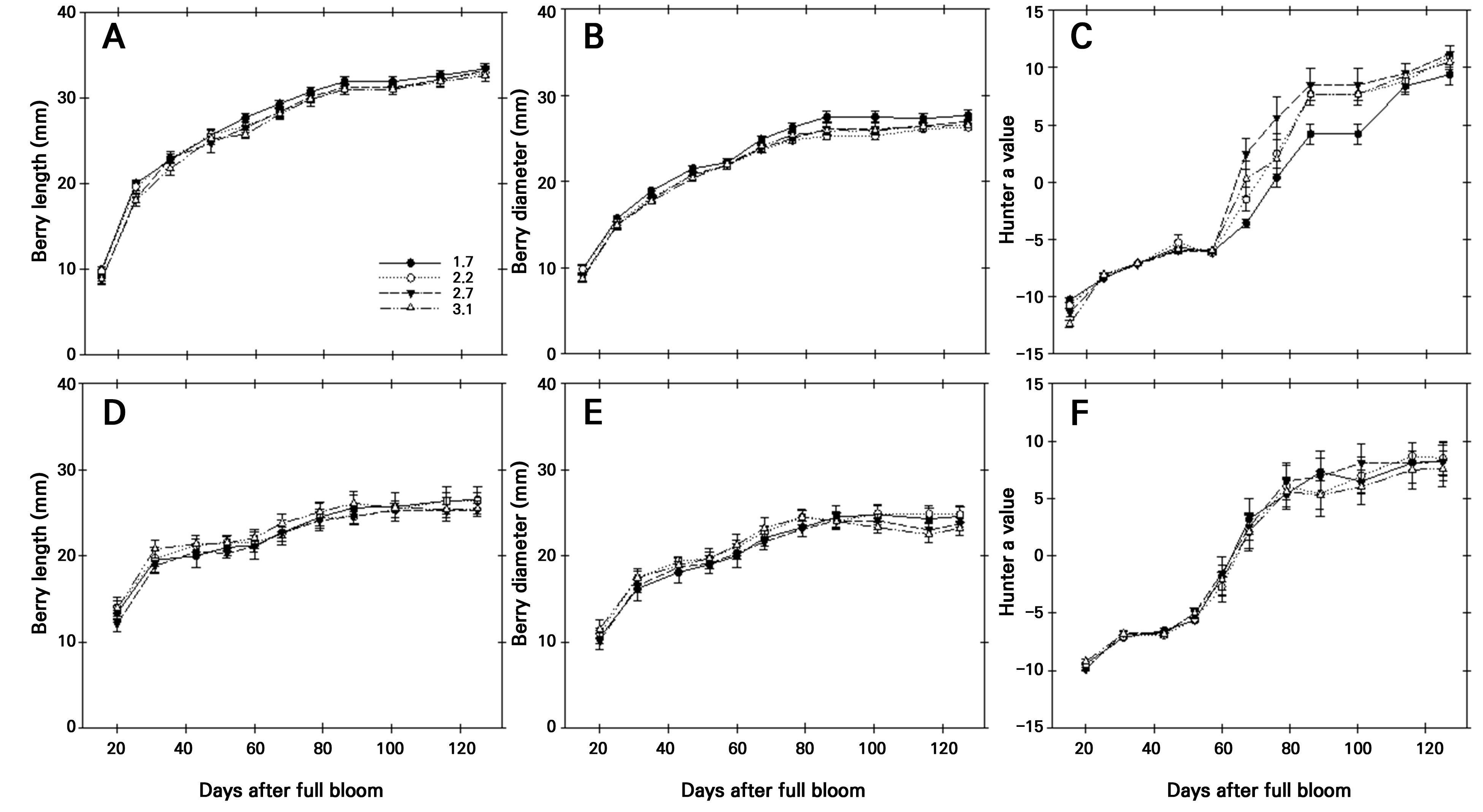
Fig. 1.
Effect of berry density on the berry length (A and D), berry diameter (B and E), and hunter a value (C and F) during the growing stage of ‘Hongju Seedless’ grape in 2021 (A-C) and 2022 (D-F). Vertical bars represent the standard error of the means (n = 15). The legend indicates the number of berries per 1 cm-1 of cluster.

Fig. 2.
Photos of cluster during the growing stage and berry density of ‘Hongju Seedless’ grapes in 2022. These photos were taken at 20, 31, 43, 52, 60, 68, 79, 89, 101, and 116 days after full bloom in order from the left. A1-A10: 1.7, B1-B10: 2.2, C1-C10: 2.7, and D1-D10: 3.1 berries per 1 cm-1 of cluster. The red scale bars indicate 6 cm.
수확기 과실품질 조사 결과(Table 1), 과립밀도가 증가함에 따라 과방중, 과립수는 높았으며, 통계적 차이는 없었으나 과립중, hunter a값, 산 함량은 낮은 경향을 보였다. 이와 같은 결과는 2021년과 2022년 모두 유사한 결과를 나타내었다. 특히 과방중의 경우 2022년 과립밀도 3.1개·cm-1 처리에서 488.5g으로 2.2개·cm-1(508.1g), 2.7개·cm-1(619.2g) 처리에 비해 낮았는데, 이는 상대적으로 과방당 열과 발생이 많아 수확기 평균 과중이 낮아진 결과로 판단되었다.
Table 1.
Effects of berry density on the fruit quality at harvest stage (130 days after full bloom) of ‘Hongju Seedless’ grapes in 2021 and 2022
|
Surveyed year |
Berry density (berries·cm1) |
Cluster weight (g) |
Number of berries (per cluster) |
Berry weight (g) |
Hunter a valuez |
Soluble solids (°Brix) |
Titratable acidity (%) |
| 2021 | 1.7 | 371.5 cy | 35.9 d | 10.4 a | 16.2 a | 16.8 a | 0.42 a |
| 2.2 | 396.1 c | 42.8 c | 9.3 a | 15.6 a | 16.9 a | 0.44 a | |
| 2.7 | 492.6 b | 51.3 b | 9.7 a | 15.2 a | 16.7 a | 0.44 a | |
| 3.1 | 549.0 a | 60.1 a | 9.2 a | 14.7 a | 16.9 a | 0.51 a | |
| 2022 | 1.7 | 367.6 c | 32.2 c | 11.5 a | 8.9 a | 17.0 a | 0.41 b |
| 2.2 | 508.1 b | 44.1 b | 11.5 a | 7.7 a | 16.6 ab | 0.45 ab | |
| 2.7 | 619.2 a | 53.6 a | 11.5 a | 8.7 a | 16.5 ab | 0.46 a | |
| 3.1 | 488.5 b | 52.4 a | 9.9 a | 9.1 a | 15.9 a | 0.50 a | |
| Significance | |||||||
| Surveyed year (A) | NS | NS | * | ** | NS | NS | |
| Berry density (B) | NS | * | NS | NS | NS | * | |
| A × B | ** | * | NS | NS | NS | NS | |
포도 과립은 수정 후 세포 분열과 비대 과정을 거치면서 생장 양상이 2중 S자 곡선을 보이는데(Coombe 1992), 과립의 비대와 착색 변화에는 과방당 착립된 과립 수가 영향을 미칠 수 있다(Choi et al. 2024). 특히 과다 착과는 착색불량(Jung et al. 2014), 당 함량 감소와 산 함량 증가(Matsumoto et al. 2007)로 결국 성숙을 지연시킨다(Takahashi 1986). 따라서 본 실험의 Fig. 1과 Table 1에서와 같이 ‘홍주씨들리스’의 과립밀도 증가와 함께 과방중과 과방당 과립 수의 증가가 관찰되기는 하나 hunter a값과 가용성 고형물 함량의 감소와 산 함량의 증가가 관찰되어 결과적으로 과실 성숙이 지연된 것으로 판단되었다.
과립밀도에 따른 열과 발생
포도 ‘홍주씨들리스’의 열과 발생(Fig. 3)은 2021년과 2022년에 각각 만개 후 76일, 60일에 발생이 시작되어 주로 과실 착색기에 나타남을 확인할 수 있었다. 2021년 열과된 과립이 있는 과방의 비율(Fig. 3A)은 과립밀도 1.7, 2.2, 2.7, 3.1개·cm-1 처리에서 각각 33.3%, 46.7%, 46.7%, 73.3%로 조사되었고, 2022년에도 2021년과 마찬가지로 과립밀도가 높아짐에 따라 증가하는 경향이었다. 과방당 열과된 과립의 비율도 2021년(과립밀도 순서대로 2.7%, 5.5%, 6.1%, 18.3%), 2022년(2.7%, 4.9%, 13.5%, 24.7%) 모두 과방 비율과 유사한 경향을 보였으며, 이와 같은 결과는 선행 연구(Choi and Choi 2023; Choi et al. 2024)와 일치하였다. 추가적으로 과립을 과경부, 적도부, 과정부로 구분하여 열과 발생 부위를 조사한 결과(Fig. 3C), 2021년 과정부 95.0%, 과경부 5.0%, 2022년에는 과정부 74.1%, 과경부 22.2%, 적도부 3.6%로 조사되어 상대적으로 과정부 열과가 많았다(Son et al. 2007). 과립 과정부 열과는 초기 약 1–3cm 정도의 미세 균열(Fig. 4A)이 관찰된 이후, 미생물 감염에 따라 부패가 시작되었고(Fig. 4B and 4C), 흘러내린 과즙과 병해충 피해로 인해 상품성을 잃는 경우(Fig. 4D)도 발생하였다(Meneguzzo et al. 2008). 또한 연차간 차이에서는 2021년에 비해 2022년 열과 발생량이 많았는데, 이는 강우 일수, 수령 경과에 따른 수세 증가 등이 영향을 준 것으로 추정되었다.
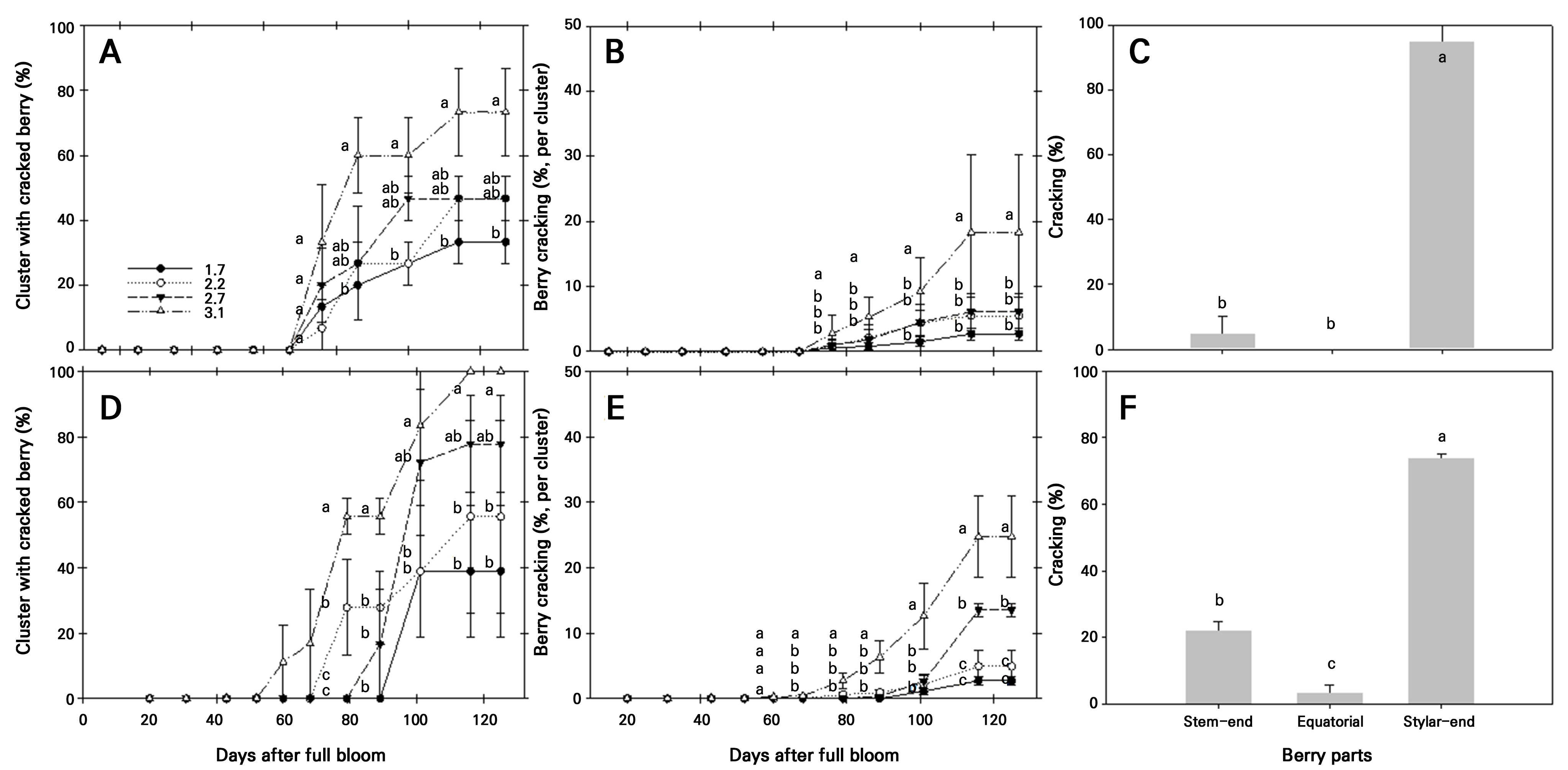
Fig. 3.
Effects of berry density on a cluster with cracked berry (A and D), berry cracking per cluster (B and E), and occurrence parts (C and F) during the growing stage of ‘Hongju Seedless’ grapes in 2021 (A-C) and 2022 (D-F). Vertical bars represent the standard error of the means (n = 4). The legend indicates the number of berries per 1 cm-1 of cluster. Different letters indicate statistically significant differences according to the DMRT test at p < 0.05.

Fig. 4.
Cracking degree at the stylar-end parts of ‘Hongju Seedless’ grape berries. A: early stage of cracking occurrence, B and C: rotting caused by pathogen infection in cracked berries, D: loss of marketability of cluster. The red arrows denote cracked berries. The red solid and dotted scale bars indicate 2 cm and 3 cm, respectively.
열과는 과실 생장과 발달, 성숙 과정, 그리고 저장 조건에서도 발생할 수 있다(Khadivi-Khub 2009). 열과의 중요 메커니즘은 과실 내부로의 급속한 수분 흡수로 증가된 팽압이며(Yamamoto et al. 1990), 이로 인해 과피의 기계적 결함을 초래하여 열과로 이어지게 된다(Dean et al. 2016). 열과를 유도하는 여러 요인 중 과다 착과는 과방 내 과립간 압박을 발생시켜 과립 표면의 과분과 과피세포의 정상적 발달을 방해(Choi et al. 2024)하여 열과 발생을 증가시킬 수 있다(Considine and Brown 1981). 또한 수확 전 과피 세포의 신장성이 감소하는 ‘hardening-off’가 일어나며(Matthews et al. 1987), 과피 경도 감소가 빠른 품종(Kataoka et al. 1997)이 급격한 수분 변화에 의해 열과 민감성을 나타낼 수 있다(Khadivi-Khub 2009).
과립 부위별 과피 특성
포도 ‘홍주씨들리스’ 과립 부위별 과피세포와 경도 측정 결과는 Table 2와 같다. 외표피는 과경부, 적도부, 과정부가 1–2개 층으로 구성되어 있었으며, 두께는 과정부가 9.1um로 과경부(10.8um), 적도부(10.6um)보다 얇았으나 통계적 차이는 없었다. 아표피는 5–9개 층으로 구성되어 있었고, 두께는 과정부 102.0um, 적도부 89.7um, 과경부 77cracking.9um 순이었다. 아표피의 두께와 마찬가지로 1mm2당 아표피 세포의 수도 과정부가 7.8개로 가장 많았으며, 적도부(7.3개), 과경부(6.3)로 조사되었다. 또한 과립 부위별 경도는 과정부가 774.1g으로 가장 낮았으며, 과경부(847.6g), 적도부(947.8g) 순으로 나타나 아표피층의 두께와 단위면적 당 아표피 세포 수와는 반대의 결과를 나타내었다.
Table 2.
Anatomical characteristics of the exocarp and hardness of ‘Hongju Seedless’ grapes observed from berry parts, determined 120 days after full bloom
| Position | Outer epidermis | Hypodermis |
Skin thickness (um) |
Hypodermis cell number (per 1 mm2) |
Berry hardness (g) | |||
| Cell layer | Thickness (um) | Cell layer | Thickness (um) | |||||
| Stem-end | 1–2 | 10.8 az | 5–9 | 77.9 b | 88.7 a | 6.3 b | 847.6 b | |
| Equatorial | 1–2 | 10.6 a | 5–9 | 89.7 ab | 100.2 a | 7.3 ab | 947.8 a | |
| Stylar-end | 1–2 | 9.1 a | 6–9 | 102.0 a | 102.0 a | 7.8 a | 774.1 c | |
큐티클, 표피, 아표피 두께가 열과 발생과 부의 관계가 있다는 연구 보고(Zhang et al. 2020; Zhang et al. 2024)는 많은 편이나 반대의 결과도 존재한다(Demirsoy and Demirsoy 2004). 본 실험 결과에서 과정부는 과경부와 적도부에 비해 조밀한 아표피 세포 조직을 갖는 것으로 확인되었으나 경도는 낮았다. 특히 ‘홍주씨들리스’는 열과 발생이 적은 것으로 알려진 ‘샤인머스캣’과 비교했을 때 과분의 구조와 형태(Fig. 5A and 5B)에 있어 큰 차이를 보이지 않았으나 중심부가 함몰되고 2–3배 큰 주두흔 조직(Fig. 5C and 5D)을 가진 것으로 확인되었다. 과점(berry spot 또는 lenticel)은 과피 표면에 존재하는 기공이 광, 온도 및 수분 등과 같은 재배적 환경요인에 의해 코르크화된 것으로 변색기에 가까워질수록 발생정도가 증가할 수 있다(Son et al. 2007). 이러한 과점의 미세균열(micro-cracks)은 과피의 구조적 약화 및 열과의 발생을 조장하는 요인(Borve et al. 2000)이므로 ‘홍주씨들리스’의 과정부는 상대적으로 퇴화된 주두흔 조직이 있어 과경부, 적도부에 비해 열과 발생이 많았던 것으로 생각되었다.
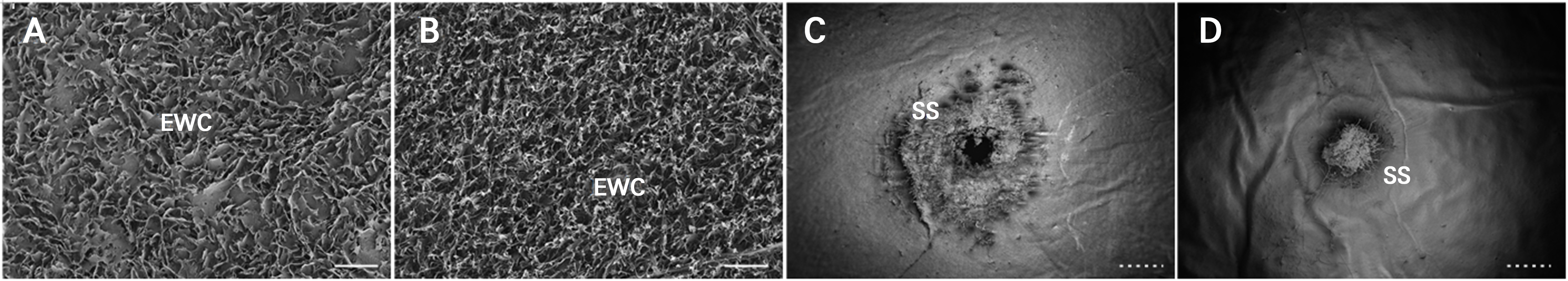
Fig. 5.
Field-emission scanning electron microscopy (FE-SEM) images of epicuticular wax (A and B) and stigma-stain (C and D) determined 130 days after full bloom of ‘Hongju Seedless’ (A and C) and ‘Shine Muscat’ (B and D) grapes. A and B were observed at ×2,000 and C and D were observed at ×50. EWC: epicuticular wax crystals, SS: stigma-stain. The white solid and dotted scale bars indicate 4 µm and 200 µm, respectively.
지금까지의 결과를 요약하면, 포도 ‘홍주씨들리스’는 과립밀도가 높을수록 과방중, 과립 수, 산 함량, 열과 발생량이 증가하였고, 가용성 고형물 함량과 hunter a값이 감소하는 경향을 보여 숙기가 지연되었다. 과정부는 과경부와 적도부보다 아표피 세포의 두께와 단위면적당 세포 수가 많아 조밀한 조직을 보였음에도 경도가 낮고 열과 발생이 많았으며, 이는 과정부의 주두흔이 함돌되어 있고, 상대적으로 ‘샤인머스캣’보다 면적이 커 과피의 기계적 특성이 약화된 것으로 추정되었다. 추가적으로 ‘홍주씨들리스’의 과방당 열과된 과립 발생율을 과립밀도에 적용할 경우, 과립밀도가 높은 처리 순으로 과방당 각각 0.9개, 2.3개, 5.4개, 14.0개의 과립이 열과되었는데, 상대적으로 과립밀도 1.7–2.2·cm-1 범위에서는 외관적 상품성이 악화되지는 않았다(데이터 미제시). 따라서 포도 ‘홍주씨들리스’의 생장조정제 처리 시 과립밀도와 열과 발생량이 비례적으로 증가하므로, 열과 발생량을 줄이기 위해서는 과방 당 35–45립(열과립 발생율이 5% 이내)으로 조절해야 할 것으로 판단되었다. 또한 과방중 증가와 열과 발생을 경감시키기 위해서는 생장조정제의 처리 농도와 횟수, 과방 내 과립의 배치 방법 등 추가적인 관련 연구가 필요할 것으로 생각되었다.
