

서 언
재료 및 방법
식물 재료 및 재배 관리
온실 보광 실험
데이터 수집 및 통계 분석
결 과
보광원에 따른 온실 내 광환경 차이
광원에 따른 영양기관의 생장 및 형태적 특성
보광원에 따른 개화 특성, 절화 품질 및 수량
고 찰
서 언
장미(Rosa hybrida)는 호광성 식물로서 광합성에 미치는 환경요인 중 빛의 영향을 가장 많이 받으며, 절화 품질과 수량을 높이기 위해 충분한 광량(일적산광량)을 확보해야 한다(Dole and Wilkins, 2005; Dole and Warner, 2017; RDA, 2018). 절화 장미의 적정 일적산광량(daily light integral, DLI)은 30mol·m-2·d-1 이상이며, 최소 13mol·m-2·d-1를 권장하고 있다(Dole and Warner, 2017). 우리나라의 겨울철 DLI는 북유럽이나 미국 북부에 비해 높지만, 피복재가 유리보다 광투과율이 낮은 플라스틱이며 보온을 위해 2중 피복에 보온용 피복까지 추가되므로 겨울철에는 최소 권장 DLI보다 낮은 수준의 광량을 보일 것으로 예측된다(RDA, 2018).
절화 재배 시 낮은 DLI는 광합성 저하에 따른 생장 감소, 절화 수량 및 품질 감소, 화아분화 지연 및 화뢰 퇴화(화아 고사, 낙뢰 등) 등 여러 가지로 절화 생산에 부정적인 영향을 미친다(Dole and Wilkins, 2005; Dole and Warner, 2017). 장미 신초의 블라인드(blindness) 현상은 품종에 따라 조금 다른 양상을 보이지만 평균 DLI가 13mol·m-2·d-1보다 낮을 때 증가한다.
장미와 같이 광요구도가 높은 절화의 생산에는 DLI를 높이려는 노력이 필요한데, 인공조명을 이용한 보광(supplemental lighting)은 가장 적극적인 방법이다. 겨울철, 흐린 날, 장마철 등 약광 환경 하에서 보광은 광에너지의 공급을 통한 광합성 증가, 생장 및 발육 속도 증가, 개화 촉진, 수량 및 품질 향상, 기형화 발생 감소 등 많은 혜택을 준다(Dole and Wilkins, 2005; Dole and Warner, 2017). 하지만 보광은 조명기구의 구입 및 설치, 지속적인 전기에너지의 투입, 유지보수 등 상당한 비용을 요구하므로 도입에 신중을 기해야 한다(Fisher et al., 2017).
보광 도입 시 고려사항 중 인공광원의 종류 또한 매우 중요한데, 인공광원에 따라 초기 투자비용, 소비전력, 내구성은 물론 광량 및 광질, 광합성 등 생리적 유효파장 비율 등이 다르기 때문이다(Chon et al., 2018). 최근 소비전력이 매우 낮고, 내구성이 강한 것으로 알려진 발광 다이오드(light-emitting diode, LED)가 온실 보광용으로도 개발되고 있으나, 아직까지는 효율이 높은 고압나트륨 램프(high-pressure sodium lamp, HPS)가 온실 보광용으로 주로 사용되고 있다(Fisher et al., 2017). 절화 장미에 대한 보광 연구는 그간 많이 이루어져 왔으며, HPS 보광이 초장 및 건물중 증가, 절화수명 및 수량 증가 등 좋은 효과를 보여주었다(Armitage and Tsujita, 1979; Bredmose, 1993; Roberts et al., 1993; Maas and Bakx, 1995; Farina and Veruggio, 1996; Mortensen and Fjeld, 1998; Marissen, 2001; Na et al., 2007; Choi et al., 2008; Kwon et al., 2014; Shi and Kim, 2014; Lee and Kim, 2015; Chon et al., 2018; Jeong et al., 2018). HPS는 온실 보광용으로 장점이 많은 광원이지만 청색광(blue light, 400–500nm) 비율이 낮아 일부 작물에서 줄기 신장이 촉진되기도 하지만(Wheeler et al., 1991; Maas and Bakx, 1995), 엽온 증가에 영향으로 잎의 크기를 작게 하기도 한다(Guo et al., 2016). 반면 적색광:원적색광 비율(R:FR)이 높아 줄기 신장이나 엽면적 증가를 억제할 수도 있다(Smith, 1982; Park et al., 2016; Park and Runkle, 2017).
최근 온실 보광용으로 가능성을 가진 플라즈마 램프(sulfur plasma lamp, PLS)가 주요 온실작물을 대상으로 시험되고 있는데(Lee et al., 2014; Kwon et al., 2018; Park et al., 2018a, 2018b), 이 광원은 자연광에 가장 근접한 파장을 방출하며 청색광이 부족한 HPS에 비해 식물 생육에 필요한 광합성유효방사(photosynthetically active radiation, PAR, 400–700nm) 영역의 파장을 고루 방출한다(Kwon et al., 2018). 채소류에 대한 보광 연구에서 PLS 보광이 HPS에 비해 바질(Dörra et al., 2020), 미니 오이(Guo et al., 2016), 오이와 토마토(Hogewoning et al., 2012), 파프리카(Park et al., 2018b) 등의 초장과 엽면적, 건물중을 증가시켰으며, 이는 PLS의 상대적으로 낮은 적색광:원적색광 비율(R:FR)과 적외선 방사에 기인한다고 하였다.
PLS를 온실 화훼작물의 보광에 적용한 연구는 아직 없는 것으로 파악되어 화훼류 중보광 효과가 가장 큰 장미에 대한 적용이 우선 필요하다고 생각된다. 따라서, 이 연구에서는 절화 장미 농가의 플라스틱 하우스 내에서 현재 국내에서 사용하고 있는 보조난방용 HPS를 대조구로 하여 높은 수준의 네덜란드산 HPS와 국내 기업에서 온실보광용으로 시험 중인 PLS를 보광 광원으로써 시설 내에 장미의 생육 및 품질, 생산성에 미치는 영향을 구명하였다.
재료 및 방법
식물 재료 및 재배 관리
경북 경산시 임당동 소재의 장미 플라스틱 하우스(N35°84'82'', E128°74'23'')에서 재배되고 있는 Hybrid Tea 계통의 대형계 절화용 장미(Rosa hybrida) ‘Aqua(아쿠아)’와 ‘Brut(브루트)’를 대상으로 보광 실험을 실시하였다. 가시가 없는 분홍색 품종인 ‘Aqua’는 1년생 접목묘를 식재한 지 2년째가 되었고, 아이보리색의 ‘Brut’는 1년생 접목묘를 정식한 후 6개월째 재배되고 있는 것으로 하였다.
장미는 기비 시비 후 토양에 정식하여 양액을 관주하는 관비재배를 실시하였다. 양액 조성은 다량원소 NO3-N 11.0, NH4-N 2.0, P 3.5, K 4.5, Ca 6.5, Mg 2.0, S 2.0me·L-1와 미량원소 Fe 2.0, Mn 0.5, B 0.25, Zn 0.2, Cu 0.05, Mo 0.05mg·L-1로 조제되었다. 온실 내 최고 기온 27°C 이하로 유지하였으며, 최저 기온은 15–18°C로 유지하였고, 상대습도 및 이산화탄소 농도는 제어하지 않았다.
온실 보광 실험
보광은 예비실험을 위해 2015년 10월 31부터 2016년 4월 15일까지 실시하였고, 본 실험을 위해 2016년 10월 10일부터 2017년 3월 24일까지 실시하였다. 보광광원으로는 고압나트륨 램프(high-pressure sodium lamp, HPS 600W; Son-T, Philips, the Netherlands)와 황 플라즈마 램프(PL sulfur, PLS 700W; LG Electronics, Korea)를 사용하였고, 수관 위의 광합성 유효 광양자속 밀도(photosynthetic photon flux density, PPFD)가 120 ± 20µmol·m-2·s-1가 되도록 설치하였다. 대조구는 광주기의 효과를 상쇄하기 위해 HPS로 10µmol·m-2·s-1가 되도록 설치하였다.
광원의 광도(PPFD)와 광질은 light meter(LI-250A, Li-Cor Inc., Lincoln, NE, USA)와 spectroradiometer(PAR-200, J&C Tech Co., Ltd., Korea)를 이용하여 PPFD가 120µmol·m-2·s-1가 되는 수관 높이(지면으로부터 평균 75cm 높이)에서 측정하였다(Fig. 1, Table 1). 모든 처리구는 농가의 요구를 반영하여 17:00부터 익일 07:00까지 14시간 점등하였다. 이에 따라 처리구에서는 일적산광량(DLI)으로 약 6mol·m-2·d-1, 대조구에서는 약 0.5mol·m-2·d-1가 인공광원에 의해 각각 추가된 것으로 볼 수 있다. 처리구별 광질 분포와 월별 DLI 수준은 Fig. 1과 Fig. 2에 각각 제시되었다.
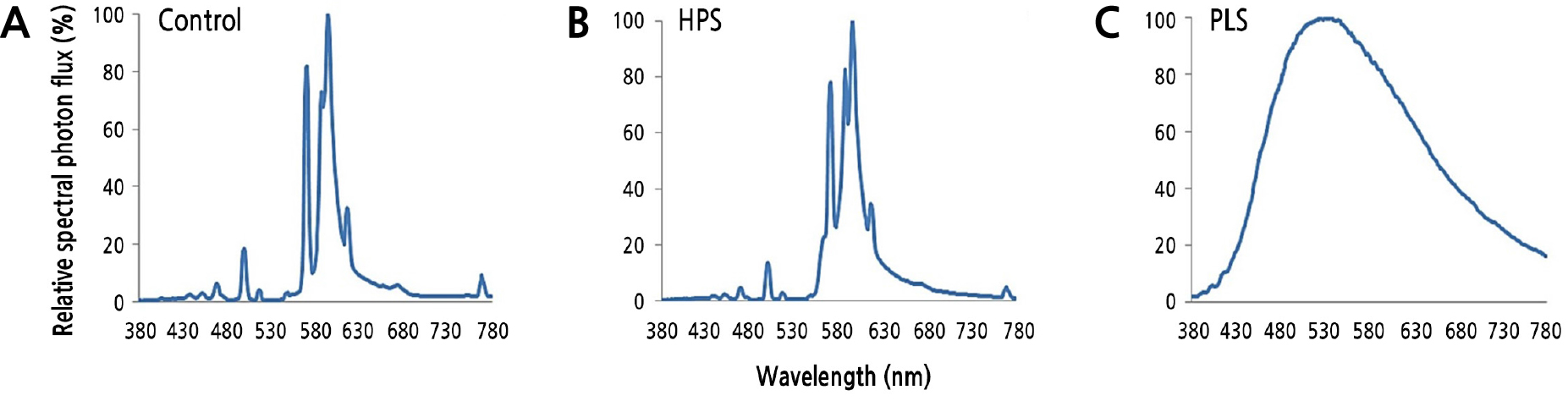
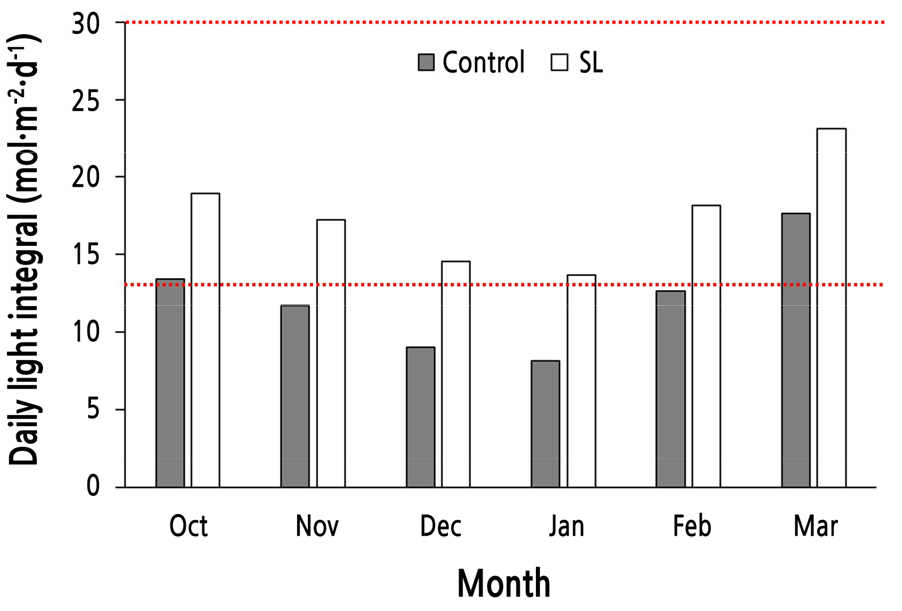
Fig. 2.
Changes in monthly daily light integral in the greenhouse without supplemental lighting (SL, control) and with SL. Dotted lines represent the minimum level (13 mol·m-2·d-1) and the optimum level (30 mol·m-2·d-1 or more) for cut rose production recommended by Dole and Warner (2017).
데이터 수집 및 통계 분석
절화 수확은 2016년 12월부터 2017년 3월까지 주당 2회 생육 관찰과 더불어 수행되었고, 생장 특성(절화장, 절간장, 엽수, 생체중, 건물중, 엽면적, 신초 발생수), 개화소요일수, 절화 특성(꽃 크기, 엽색, 절화수명), 절화 수확량 및 절화 등급별 수확량을 조사하였다. 절화 등급은 국립농산물품질관리원 농산물 표준규격(NAQS, 2019)의 스탠다드 장미 크기 구분을 따라 1급(절화장 80cm 이상), 2급(70–80cm), 3급(20–70cm)으로 구분하였다. 처리별 2반복으로 배치하였고, 각 반복에는 평균 10개체가 심겨져 있었다.
실험 자료에 대한 통계 분석을 위해 SPSS 프로그램(SPSS Statistics ver. 23, IBM, Amonk, NY, USA)을 사용하여 분산 분석(ANOVA)을 실시하였고, Tukey의 정직 유의차 검정(Tukey’s honestly significant difference test)으로 5% 유의수준에서 처리 평균 간 유의차를 검정하였다.
결 과
보광원에 따른 온실 내 광환경 차이
이 연구에 사용된 보광원인 HPS과 PLS의 분광 특성을 조사한 결과, HPS가 PLS보다 300–1,100nm 범위의 광양자속 밀도가 더 높았다(Table 1, Fig. 1). PPFD는 두 광원 모두 120µmol·m-2·s-1로 맞췄는데, HPS가 600W, PLS가 700W임을 감안하면 W당 PPFD도 HPS가 PLS보다 높다고 볼 수 있다.
Table 1.
Spectral characteristics of high-pressure sodium (HPS) and sulfur plasma (PLS) lamps at the same photosynthetic photon flux density (PPFD, 120 µmol·m-2·s-1)
PAR 범위 내에서 HPS는 PLS와 비교했을 때 청색광(B) 비중이 매우 낮은 반면, 적색광(R) 비중이 높았고, 녹색광(G)은 비슷하였다(Table 1). 이에 따라 HPS의 B:R 비율은 0.07로 매우 낮은 데 비해 PLS는 1.02로 B와 R이 비슷한 수준으로 들어 있었다.
PAR 이외의 파장 분포를 보면, 자외선(UV)은 둘다 매우 적은 수준(0.4µmol·m-2·s-1)이었고, 원적색광(FR) 또한 14.9 및 14.0µmol·m-2·s-1으로 비슷한 수준이었다(Table 1). 이에 따라 R:FR 비율은 적색광 수준이 높았던 HPS(3.85)가 PLS(2.30)보다 더 높은 수치를 보였다. 근적외선(near infra-red, NIR)은 HPS에서 46.3µmol·m-2·s-1, PLS에서 31.2µmol·m-2·s-1이 측정되어 HPS에서 상대적으로 열이 더 발생할 것으로 보였다.
광원에 따른 영양기관의 생장 및 형태적 특성
보광에 따른 절화용 장미의 생장 및 형태적 특성(Table 2)을 보면, ‘Aqua’에서는 절화장, 경경, 절간장, 엽면적 등 대부분의 특성에서 HPS 및 PLS 보광처리구가 대조구보다 높은 수치를 보였다. 광원별로는 PLS 보광처리에서 ‘Aqua’의 절화장과 절간장은 HPS와 유의차가 없었으나, 경경(6.3mm)과 엽면적(43.3cm2)은 HPS(각각 5.9mm, 38.1cm2) 처리보다 컸다(Table 2). 엽록소 함량(SPAD)은 처리 간 유의차가 없었다.
Table 2.
Growth characteristics of standard cut roses ‘Aqua’ and ‘Brut’ as influenced by different supplemental lighting sources (high-pressure sodium, HPS; sulfur plasma, PLS, lamps in a plastic greenhouse
| Light source |
Cut flower length (cm) |
Stem diameter (mm) |
No. of leaves per cut flower |
Internode length (cm) |
Leaf area (cm2/leaf) |
Chlorophyll (SPAD) |
| ‘Aqua’ | ||||||
| Control | 64.8 cz | 5.2 c | 12.1 a | 5.05 b | 32.7 c | 49.2 a |
| HPS | 68.9 b | 5.9 b | 12.4 a | 5.44 ab | 38.1 b | 50.2 a |
| PLS | 72.0 a | 6.3 a | 11.9 a | 5.80 a | 43.3 a | 49.1 a |
| ‘Brut’ | ||||||
| Control | 72.2 b | 5.7 b | 16.7 b | 4.09 b | 33.1 c | 50.4 a |
| HPS | 78.0 a | 6.4 a | 17.6 a | 4.23 ab | 37.9 b | 50.5 a |
| PLS | 79.0 a | 6.5 a | 17.3 a | 4.41 a | 41.8 a | 48.8 a |
‘Brut’에 있어서는 절화장, 경경, 엽수, 절간장, 엽면적 등의 수치가 대조구보다 HPS 및 PLS 보광처리구에서 높았다(Table 2). 절화장에서는 PLS 및 HPS 처리구에서 각각 79.0, 78.0cm로 대조구의 72.2cm보다 컸고, 경경과 엽수도 보광처리구에서 대조구보다 높은 수치를 보였다(Table 2). 절간장은 PLS 보광처리가 대조구보다 길었으며, 엽면적은 PLS > HPS > 대조구 순으로 커서 처리 간 유의차가 뚜렷하게 나타났다.
보광 처리에 따른 ‘Aqua’ 절화의 생체중과 건물중에 있어서도 전반적으로 보광 처리가 유의하게 높은 수치를 나타내었다(Table 3). 보광 광원별로는 PLS 보광이 잎, 꽃 및 전체 생체중, 그리고 줄기, 꽃 및 전체 건물중에서 HPS보다 더 높은 수치를 나타내었다. 줄기 생체중과 잎 건물중에서는 두 보광원 간의 유의차가 없었고, 꽃의 생체중과 건물중에서는 HPS 보광구와 대조구 간의 유의차가 없었다.
Table 3.
Fresh and dry weights of standard cut rose cultivars ‘Aqua’ and ‘Brut’ as influenced by different supplemental lighting sources (high-pressure sodium, HPS; sulfur plasma, PLS, lamps) in a plastic greenhouse
| Light source | Fresh weight (g) per cut flower | Dry weight (g) per cut flower | |||||||
| Leaf | Stem | Flower | Total | Leaf | Stem | Flower | Total | ||
| ‘Aqua’ | |||||||||
| Control | 12.8 cz | 11.4 b | 13.0 b | 36.6 c | 3.85 b | 4.52 c | 2.47 b | 10.84 c | |
| HPS | 15.9 b | 12.8 a | 13.3 b | 42.8 b | 4.47 ab | 5.52 b | 2.53 b | 12.52 b | |
| PLS | 19.0 a | 13.3 a | 15.8 a | 48.6 a | 5.11 a | 6.63 a | 3.01 a | 14.75 a | |
| ‘Brut’ | |||||||||
| Control | 14.4 b | 16.0 b | 11.6 b | 44.7 b | 4.51 b | 5.44 b | 2.35 b | 12.31 b | |
| HPS | 17.7 a | 21.5 a | 12.1 ab | 55.4 a | 5.59 a | 7.37 a | 2.51 ab | 15.47 a | |
| PLS | 19.3 a | 22.8 a | 13.2 a | 57.0 a | 6.23 a | 7.98 a | 2.73 a | 16.95 a | |
‘Brut’의 생체중과 건물중은 보광처리구가 대조구 대비 크게 증가한 수치를 보였으며, 부위에 따라서는 PLS가 HPS보다 조금 높은 부위도 있었으나 대체로 유의차는 없었다(Table 3). 잎 생체중은 대조구, HPS 및 PLS 보광처리구에서 각각 14.4g, 17.7g, 19.3g이었고, 줄기 생체중은 대조구, HPS 및 PLS 보광처리구 각각 16.0g, 21.5g, 22.8g이었는데, 대조구보다 HPS와 PLS 처리에서 생체중은 더 컸지만, HPS와 PLS 처리 간에는 차이가 없었다. 건물중에서도 이와 유사한 경향을 보였다(Table 3).
보광원에 따른 1일 건물 축적률(일평균 건물중 증가율)은 두 품종 모두 대조구보다 보광처리구에서 높았는데(Fig. 3A and 3B), ‘Aqua’는 대조구 대비 HPS는 33.3%, PLS는 54.2%를 증가시켰으며(Fig. 3A), ‘Brut’는 각각 58.3%, 62.5%를 증가시켰다(Fig. 3B). 이것은 보광에 의해 평균적으로 광합성량의 증가가 있었고 이것이 절화의 무게 증가로 이어졌다는 의미이다. 보광원별 1일 건물 축적률은 ‘Aqua’에서는 PLS 하에서 HPS보다 1.16배 높았으나, ‘Brut’에서는 보광원 간 차이가 없었다(Fig. 3B).
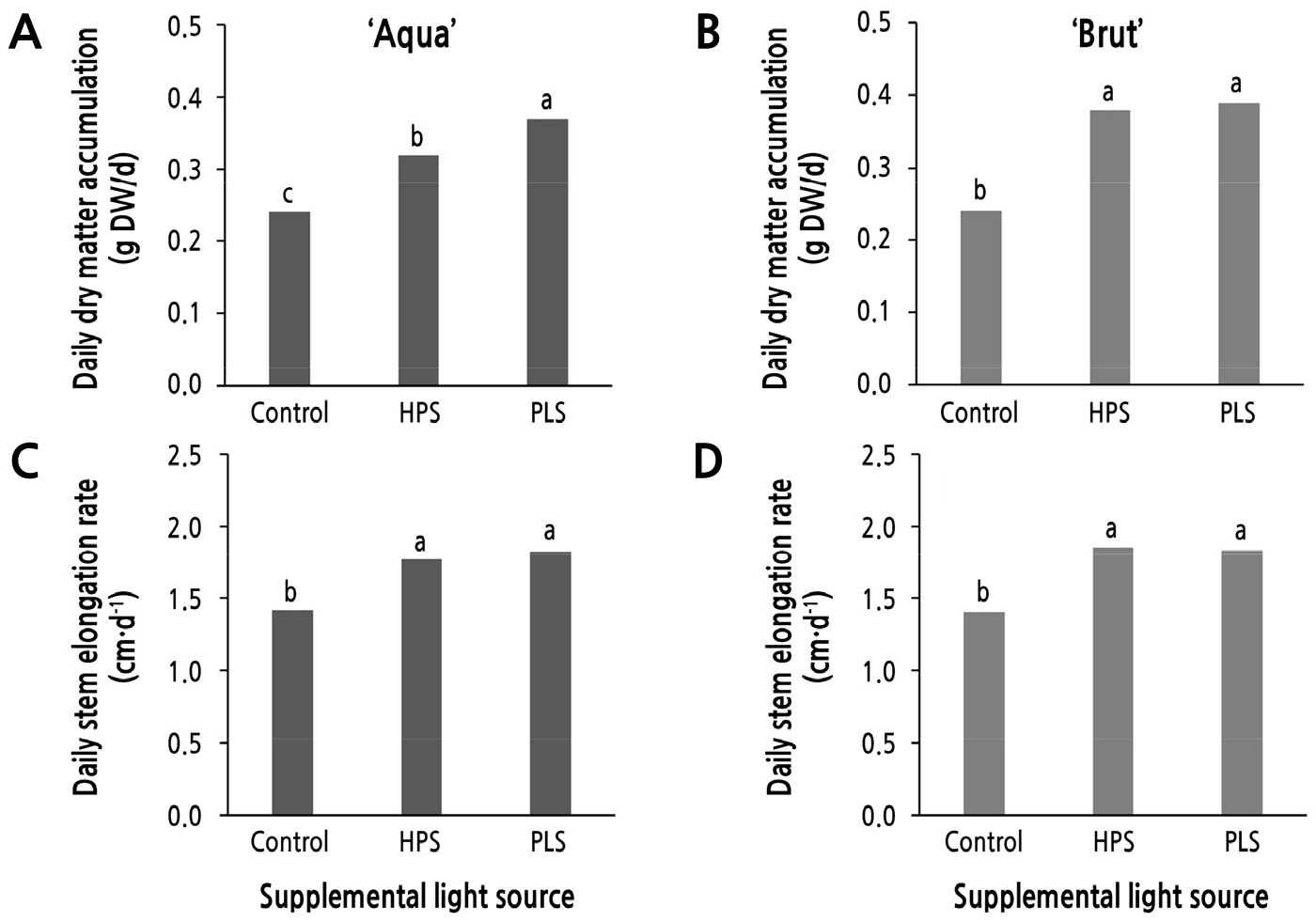
Fig. 3.
Daily dry matter accumulation rate (A, B) and daily stem elongation rate (C, D) of cut flowers in standard rose cultivars ‘Aqua’ (A, C) and ‘Brut’ (B, D) as influenced by different supplemental lighting sources (high-pressure sodium, HPS; sulfur plasma, PLS lamps) in a plastic greenhouse. Within-graph means followed by the same letter are not significantly different by Tukey’s honest significant difference test at p ≤ 0.05.
보광원에 따른 1일 줄기 신장률 역시 보광 처리구에서 대조구보다 높았는데(Fig. 3C and 3D), 두 품종 모두 보광원 간 유의차는 없었다. ‘Aqua’는 대조구 대비 HPS는 24.6%, PLS는 28.9%를 증가시켰으며(Fig. 3C), ‘Brut’는 각각 32.1%, 31.4%를 증가시켰다(Fig. 3D). 보광원별 1일 건물 축적률은 두 품종 모두 HPS와 PLS 간 유의차가 없었다(Fig. 3B).
보광원에 따른 개화 특성, 절화 품질 및 수량
보광에 따른 절화용 장미 ‘Aqua’의 개화 및 품질 특성(Table 4)을 보면, 보광처리는 대조구 대비 개화소요일수를 6.2–6.7일 감소시켰으나, PLS와 HPS 간에는 차이가 없었다. 화고와 절화수명에서는 PLS 처리가 대조구 및 HPS에 비해 증가되었다. 반면 꽃잎수와 화폭에 있어서는 처리 간 차이가 없었다.
Table 4.
Flower quality characteristics and daily dry matter accumulation of standard cut rose cultivars ‘Aqua’ and ‘Brut’ as influenced by different supplemental lighting sources (high-pressure sodium, HPS; sulfur plasma, PLS, lamps) in a plastic greenhouse
| Light source | Days to flower | Flower size (mm) | No. of petals per cut flower | Vase life (days) | |
| Height | Diameter | ||||
| ‘Aqua’ | |||||
| Control | 45.6 az | 44.2 b | 41.4 a | 35.7 a | 28.4 b |
| HPS | 38.9 b | 44.9 b | 38.8 a | 34.9 a | 28.8 b |
| PLS | 39.4 b | 46.5 a | 39.0 a | 36.6 a | 31.4 a |
| ‘Brut’ | |||||
| Control | 51.5 a | 32.0 a | 43.8 b | 36.8 a | 29.3 a |
| HPS | 42.1 b | 33.3 a | 44.5 ab | 35.6 a | 29.6 a |
| PLS | 43.0 b | 32.5 a | 45.3 a | 35.8 a | 30.9 a |
‘Brut’의 경우, 대조구 대비 보광 처리구의 개화소요일수는 8.5–9.4일 단축되었으나, HSP와 PLS 간에는 차이가 없었다(Table 4). 그리고 PLS 보광 처리 시 화폭이 대조구보다 증가하였다. 하지만 화고, 꽃잎수, 절화수명 등의 특성에서는 처리 간 차이가 없었다.
절화의 m2당 수량(3등급 이내)은 ‘Aqua’에서 대조구 159본, HPS 234본, PLS 240본, ‘Brut’에서 대조구 184본, HPS 243본, PLS 252본으로 두 품종 모두 보광처리구가 통계적으로 유의하게 절화 수확량을 증가시켰다(Fig. 4). 총 수량에 있어서는 HPS와 PLS의 보광처리 간 유의차는 없었다. 월별 절화 수확량에 있어서는 HPS와 PLS의 보광처리가 대조구와 비교하여 두 품종 모두 매월 모두 더 많았으나, 두 보광원 사이에서는 큰 차이가 없었다(Fig. 4A and 4B).
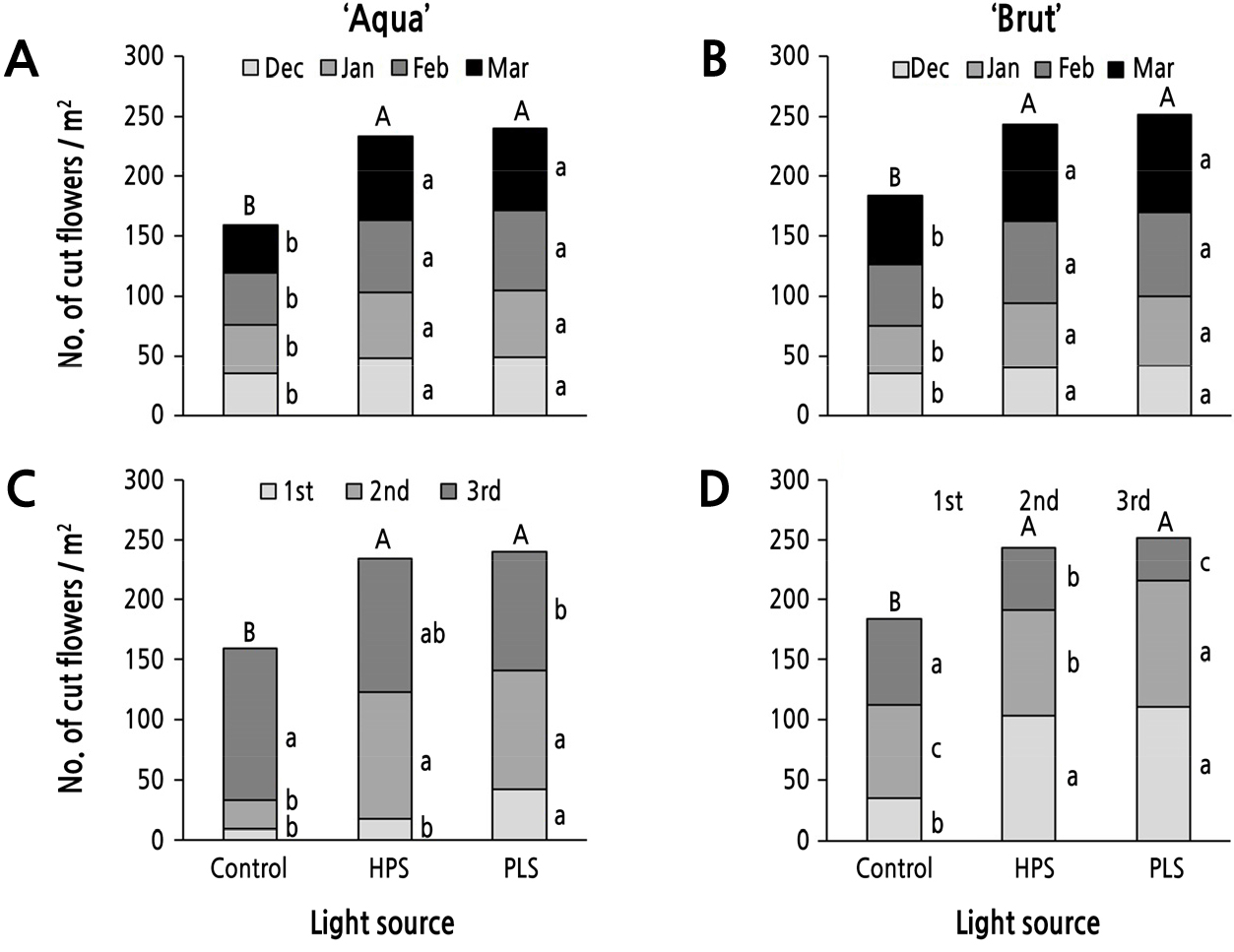
절화 등급별 수확량을 보면, ‘Aqua’에서는 PLS의 1등급(절화장 80cm 이상) 수량이 가장 많았는데, 대조구 및 HPS 대비 각각 4.67배, 2.33배 많았다(Fig. 4C). 2등급(절화장 70cm 이상–80cm 미만) 수량은 두 보광처리구에서 대조구보다 많았으나, 3등급(절화장 70cm 미만) 수량에서는 대조구가 PLS 보광구보다 많았다. ‘Brut’에서는 1등급 수량이 두 보광처리구에서 대조구보다 많았고, 보광처리 간에는 유의차가 없었다(Fig. 4D). 2등급 수량은 PLS > HPS > 대조구 순으로 많았던 반면, 3등급은 그 역순으로 많았다. 절화 등급은 절화장을 기준으로 한 것이므로 절화장의 처리별 순서(Table 2)와 관계가 있었다.
고 찰
이 연구에 사용된 PLS는 관행의 보광원인 HPS에 비해 300–1,100nm 범위의 광양자속 밀도가 조금 낮았고, W당 PPFD도 HPS보다 조금 낮은 수준이었는데(Table 1, Fig. 1), 이는 Park et al.(2018b)의 보고와 유사하였다. 또한 HPS가 완성된 온실 보광 전용 인공광원인 데 비해, PLS는 아직 개발 중이라 아직 반사판 등 온실 보광용으로써 개선의 여지가 있다고 판단된다. PAR 범위 내에서 PLS는 HPS와 비교했을 때 청색광(B) 비중이 매우 높은 반면, 적색광(R) 비중이 낮았고, 녹색광(G)은 비슷하였다(Table 1). 이에 따라 HPS의 B:R 비율은 0.07로 매우 낮은 데 비해 PLS는 1.02로 B와 R이 비슷한 수준으로 들어 있었다.
PAR 이외의 파장에서는, UV와 FR은 두 광원에서 유사하였으나 NIR은 HPS(46.3)에서 PLS(31.2)보다 많이 방출되었다(Table 1). 이에 따라 R:FR은 R 수준이 높았던 HPS(3.85)가 PLS(2.30)보다 상대적으로 더 높아 광형태형성에서 두 광원이 다른 효과를 보일 가능성이 있다(Smith, 1982; Park et al., 2016). NIR이 많은 HPS는 상대적으로 난방 보조 효과가 있으며 엽온을 더 높여 형태적 및 생리적 차이를 가져올 가능성이 있다(Oh et al., 2010; Guo et al., 2016).
두 광원을 비교하면, HPS는 상대적으로 B가 매우 적고 R이 많아 B:R이 0.07로 매우 낮은 반면, G는 R과 유사한 수준으로 분포하였으며, R이 FR보다 많아 R:FR가 상대적으로 높은 3.58이었다. 반대로 PLS는 B와 R이 유사한 수순으로 분포하여 B:R은 태양광과 유사한 1.02이고, FR은 HPS와 유사한 수준이어서 R:FR가 2.30이었는데, 태양광의 1.09(Smith, 1982)보다는 약 2배 높았다. 이렇게 두 광원은 B와 R의 양과 비율(B:R), 그리고 R:FR에서 차이가 있어서 온실에서 자연광을 보조하는 조건에서도 바질(Ocimum basilicum L.; Dörra et al., 2020), 미니 오이(Guo et al., 2016), 오이와 토마토(Hogewoning et al., 2012), 파프리카(Lee et al., 2014; Park et al., 2018b) 등에서와 같이 차이가 나타날 것으로 보인다.
장미는 일장이 개화에 영향을 주지 않는 중성식물(day neutral plant)이면서 광요구도가 높은 대표적인 호광성 작물로 일적산광량(DLI)이 절화의 수량과 품질에 직접적으로 영향을 준다(Dole and Wilkins, 2005; Dole and Warner, 2017). 위도가 높은 지역에서의 절화 장미 생산 시 약광기에 수량과 품질을 향상시키기 위해 보광을 실시하는데, HPS 램프를 이용하여 8–24시간 동안 60–200µmol·m-2·s-1의 인공광을 조사하는 것이 일반적이다(Dole and Warner, 2017). 보광의 효과는 광도와 조사시간, 자연광량 등에 따라 달라질 수 있는데(Lee and Kim, 2015), 보광 시간은 계절, 보광등의 광도, 전기료 등을 고려하여 결정한다. 절화 장미 재배 시 권장되는 광량은 DLI 기준으로 최저 13mol·m-2·d-1, 최고 30mol·m-2·d-1 이상(Fig. 2)으로 다른 절화류에 비해 높은 편이다(Dole and Warner, 2017).
이 연구에서는 일반적인 보광원인 HPS와 신광원인 PLS가 사용되었는데, 일부 생장 및품질 특성에서 차이는 있었으나 전반적으로 보광 처리가 신초의 생장, 절화 품질, 개화소요일수, 절화수명, 총 수량 및 절화 등급(절화장)에서 대조구와 비교하여 좋은 결과를 보였다(Tables 2, 3, 4, Figs. 3 and 4). 이는 보광 처리구의 DLI가 대조구보다 5.5mol·m-2·d-1 더 많이 공급된 것에 기인한다고 할 수 있다. Choi et al.(2008)은 장미 ‘Vital’에 대한 보광은 광합성 촉진과 탄수화물의 축적 및 전류를 활성화하여 생육을 촉진시키고, C/N율이나 호르몬 등에 영향을 주어 결과적으로 개화소요일수를 단축시켰다고 보고하였다. Armitage and Tsujita(1979)와 Bredmose(1993)는 보광이 장미의 줄기 직경을 증가시켰다고 보고하였다. Marissen(2001)은 절화 장미 재배 시 보광이 체내 탄수화물의 증가 등으로 절화수명을 증가시켰다고 하였다. 또한 보광 처리 시 광합성의 증가에 의해 개화가 촉진되는 부분도 있지만, 야간에 인공광 조사에 따른 엽온 및 생장점 온도 상승이 발육을 촉진한 것(Faust and Heins, 1997; Oh et al., 2010; Kwon et al., 2014)으로 추정할 수도 있다.
보광은 보통 절화 장미의 줄기 신장을 촉진시키므로 절화장이나 절간장을 증가시키며(Armitage and Tsujita, 1979), HPS 보광이 절화장 등급이 높은 절화의 비율을 증가시켰다(Farina and Veruggio, 1996). 품종에 따라 다른 결과가 나올 수는 있으나(RDA, 2018), 이 연구에서 사용된 두 품종은 보광이 줄기 신장을 촉진시킨다고 볼 수 있다.
이 연구의 결과와 이전의 보고들을 종합해 볼 때, 겨울철 약광기에 보광으로 증가된 DLI가 광합성을 촉진하여 생장 속도를 증가시키고 이로 인해 작기의 단축, 생산량 및 품질 향상으로 이어졌다고 볼 수 있다(Fig. 5A). 또한 인공광 조사는 잎이나 생장점의 온도 상승을 유발하며 온실 내부의 온도도 증가시키는데 이로 인해 식물체의 발육 속도가 증가되므로 개화소요일수가 짧아지고 난방비 절감을 기대할 수 있다(Fig. 5A). 보광은 식물체의 잎이나 정단부의 체온 상승을 동반하므로 발육 속도 역시 증가하였고 개화도 촉진되었다고 볼 수 있다.
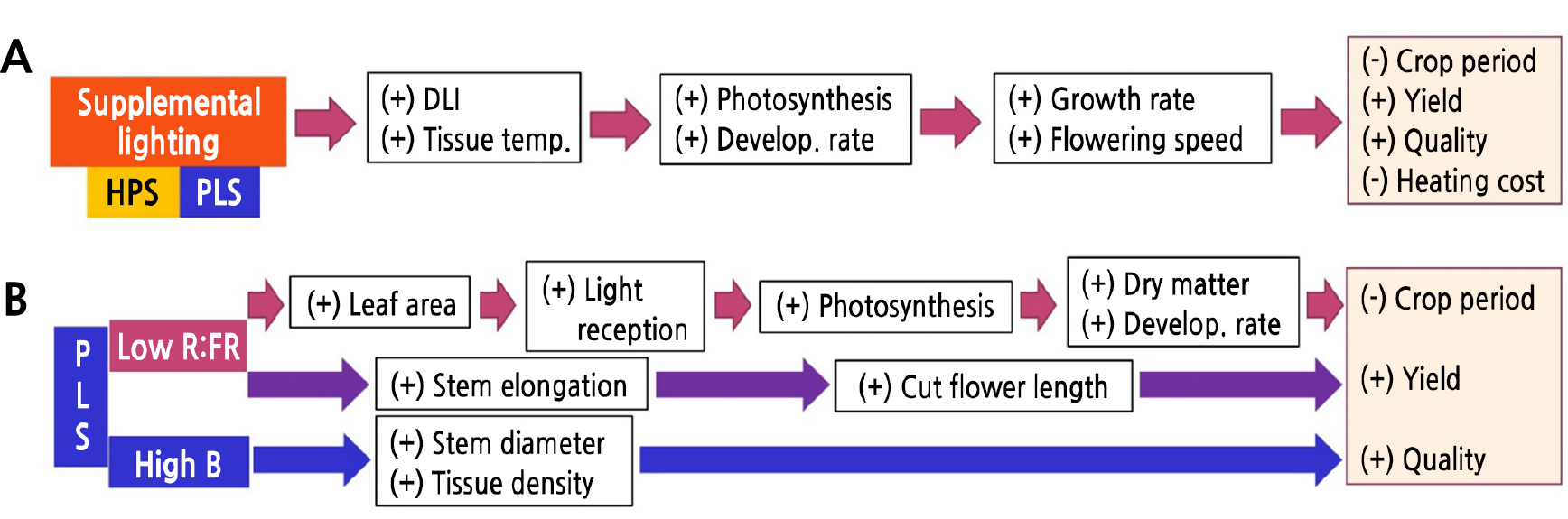
두 보광원, 즉 HPS와 PLS는 이 실험에서 120µmol·m-2·s-1의 같은 PPFD로 보광처리가 되었지만, PLS가 HPS보다 상대적으로 B가 많고 R이 적어 B:R이 높고, R이 적고 FR은 비슷하여 R:FR이 낮았다(Table 1). 그리고 HPS는 NIR을 더 많이 방출하였다. 이러한 차이는 장미 절화에 생육 특성은 물론 결과적으로 수량과 품질의 차이(Tables 2, 3, 4, Figs. 3 and 4)를 가져왔다고 볼 수 있다. ‘Aqua’에서는 절화장, 경경, 엽면적, 생체중(줄기 제외), 건물중(잎 제외), 화고, 절화수명, 일일 건물 축적률, 1등급 수량 등에서 PLS가 HPS보다 상대적으로 우수한 수치를 보였다.
우선 줄기 신장 및 잎의 팽창의 결과로 나타나는 절화장과 엽면적은 PLS에서 HPS보다 더 컸는데, 이는 PLS가 HPS보다 낮은 R:FR 비율을 갖고 있어 줄기 신장 및 잎의 팽창을 촉진시켰기 때문이라 판단된다. R:FR이 낮은 환경에서는 광요구도가 높은 양지식물의 절간 신장 증가, 엽면적 증가 등의 피음반응(shade avoidance response)이 발생한다(Halliday et al., 1994; Smith and Whitelam, 1997; Franklin, 2008). R:FR 비율이 낮을수록 초장과 엽면적이 커지는 현상은 다양한 식물에서 나타난다(Runkle et al., 2012; Oh and Runkle, 2016; Park et al., 2016; Hernández and Kubota, 2017).
특히 PLS와 HPS의 차이를 비교한 연구들도 다수 존재한다. Dörra et al.(2020)은 PLS 보광이 바질(Ocimum basilicum L.)의 초장과 절간장을 HPS보다 각각 11%, 27% 증가하였는데 이는 PLS의 R:FR가 1.66으로 HPS의 2.90보다 낮기 때문이라 보고하였다. Hogewoning et al.(2012)은 보광용으로 조사된 PLS가 HPS보다 오이의 초장, 엽병장을 더 증가시켰고, 토마토의 초장도 증가시켰는데, 이는 PLS의 R:FR가 HPS보다 낮기 때문이라 하였다. R:FR 값이 낮아서 phytochrome photostationary state[Pfr/(Pr+Pfr), PSS]가 낮은 인공태양(AS)이나 HPS+FR에서는 HPS에 비하여 공통적으로 줄기와 엽병의 신장이 촉진되었고 엽면적이 증가하였다(Kang et al., 2019). Lee et al.(2014)은 자연광에 가장 근접한 파장을 방출하는 것으로 알려져 있는 플라즈마 광원인 lighting-emitting plasma(LEP) 램프가 HPS에 비해 파프리카의 엽면적을 더 증가시켰다고 보고하였으며, Park et al.(2018b)은 PLS 보광 하에서 파프리카의 초장과 엽면적이 HSP보다 더 컸으며, Park et al.(2016)도 R:FR이 낮은 광원이 피튜니아의 초장을 증가시켰다고 하였다.
이들 연구 결과에서 많은 경우 초장과 엽면적의 증가가 생체중 및 건물중의 증가로 이어졌는데, 이는 수광량의 증가에 의한 광합성의 증가로 볼 수 있다. Hogewoning et al.(2010)은 낮은 PSS 값에서 나타나는 엽병의 길이 증가와 엽면적의 증가는 작물의 수광 면적을 증가시키며, 이로 인해 R:FR이 낮은 AS와 HPS+FR에서 작물 전체의 생육이 높게 나타난 것으로 유추한다고 하였다. Lee et al.(2014)은 자연광에 가장 근접한 파장을 방출하는 것으로 알려져 있는 플라즈마 광원인 lighting-emitting plasma(LEP) 램프가 HPS에 비해 파프리카의 엽면적과 더불어 건물중도 더 증가시켰고, Park et al.(2018b)은 PLS 보광 하에서 파프리카의 초장, 엽면적 외에도 생체중 및 건물중이 HSP에 비해 더 증가하였다고 보고하였다. 즉 낮은 R:FR는 식물의 줄기(절간) 및 엽병 신장과 잎의 팽창을 촉진하여 광원과 더 가까워지면서 수광 면적도 증가시키기 때문에 이로 인한 광합성의 증가로 이어진다고 볼 수 있다. 그 일일 줄기 신장률이나 건물 축적률이 증가하였고, 개화소요일수의 감소와 절화수명 증가에도 영향을 주었으며, 작기 단축, 수확량 증가, 품질 향상으로 이어졌다고 추론할 수 있다(Fig. 5B).
‘Aqua’에 비해 ‘Brut’에서는 엽면적과 2등품 수량에서만 PLS가 HPS보다 통계적으로 유의하게 높은 수치를 보였고, 다른 항목에서는 차이가 나타나지 않았다. 이는 PPFD가 같을 경우 이 연구에 사용된 두 광원 간 광질의 차이가 품종에 따라서는 그 효과가 다르게 나타날 수 있음을 보여준다. 같은 광원에 의한 보광 시에도 보광의 효과가 품종에 따라 조금 다르게 나타난 사례도 있었다(RDA, 2018).
한편, PLS는 HPS에 비해 청색광 비율도 높은데, 이 특성도 작물 생육에 영향을 주었을 것으로 생각된다. 청색광은 기공 개폐에 영향을 주는데, 청색광 조사는 잠두(Vicia faba)의 기공을 열며, 녹색광을 잠시 조사하면 기공이 닫혔다가 다시 청색광을 재조사하면 기공이 열린다(Assmann and Shimazaki, 1999). Park et al.(2018b)은 PLS 보광 하에서 파프리카의 초장, 엽면적, 생체중 및 건물중이 HSP보다 더 컸으며, 이는 PLS 하에서 증가된 기공전도도, 증산률, 광합성 등에 기인한다고 하였다. Eun et al.(2011)은 들깨와 고추에서 지상부의 생체중과 건물중 향상에 청색광이 좋았다고 하였으며, Kim et al.(2013)도 청색광 하에서 임파첸스의 생체중과 건물중이 증가하였다고 보고하였다. 이로부터 청색광이 상대적으로 많은 PLS가 B:R이 낮은 HPS와 비교할 때, 기공을 더 열게 하여 기공전도도를 높이고 증산률을 높였으며 이로 인해 광합성이 촉진되었을 것으로 추론할 수 있다(Fig. 5B).
또한 청색광은 줄기 신장을 억제하고 조직을 치밀하게 하는 경향도 있다(Warpeha and Kaufman, 1989; Shimizu et al., 2005). PLS에서 높은 청색광의 비율로 HPS에 비해 광합성 및 줄기의 부피 생장이 증가하였고 조직이 치밀해져 절화수명 연장에 영향을 주었다고 생각된다. 반면 낮 동안 청색광(B) 비율을 줄인 고광도의 빛에 대한 절화 장미 ‘Mercedes’ 식물체의 반응은 줄기 길이와 건물중의 증가였다(Maas and Bakx, 1995). 다만 이 연구는 생장상에서 형광등과 필터를 이용하여 몇 가지 파장을 만들어낸 것이라 이 연구와는 조건이 많이 달라 직접 비교할 수는 없다. 그리고 이 연구에서는 PLS가 청색광이 상대적으로 많으면서도 R:FR이 낮기 때문에 청색광에 의한 줄기 신장 감소가 나타나지 않은 것으로 보인다.
한편, PLS의 파장 분포는 500–600nm 범위에서 균일하게 분포하는 데 비해, HPS는 560–590nm 사이에 피크가 있으며 800nm 이상의 적외선을 많이 방출하는데, 이런 HPS의 파장 분포가 엽온을 더 증가시킨다고 볼 수 있다(Guo et al., 2016). 또한 Guo et al.(2016)는 HPS 하에서 자란 미니 오이의 제5엽의 면적은 플라즈마 램프 하에서 자란 식물체에 비해 유의하게 작았는데 이는 높은 엽온에 기인한다고 추론하였다. 이 연구에서도 HPS에서 NIR이 상대적으로 많아 PLS보다 엽온이 더 증가함에도 불구하고, 기공 폐쇄로 기공전도도와 증산이 감소하면서 엽온을 낮추지 못해 광합성 감소와 호흡 증가로 인한 건물 축적 감소로 이어졌을 수도 있다. 그러나 엽온의 수준이 식물에 억제적인 작용을 할 정도로 높지 않을 경우 오히려 광합성을 증가시키고 발육을 촉진할 수도 있다. PLS가 더 좋은 특성을 가지고 있음에도 항목에 따라 그리고 품종에 따라 많은 특성에서 큰 차이가 나지 것이 이와 관련이 있을 것으로 보인다. 이에 대해서는 보광원에 따라 엽온을 비교하는 실험이 추가로 수행되어야 할 것이다.
결론적으로, 절화 장미 두 품종 모두 보광에 의해 절화의 수량과 품질이 향상되는 결과를 보였으며, 특히 ‘Aqua’ 품종에서는 PLS의 보광 효과가 HPS보다 더 높게 나왔다. 이 결과는 PLS가 가진 파장 특성과 관련이 있는 것으로 보이는데, HPS와 비교했을 때 높은 청색광 비율과 낮은 R:FR 비율이 영향을 준 것으로 판단된다. 그리고 PLS는 관행의 HPS에 비해 효과적인 파장 특성을 가지고 있다고 판단되는데, 온실 보광 실험 시 일부 특성에서 통계적으로 향상된 결과를 보이지 못하였다. 그 이유 중 하나는 품종 간 보광 효과의 차이 때문이며, 다른 하나는 아직 개발 중인 PLS가 광합성유효파장(PAR)의 발광 효율 향상, 작물 수광에 적합한 반사판 설계 등의 후속 개발이 필요하기 때문이라 생각된다. 앞으로 성능이 향상된 PLS가 출시되면 품종을 추가하고 면적을 넓혀 실증 실험을 추가로 수행할 필요가 있다.
