서 언
재료 및 방법
실험재료
숙기조사
통계분석
결과 및 고찰
교배친 품종의 숙기 변이
숙기의 유전력
교배실생의 숙기 분포
숙기의 양친 평균과 교배실생 평균과의 상관
숙기와 과중간의 상관
숙기 관련형질의 우량 개체 출현 빈도의 산출
서 언
우리나라 배 재배면적은 1990년에 9천ha에서 2000년에 26천ha로 증가하였다가 2013년에는 14천ha로 감소되었다(KOSIS, 2013). 이러한 재배면적 감소 원인으로는 만생종 ‘신고’ 품종이 2013년 82.9%로 편중 재배되고 있어(KREI, 2013) 추석이 이른 해에 ‘신고’ 미숙 과실이 출하되어 소비자들에게 배는 맛없는 과실로 인식되어 소비가 감소되기 때문이다. 따라서 소비자와 생산자의 요구를 모두 충족시킬 수 있는 수확시기가 다양한 배 품종의 개발과 보급이 시급한 실정이다.
배는 자가불화합성으로 유전적으로 이형접합 비율이 높고, 일년생 작물에 비해 한 세대의 길이가 길고 각 개체의 크기가 커서 육종을 위해서는 많은 포장면적과 경비가 소요되어 육종효율이 낮은 반면 우량 형질을 보유한 개체가 일단 선발 되면 균일성(homogeneity)에 관계없이 영양번식으로 대량 급속 보급이 가능한 장점이 있다(Kim, 1985; Shin et al., 1983). 따라서 과수에서는 육종효율을 높이기 위하여 교배실생의 유년기간(juvenile period)을 단축하기 위한 결실 촉진, 유묘기에 목표형질을 가진 개체를 선발할 수 있는 생화학적 조기선발 표지의 개발, 주요 병의 유묘기 접종을 통한 저항성 개체의 조기선발, 그리고 잎과 과실과의 유전상관을 이용한 조기선발 지표의 개발 등의 방법이 시도되고 있으며, 한편으로는 주요 과실형질의 유전현상을 파악하여 우수한 실생 비율이 높은 교배조합의 선정, 교배조합당 필요한 실생 개체수의 예측과 같은 유전연구를 통하여 육종효율을 높여 나가야 한다(Hong, 1991).
배 과실의 숙기 유전 연구에서 과실 성숙의 조만은 양적형질로서 교배조합별 실생의 숙기 분포는 정규분포를 보이며 다수유전자(polygene)에 의해 지배된다고 하였다(Abe et al., 1993a). 숙기의 유전력은 매우 높아 환경의 영향이 거의 무시될 정도로 작용한다고 하였다(Hansche et al., 1972a, 1972b; Shin et al., 1983). 또한 숙기는 유전력이 높으므로 실생개체의 선발이 1-2년의 단기간에 행해질 수 있으며 양친의 평균숙기를 육종목표로 하는 숙기와 거의 일치되도록 교배조합을 선정함으로써 기대할 수 있는 개체를 가장 효율적으로 획득할 수 있다고 하였다.
Yamaguchi et al.(1984)은 복숭아 교배실생의 숙기분포에 대하여 만생종 품종간의 조합에서는 실생의 숙기가 만생부근에 집중되며, 만생×조생 조합에서는 실생 숙기가 조생과 만생으로 분리되어 2개의 peak를 나타낸다고 하였다. 또, 숙기의 조만성은 주동유전자(major gene)의 지배를 받는 것으로 추정하였으며, 이들 유전자와는 별도로 다수유전자 및 비교적 효과가 작은 동의인자(multiple gene)도 숙기 발현에 관여하는 것으로 생각된다고 하였다.
또 Hayashi(1953)는 배 조생종×조생종 조합에서 실생의 숙기 평균은 양친보다 늦어지는 방향으로, 중생종×중생종 조합에서는 중생 및 만생으로, 만생종×만생종 조합에서는 조생 방향으로 분포됨을 보고하였다.
Kim(1985)은 배 과실 성숙기는 조생이 만생에 대해 우성으로 나타났으며, 양친의 평균 성숙기와 실생평균을 비교하면 실생평균이 조합에 따라 7-18일 더 빨랐다고 하였다. 또 Shin et al.(1983)은 실생의 숙기가 양친의 중간에 분포하는 것이 많았다고 하였다.
본 연구에서는 배 과실의 숙기형질에 대하여 교배양친의 연차별 환경변이 정도를 파악하고, 숙기에 대한 각 교배조합별 유전력, 후대 실생의 분포상태, 그리고 양친의 평균치와 교배실생 평균치와의 상관을 분석하였다. 또한 육종 계획 수립시 육종 목표로하는 형질을 가진 실생 비율을 높일 수 있는 교배친의 선정기준과 조합당 육성할 개체수를 결정하는 데 필요한 기초자료로서 양친평균에 따른 후대 교배실생의 우수개체 출현비율을 산출하여 교배조합을 선정할 때의 적용 가능성을 고찰하였다.
재료 및 방법
실험재료
본 연구에 공시된 재료는 국립원예특작과학원 과수과에서 배 양질 다수 내재해성 품종 육성을 위하여 1986년에서 1990년까지 13품종을 교배친으로 이용하여 얻은 실생 15조합 4,035주를 대상으로 하였다. 본 연구에서 사용된 교배친 품종의 육성내역을 보면 일본에서 육성된 9품종과 우리나라에서 육성한 3품종, 그리고 우리나라 수집 재래배 1품종 등 13품종이었다(Table 1).
과실형질 조사에 이용된 교배조합별 실생수, 교배년도는 Table 2와 같다. 수령은 6-9년생이고 재식거리는 4.0-5.0m × 0.5m였으며 수형은 주간형으로 하였다. 시비, 병해충 방제 등 재배관리는 국립원예특작과학원 표준경종법에 준하였다.
Table 1. Parental cultivars used in this experiment. |
Cultivar | Cross combination | Origin | Released year |
Imamuraaki | Chance seedling | Japan | 1872 |
Okusankichi | Chance seedling of Wasesankitsu | Japan | 1877 |
Chojuro | Chance seedling | Japan | 1893 |
Niitaka | Amanogawa × Imamuraaki | Japan | 1927 |
Kimizukawase | Shinkozo × Doitsu | Japan | 1940 |
Kosui | Kikusui × Wasekozo | Japan | 1962 |
Shinsui | Kikusui × Kimizukawase | Japan | 1965 |
Tama | Gion × Kosui | Japan | 1971 |
Choju | Asahi × Nijisseiki | Japan | 1973 |
Chuwhangbae | Imamuraaki × Nijisseiki | Korea | 1985 |
Soowhangbae | Chojuro × Kimizukawase | Korea | 1988 |
Whasan | Hosui × Okusankichi | Korea | 1992 |
Satangbae | Collected native cultivar | Korea | - |
Table 2. Cross combination and seedlings used in this experiment. |
Cross combination | Cross year | No. of seedling |
Niitaka × Choju | 1988 | 382 |
Niitaka × Tama | 1988 | 449 |
Niitaka × Chuwhangbae | 1988 | 381 |
Niitaka × Soowhangbae | 1988 | 206 |
Niitaka × Satangbae | 1986 | 116 |
Niitaka × Shinsui | 1990 | 517 |
Shinsui × Soowhangbae | 1989 | 97 |
Kosui × Soowhangbae | 1989 | 136 |
Kimizukawase × Soowhangbae | 1989 | 251 |
Imamuraaki × Soowhangbae | 1989 | 168 |
Okusankichi × Soowhangbae | 1989 | 98 |
Chuwhangbae × Soowhangbae | 1989 | 60 |
Chojuro × Soowhangbae | 1989 | 617 |
Shinsui × Whasan | 1989 | 440 |
Kosui × Whasan | 1989 | 117 |
Total 15 | - | 4,035 |
숙기조사
교배친 13품종의 과실 형질은 배 유전자원 보존 포장에 6 × 6m로 재식된 품종당 3-5주의 성목에서 1987-1996년까지 10년간 조사하였고, 교배실생에서는 1994-1996년에 걸쳐 2-3년간 조사하였으며, 연도별 조사 성적의 차이가 큰 일부 개체는 조사상의 오류로 판단하여 분석대상에서 제외하였다. 각 형질별 조사는 농촌진흥청에서 정한 ‘농사시험연구조사기준(1995)’을 참고하여 조사하였다.
숙기는 전 과실의 70-80%가 성숙한 시기를 순별로 표시하였다. 성숙기준은 먼저 황갈색배의 경우는 과피색이 녹갈색에서 갈-황갈색으로 변하고 바탕색이 황갈-갈색으로 변하는 시기로 하였고, 녹황색배의 경우는 녹색에서 황록색으로 변한 때로 하였다. 수확 시 손가락으로 가볍게 눌러 보아 과육이 움푹 들어가는 과실을 1-3회에 걸쳐 수확하여 과피색 외에도 당도, 경도, 맛, 향기 등을 고려하여 숙기를 정한 후 순별로 표시하였다. 숙기는 8월 상순부터 10일 간격으로 11월 상순까지 10등급으로 구분하여 실생의 분포 비율을 조사하였다.
통계분석
본 시험에 공시한 교배실생 4,035주와 교배친 13품종의 각 숙기 조사성적은 Excel 2000을 이용하여 자료를 정리하였다. 유전분석, 분산, 실생분포도 등은 Excel을 이용하여 분석하였고, 형질간 상관분석은 SAS프로그램(Version 6.03, SAS Institute Inc., 1991, Cary, NC)을 이용하였다. 각 형질의 유전력 분석은 Hong(1991)과 Park(1987)의 방법에 따라 양친 품종 P1과 P2의 분산평균과 교배실생의 분산을 산출하였고, 유전력은 다음의 공식을 이용하여 계산하였다.
분 산(S2) = 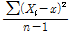 =
= 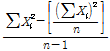
표준편차(S) = 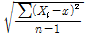
변이계수(C.V.) = 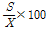
유전력(h2) = 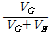
유전분산(VG) = 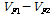
환경분산(VE) = 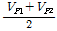
결과 및 고찰
교배친 품종의 숙기 변이
1987년부터 1996년까지 10년에 걸쳐 조사된 교배 양친 품종의 숙기 평균과 표준편차 및 변이계수는 Table 3과 같다. 평균 숙기는 ‘장수’가 8월 16일로 가장 빠르고 ‘만삼길’이 11월 6일로 가장 늦다. 이들의 표준편차를 보면 ‘군총조생’, ‘신수’, ‘사탕배’, ‘장수’ 등 조생 품종들은 연차간 변이가 6.27-7.89로 중∙만생종에 비해 큰 것이 많았다. 또 연도별 숙기 차이는 가장 빠른 해의 수확일과 가장 늦은 해의 수확일 사이에 ‘사탕배’, ‘신수’, ‘수진조생’ 등은 23일 차이를 나타냈고, ‘장수’, ‘금촌추’ 등이 22일이었으며, ‘만삼길’은 9일로 가장 차이가 적었다. 만생종 ‘금촌추’는 떫은 맛의 유무가 수확기 판정에 중요한 요소로 해에 따른 수확기 온도차이가 다른 품종에 비해 수확기 결정에 더 큰 영향을 준 것으로 판단된다. 변이계수에 있어서도 비슷한 경향으로 ‘장수’, ‘사탕배’, ‘신수’, ‘군총조생’ 순으로 변이계수가 큰 경향을 보여 대부분의 조생종의 숙기가 만생종에 비해 숙기변이가 큰 것으로 나타났다. 그러나 모든 품종의 숙기 변이계수는 0.92-3.41 사이로 과실의 다른 형질에 비해 변이 폭이 좁고 안정된 형질임을 보여주었다.
Table 3. Yearly variations of parental cultivars on fruit ripening date (1987-1996). |
Cultivar | Ripening date | Meanz | C.V. |
First | Last |
Kimizukawase | Aug. 7 | Aug.25 | Aug.17.1 ± 6.27 | 2.79 |
Satangbae | Aug. 2 | Aug.25 | Aug.18.8 ± 7.72 | 3.41 |
Shinsui | Aug. 2 | Aug.25 | Aug.20.2 ± 6.81 | 2.98 |
Choju | Aug. 4 | Aug.26 | Aug.16.4 ± 7.89 | 3.52 |
Kosui | Aug.28 | Sept.10 | Sept. 2.6 ± 3.34 | 1.33 |
Tama | Aug.13 | Sept. 1 | Aug.22.2 ± 4.82 | 2.10 |
Soowhangbae | Sept.22 | Oct. 8 | Sept.28.3 ± 6.75 | 2.54 |
Chojuro | Sept.12 | Sept.25 | Sept.20.1 ± 4.38 | 1.70 |
Niitaka | Oct. 1 | Oct.15 | Oct.08.1 ± 5.49 | 1.99 |
Chuwhangbae | Oct.15 | Oct.30 | Oct.22.3 ± 6.09 | 2.11 |
Imamuraaki | Oct.24 | Nov.10 | Nov. 2.3 ± 5.46 | 1.74 |
Okusankichi | Nov. 1 | Nov.10 | Nov. 7.7 ± 2.79 | 0.92 |
Whasan | Sept.29 | Oct.18 | Oct. 6.6 ± 6.41 | 2.34 |
Mean | - | - | Sept.19.3 ± 5.68 | 2.27 |
zMean ± standard deviation. |
숙기의 유전력
배 15개의 교배조합에서 양친 평균 숙기가 가장 빠른 조합은 ‘군총조생’ × ‘수황배’로서 9월 8일이었고, 가장 늦은 조합은 ‘만삼길’ × ‘수황배’로 10월 17일이었다(Table 4). 교배실생의 숙기 평균도 비슷한 경향으로서 9월 5일-10월 13일이었다. 유전력은 대부분의 조합에서 0.8 이상의 높은 값을 나타내었다. 숙기의 유전력은 다른 형질에 비해 매우 높아 표현형에 의해 양친을 선택하여 교배하였을 때 후대 실생의 평균치를 추정할 수 있을 것이라고 하였다(Anderman, 1939; Brown, 1960; Haniuda et al., 1975; Hansche et al., 1966, 1972a, 1972b; Hayashi, 1953; Hong, 1991; Kiyoji, 1973; Shin et al., 1983; Wellington and Home, 1944).
Table 4. Heritability of fruit ripening date in the 15 pear families. |
Cross combination | No. of seedlings | Mid-parent | Progeny meanz | Heritability |
Niitaka × Choju | 375 | Sept.12 | Oct. 3.0 ± 13.7 | 0.75 |
Niitaka × Tama | 447 | Sept.15 | Sept.10.0 ± 12.7 | 0.83 |
Niitaka × Chuwhangbae | 378 | Oct.15 | Sept.12.5 ± 12.9 | 0.80 |
Niitaka × Soowhangbae | 202 | Oct. 3 | Oct. 1.2 ± 15.5 | 0.84 |
Niitaka × Satangbae | 116 | Sept.13 | Sept.15.9 ± 18.3 | 0.87 |
Niitaka × Shinsui | 494 | Sept.14 | Sept. 9.6 ± 13.1 | 0.78 |
Shinsui × Satangbae | 96 | Sept. 9 | Sept.12.2 ± 12.0 | 0.68 |
Kosui × Soowhangbae | 134 | Sept.15 | Sept.10.4 ± 12.8 | 0.83 |
Kimizukawase × Soowhangbae | 250 | Sept. 8 | Sept. 5.0 ± 13.3 | 0.76 |
Imamuraaki × Soowhangbae | 168 | Oct.15 | Oct. 9.5 ± 16.6 | 0.86 |
Okusankichi × Soowhangbae | 98 | Oct.17 | Oct.13.2 ± 13.3 | 0.85 |
Chuwhangbae × Soowhangbae | 59 | Oct.10 | Oct.01.1 ± 18.0 | 0.87 |
Chojuro × Soowhangbae | 614 | Sept.24 | Sept.25.8 ± 14.9 | 0.85 |
Shinsui × Whasan | 437 | Sept.13 | Sept.12.6 ± 10.8 | 0.63 |
Kosui × Whasan | 116 | Sept.20 | Sept.12.2 ± 9.5 | 0.71 |
Mean | 265 | Sept.23.7 | Sept.18.5 ± 17.3 | 0.88 |
zMean ± standard deviation. |
교배실생의 숙기 분포
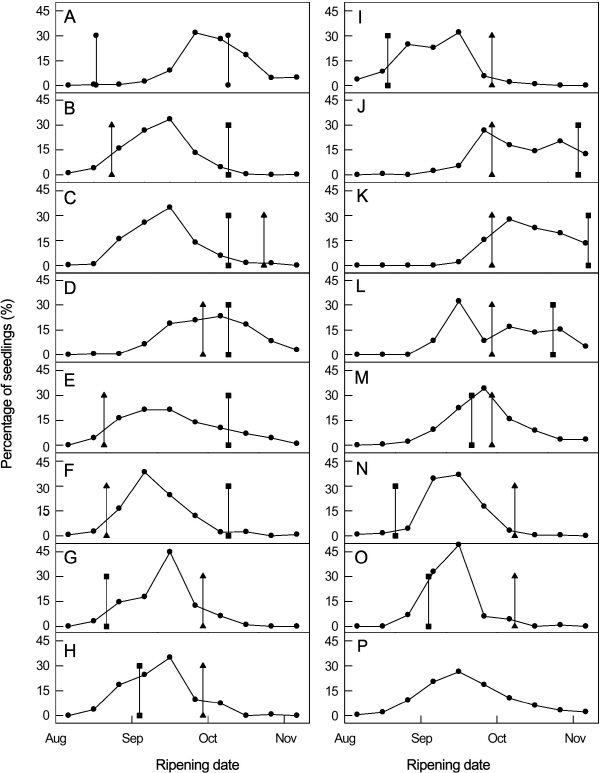
배 교배실생의 숙기는 8월 상순부터 11월 상순까지 매우 폭넓게 분포하였다. 숙기에 대한 교배실생의 교배조합 내 분포는 연속적인 변이를 보이며 대부분의 조합에서 단일 peak를 보여 그 분포는 정규분포를 따르는 것으로 판단되었다(Fig. 1).
교배 양친의 숙기에 따른 교배 실생 분포는 15조합 가운데 12조합이 실생의 최대 빈도가 양친 숙기 범위 안에 분포하였다. 그 나머지 조합에서도 양친 가운데 한쪽 친에 가까운 쪽에 최대 빈도가 분포하는 양상을 보였다. 그러나 ‘추황배’를 교배친으로 한 두 조합에서는 양친의 숙기 범위를 크게 벗어난 실생이 많았다. ‘신고’ × ‘추황배’에서 숙기가 양친보다 빠른 개체의 출현빈도가 높은 것은 과실비대가 빠른 ‘신고’의 특성과 조기 전엽과 충분한 엽수 확보에 따른 당분축적이 빠르게 진행되는 고당도 ‘추황배’의 특성에 따라 숙기 판단에 기준이 되는 과육 식감과 감미가 조기에 형성된 것으로 판단된다. 또한 ‘추황배’ × ‘수황배’ 조합의 실생 평균숙기가 빠른 것은 조사된 후대 개체수가 충분하지 않아 양친 평균숙기보다 다소 빠르게 조사된 것으로 보여진다.
교배실생의 숙기 변이의 방향은 ‘금촌추’, ‘만삼길’, ‘추황배’와 같은 만생종 품종을 교배친으로 한 조합에서는 교배실생의 평균 숙기가 양친 평균보다 빨라 만생에 비해 조생이 우성으로 작용하는 것으로 생각되었다. 그러나 중생종 × 조생종 조합이나 중생종 × 중생종 조합에서는 평균 숙기와 거의 일치되었다. 숙기의 실생 분포에 대하여 많은 연구자들이 양친의 중간치와 실생 평균치가 거의 일치한다고 하였다(Haniuda et al., 1975; Ikeda et al., 1975; Schneider, 1949; Shin et al., 1983). 그러나 조합에 따라 다른 숙기 분포를 보이는 조합도 있다고 하였으며(Brown, 1960; Shin et al., 1983), 본 시험에서도 ‘추황배’를 교배친으로 한 두 조합에서 이와 같은 경향을 나타냈다.
|
Fig. 1. Frequency distributions of fruit ripening date in 15 families of pear seedlings. A, ‘Niitaka’ × ‘Choju’; B, ‘Niitaka’ × ‘Tama’; C, ‘Niitaka’ × ‘Chuwhangbae’; D, ‘Niitaka’ × ‘Soowhangbae’; E, ‘Niitaka’ × ‘Satangbae’; F, ‘Niitaka’ × ‘Shinsui’; G, ‘Shinsui’ × ‘Soowhangbae’; H, ‘Kosui’ × ‘Soowhangbae’; I, ‘Kimizukawase’ × ‘Soowhangbae’; J, ‘Imamuraaki’ × ‘Soowhangbae’; K, ‘Okusankichi’ × ‘Soowhangbae’; L, ‘Chuwhangbae’ × ‘Soowhangbae’; M, ‘Chojuro’ × ‘Soowhangbae’; N, ‘Shinsui’ × ‘Whasan’; O, ‘Kosui’ × ‘Whasan’; P, Total. -●- percentage of seedling, -■- Seed parent, -▲- pollen parent. |
숙기의 양친 평균과 교배실생 평균과의 상관
배 과실 숙기의 양친평균과 실생 평균간에는 높은 정의 상관(r = 0.68**)이 인정되었다(Fig. 2). 즉, 양친 평균의 숙기가 늦을수록 교배실생의 평균 숙기도 늦어졌다. 이러한 결과는 Hong(1991)과 Shin 등(1983)이 각각 산출한 배 숙기의 양친 평균과 실생 평균간의 상관계수 0.75와 0.58과 큰 차이가 없었다. 따라서, 배 숙기에 대한 육종은 교배조합 작성시 양친의 평균이 목표로 하는 숙기와 거의 일치되도록 조합을 작성함으로써 목표로 하는 숙기에 가까운 실생을 가장 많이 획득할 수 있을 것으로 생각되었다.
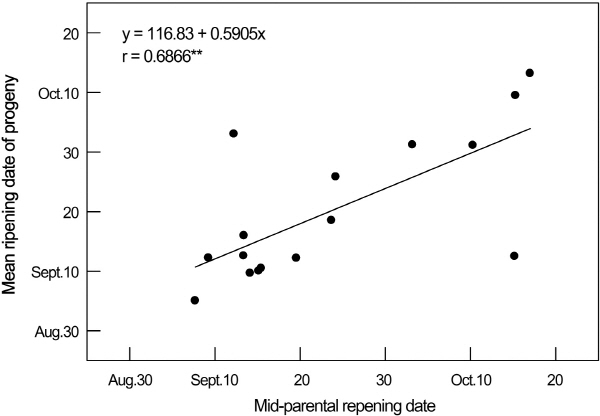
|
Fig. 2. Relationship between mid-parental and progeny mean fruit ripening dates in 15 pear families. |
숙기와 과중간의 상관
배 교배조합별 실생의 숙기와 과중의 상관을 Table 5에 나타냈다. 각 실생 개체간에 숙기와 과실 무게 사이에 정의 상관관계가 인정된 조합이 15조합 가운데 9조합이었으며 실생 전체에 대해서도 높은 정의 상관이 인정되었다.
배 교배실생 집단에 있어서 숙기가 늦은 실생의 과실은 과중이 크고 숙기가 빠른 실생의 과실은 과중이 작은 경향이었다. 본 연구에서도 교배친 품종들의 평균 숙기와 평균 과중간에 높은 상관이 있었고, 이는 숙기와 과중의 상관에 대해서 밝힌 보고(Abe et al., 1993b; Hong 1991; Kim 1985)와 일치하였다. Abe et al.(1993b)은 숙기와 과중간의 상관은 주로 유전적인 상관으로서 유전자의 다면발현 작용임을 강하게 암시했다. 동일 품종 내에 각각 과실 숙기에 있어서 조만의 차이가 나고 품종에 따라서 20일 전후의 차이가 나타나기도 하지만 이 경우에도 과중의 차이는 거의 없는 것으로 보아 과실의 숙기와 과중의 상관은 발육과정에서 환경변화가 미치는 영향은 적고 오히려 유전적 요인이 크게 작용한 것이라고 추정할 수 있다.
또한 과중은 연차간 또는 나무간과 나무 내 과실간 변이가 모두 크므로 2-3년간의 짧은 조사로는 정확히 평가하기가 곤란한 형질이다. 따라서, 배 과실의 숙기와 과중 사이에 밀접한 상관이 존재함으로 숙기에 대해 선발을 행하는 경우 과중에 대해서도 간접적인 선발 효과를 유기할 수 있다는 것을 의미한다. 그러므로 과실 숙기와 과중 사이의 상관을 이용하면 숙기의 선발에 의해 과중의 간접 선발을 할 수가 있을 것으로 생각되었으며, 특정 형질의 선발에 대한 다른 형질의 상관 반응에 대해 앞으로 자세한 검토가 필요할 것으로 생각되었다.
Table 5. Correlations between fruit ripening date and fruit weight of seedlings in 15 pear families. |
Cross combination | No. of seedling | Mean ripening date | Mean fruit wt. (g) | Correlation coefficients (r) |
Niitaka × Choju | 375 | Oct. 3.0 | 305.8 | -0.0760 |
Niitaka × Tama | 441 | Sept.10.0 | 270.7 | 0.3575** |
Niitaka × Chuwhangbae | 378 | Sept.12.5 | 291.2 | 0.2672** |
Niitaka × Soowhangbae | 202 | Oct. 1.2 | 276.0 | -0.0186 |
Niitaka × Satangbae | 116 | Sept.15.9 | 395.7 | 0.3056** |
Niitaka × Shinsui | 494 | Sept. 9.6 | 298.2 | 0.3003** |
Shinsui × Soowhangbae | 96 | Sept.12.2 | 252.5 | 0.2844** |
Kosui × Soowhangbae | 134 | Sept.10.4 | 261.4 | 0.2272** |
Kimizukawase×Soowhangbae | 250 | Sept. 5.0 | 281.1 | 0.2518** |
Imamuraaki × Soowhangbae | 168 | Oct. 9.5 | 304.4 | 0.1135 |
Okusankichi × Soowhangbae | 98 | Oct. 13.2 | 373.0 | 0.1889 |
Chuwhangbae × Soowhangbae | 59 | Oct. 1.1 | 247.1 | 0.1247 |
Chojuro × Soowhangbae | 614 | Sept.25.8 | 281.9 | 0.1890** |
Shinsui × Whasan | 437 | Sept.12.6 | 272.9 | 0.1521** |
Kosui × Whasan | 116 | Sept.12.2 | 256.9 | -0.0188 |
Mean | 265 | Sept.18.5 | 289.5 | 0.1968** |
**Significant at p = 0.01. |
숙기 관련형질의 우량 개체 출현 빈도의 산출
배 교배조합별 후대 실생의 조숙(8월 25일 이전) 개체 및 만숙(10월 15일 이후) 개체의 비율은 Table 6과 같다. 양친의 숙기가 빠를수록 후대에 숙기가 빠른 실생 개체 비율이 높았고 양친 숙기가 늦을수록 후대 실생의 숙기가 늦은 개체 비율이 높았다. 양친의 평균 숙기와 조숙 및 만숙 실생의 비율과는 높은 상관(Fig. 3)이 인정되었고, 이들로부터 각각의 회귀식 y = -0.4605 × +146.88(r = -0.5852*)과 y = 0.9287 × -257.52(r = 0.7157**)를 얻었다. 이 두 회귀식을 이용하여 배 숙기에 대한 양친 평균과 그 실생 후대에 조숙 개체 및 만숙 개체 출현빈도에 대한 이론적인 수치를 구한 값은 Table 7과 같다. 양친 평균 숙기가 8월 하순 이전일 때 조생 실생의 출현빈도는 24.9% 이상이었으며, 양친 숙기가 10월 중순 이후일 때 만생 실생의 출현빈도는 35.0% 이상이었다.
그러므로 육종 시 목표로 하는 개체를 많이 확보하기 위해서는 양친의 숙기 평균치를 목표 숙기에 가깝게 하는 것이 필수적이다.
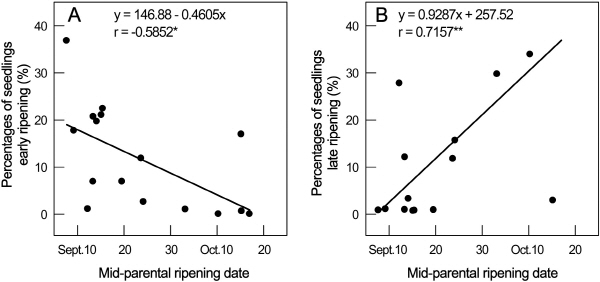
|
Fig. 3. Relationship between mid-parental fruit ripening date and percentage of early (A)- and late (B)- ripening seedling in 15 pear families. |
Table 6. Percentage of early (Aug. 25 or earlier) and late (Oct. 15 or later) ripening seedlings in 15 pear families. |
Cross combination | No. of seedling | Mid- parent | Percentage of seedlings |
Aug.25. or earlier | Oct.15 or later |
Kimizukawase× Soowhangbae | 375 | Sept. 7.7 | 36.8 | 0.8 |
Shinsui × Soowhangbae | 447 | Sept. 9.3 | 17.7 | 1.0 |
Niitaka × Chojo | 378 | Sept.12.2 | 1.1 | 27.7 |
Shinsui × Whasan | 202 | Sept.13.4 | 6.9 | 0.9 |
Niitaka × Satangbae | 116 | Sept.13.4 | 20.7 | 12.1 |
Niitaka × Shinsui | 494 | Sept.14.2 | 19.6 | 3.2 |
Niitaka × Tama | 96 | Sept.15.2 | 21.0 | 0.7 |
Kosui × Soowhangbae | 134 | Sept.15.5 | 22.4 | 0.7 |
Kosui × Whasan | 250 | Sept.19.6 | 6.9 | 0.9 |
Chojuro × Soowhangbae | 168 | Sept.24.2 | 2.6 | 15.6 |
Niitaka × Soowhangbae | 98 | Oct. 3.2 | 1.0 | 29.7 |
Chuwhangbae × Soowhangbae | 59 | Oct.10.3 | 0.0 | 33.9 |
Niitaka × Chuwhangbae | 614 | Oct.15.2 | 16.9 | 2.9 |
Imamuraaki × Soowhangbae | 437 | Oct.15.3 | 0.6 | 47.0 |
Okusankichi × Soowhangbae | 116 | Oct.17.0 | 0.0 | 55.1 |
Mean | 265 | Sept.23.7 | 11.8 | 11.7 |
Table 7. Estimated frequency of early (Aug.25 or earlier) and late (Oct.15 or later) ripening seedlings by mid-parent ripening date. |
Mid-parental date | Aug. | | Sept. | | Oct. | | Nov. |
05 | 15 | 25 | 05 | 15 | 25 | 05 | 15 | 25 | 05 |
Percentage of early seedling (%) | 34.1 | 29.5 | 24.9 | | 20.2 | 15.6 | 11.0 | | 6.4 | 1.8 | 0 | | 0 |
Percentage of late seedling (%) | 0 | 0 | 0 | | 0 | 7.2 | 16.5 | | 25.7 | 35.0 | 44.3 | | 53.6 |
초 록
동양배의 교배육종 효율을 증진시키기 위하여 13품종의 교배친을 이용한 15개 교배조합으로부터 얻어진 실생 4,035주를 시험재료로 과실숙기에 대한 유전력, 양친 평균과 실생 평균과의 상관과 후대 실생의 분포 특성을 분석하였다. 10년간 조사된 교배친 13품종의 숙기 평균과 표준편차 및 변이계수를 분석한 결과 대부분의 조생종의 숙기가 만생종에 비해 숙기 변이가 큰 것으로 나타났다. 그러나 모든 품종의 숙기 변이 계수는 0.92-3.41사이로 과실의 다른 형질에 비해 변이 폭이 좁고 안정된 형질임을 보여주었다. 유전력은 대부분의 조합에서 0.8 이상의 높은 값을 나타내었다. 숙기의 유전력은 다른 형질에 비해 매우 높아 표현형에 의해 양친을 선택하여 교배하였을 때 후대 실생의 평균치를 추정할 수 있었다. 교배 실생의 숙기 변이의 방향은 만생종 품종을 교배친으로 한 조합에서는 교배 실생의 평균숙기가 양친 평균보다 빨라 만생에 비해 조생이 우성으로 작용하는 것으로 생각되었다. 그러나 중생종 × 조생종 조합이나 중생종 × 중생종 조합에서는 평균 숙기와 거의 일치되었다. 배 과실 숙기의 양친 평균과 실생 평균간에는 높은 정의 상관(r = 0.68**)이 인정되었다. 배 과실의 숙기와 과중 사이에 밀접한 상관이 존재함으로 숙기에 대해 선발을 행하는 경우 과중에 대해서도 간접적으로 선발이 가능한 것으로 판단되었다.
Acknowledgements
본 연구는 농촌진흥청 공동연구사업(과제번호: PJ010507)의 지원에 의해 이루어진 것이며, 2013년도 강원대학교 학술연구조성비를 겸하여 연구하였음(과제번호-C1009748-01-01).
References
Abe, K., Y. Sato, T. Saito, A. Kurihara, and K. Kotobuki.1993a. Inheritance of ripening time of fruit of Japanese pear (Pyrus pyrifolia Nakai). Jpn. J. Breed. 43:289-298.
Abe, K., Y. Sato, T. Saito, A. Kurihara, and K. Kotobuki.1993b. Genetic correlation between ripening time and weight of fruits in Japanes pear (Pyrus pyrifolia Nakai). Jpn. J. Breed. 43: 439-447.
Anderman, D. C. 1939. Apple breeding; Inheritance and statistical studies on the fruits of crossbred seedling with Antonoyka parentage. Proc. Am. Soc. Hortic. Sci. 36:279-283.
Brown, A. G. 1960. The inheritance of shape, size and season of ripening in progenies of the cultivated apple. Euphytica 9:327-337.
Haniuda, T., Y. Yoshida, and S. Tsuchiya. 1975. Studies on the inheritance of fruit characters of apple. I. Inheritance of ripening season. Bull. Fruit Tree Res. Stn. C. 2:1-11.
Hansche, P.E., C.O. Hesse, and V. Beres. 1972a. Estimates of genetic and environmental effects on several traits in peach. J. Am. Soc. Hortic. Sci. 97:76-79.
Hansche, P.E., V. Beres, and H.I. Forde. 1972b. Estimates of quantitative genetic properties of walnut and their implications for cultivar improvement. J. Am. Soc. Hortic. Sci. 97:279-285.
Hansche, P.E., V. Beres, and R.M. Brooks. 1966. Heritability and genetic correlation in the sweet cherry. Proc. Am. Soc. Hortic. Sci. 88:173-183.
Hayashi, H. 1953. Inheritance of Asian pear. II. Studies on the inheritance of major characters on temperate fruits (peach, Asian pear and persimmon). Agric. Technol. Inst. Bul. E. 2:34-53.
Hong, K.H. 1991. Inheritance studies of major morphological and horticultural characters in pear (Pyrus pyrifolia var. culta Nakai). PhD Diss., Chonnam Natl. Univ., Gwangju, Korea.
Ikeda, I., T. Oohata, T. Nishida, and K. Hirose. 1975. The inheritance of some characters in Japanese persimmon (Diospyros kaki Thunb.). III. The inheritance of season of repening. Bull. Fruit Tree Res. Stn. B. 2:1-20.
Kim, W.C. 1985. Studies on the inheritance of major characters in Oriental pear cultivars (Pyrus pyrifolia var. culta Nakai). PhD Diss., Seoul Natl. Univ., Seoul, Korea.
Kiyoji, K. 1973. Research on apple breeding. Bull. Akita Fruit Tree Res. Stn. 6:1-82.
Korea Rural Economic Institute (KREI). 2014. Agriculture prospect 2014. p. 253.
Ministry for Food, Agriculture, Forestry, and Fisheries (MFAFF). 2011. Census of Agriculture Forestry and Fisheries. p. 302.
Rural Development Administration (RDA). 1995. Manual for agricultural investigation. Suwon, Korea.
Park, S.Z. 1987. Genetic breeding for crop. Kor. Natl. Open Univ. p. 257-303.
Schneider, G.W. 1949. Characteristics of progeny from certain apple crosses. Proc. Am. Soc. Hortic. Sci. 53:205-212.
Shin, Y.U., Y.J. Yim, H.M. Cho, B.W. Yae, M.S. Kim, and Y.K. Kim. 1983. Studies on the inheritance of fruit characters of oriental pear, Pyrus serotina Rehder var. culta. Res. Rept. ORD 25(Hort.):108-117.
Statistics Korea. 2013. Korean Statistical Information Service. http://kosis.kr/statisticsList/statisticsList_01List.jsp?vwcd= MT_ZTITLE&parentId=F#Subcont.
Wellington, R. and G.H. Home. 1944. The performance of seedlings derived from selfing and crossing the McIntosh apple. Proc. Am. Soc. Hortic. Sci. 44:273-279.
Yamaguchi, M., M. Yoshida, and H. Kyotani. 1984. Studies on the distribution of ripening time of certain progenies in peach breeding. Bull. Fruit Tree Res. Stn. A. 11:15-33.


