서 언
재료 및 방법
식물재료
과실과 종자의 수확 및 특성 조사
기내파종
단마디배양
다신초 유도
유식물체의 기외 순화
통계분석
결과 및 고찰
과실과 종자의 외형
저온처리가 종자 발아에 미치는 영향
실생의 표현형 분리와 초기 생육
실생에서 마디의 위치에 따른 증식 효율 검정
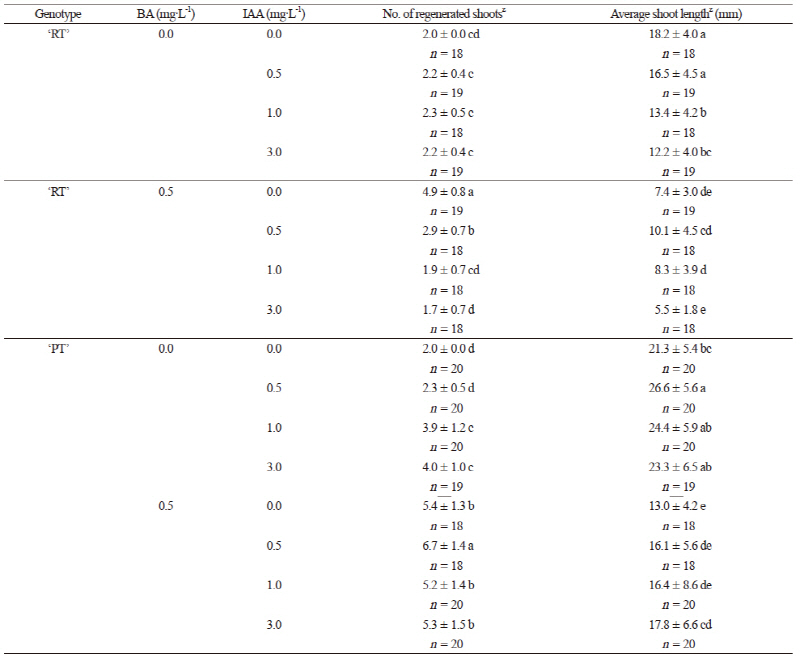
마디로부터 다신초 발생에 미치는 BA와 IAA의 영향
증식 유식물체의 기외 순화와 생활환 완성
서 언
해가 거듭될수록 다양한 생물 종이 빠르게 절멸되고 있는데 그 중 식물 종이 가장 큰 부분을 차지한다(IUCN, 2015). 특정 식물 종의 절멸은 그 종이 가지고 있던 생태적 지위가 상실된다는 것을 의미하며, 이는 생태계 교란을 야기할 수도 있어 인간생활에 직?간접적 피해를 줄 뿐만 아니라 유전자원으로서의 미래 가치는 소실될 우려가 있다. 따라서 절멸의 위기에 처한 희귀 식물 종의 자생지와 개체군을 보전하는 것은 현재를 살아가는 우리 세대의 의무라고 할 수 있다. 갯봄맞이(Glaux maritima L.)는 한국의 멸종위기 야생동·식물 적색자료집에서 최근에서야 취약(VU: vulnerable)종으로 구분되었으며, 멸종위기 야생생물 II급에 속한다(NIBR, 2012). 북한을 비롯한 세계 각국의 염습지에 분포하지만 남한에는 속초, 울산 및 포항의 자생지만 알려져 있는데, 속초 자생지는 거의 사라졌으며 그나마 포항과 울산 자생지의 개체군도 극히 소규모인 것으로 보고되었다(Son et al., 2011; NIBR, 2012; IUCN, 2015).
갯봄맞이는 진달래목(Ericales) 앵초과(Primulaceae)에 속하는 것으로 알려져 있지만, Kallersjo et al.(2000)은 자금우과(Myrsinaceae)에, APG(The Angiosperm Phylogeny Group, 2009)는 APG III 분류체계에서 자금우아과(Myrsinoideae)에 포함시키고 있는 유사일년생 초본(pseudo-annual herbaceous plant)이다. 모식물체는 하계 생육기에 지하부에 휴면아(hibernacle)가 착생한 offshoot를 신장시킨다. 겨울로 접어들면서 모식물체의 지상부와 뿌리 모두는 차츰 말라 죽고 offshoot에 착생한 휴면아는 월동을 한 후 이듬해 봄에 맹아 하여 봄과 여름 동안에 지상부 생육, 개화, 그리고 결실을 하는 생활환을 가진다(Jerling, 1988b). 이러한 생활환에 관한 관찰결과는 본 종의 종자번식 또는 영양번식을 시도할 때 참고할 수 있는 귀중한 정보라 할 수 있다.
갯봄맞이는 초장이 5-20cm 정도이고 화관 직경 1cm 내외의 아담한 꽃이 이른 봄부터 여름철에 이르기까지 연속하여 개화하므로 장기간에 걸쳐 즐길 수 있어 화단용 또는 분화용으로 이용가치가 높다. 또한 자생지에서 종자번식과 더불어 언급한 바와 같이 자발적 영양번식을 하는 것으로 관찰되므로 한번 재식하면 다년간에 걸쳐 별도의 번식관리를 할 필요가 없는 장점이 있는 것으로 판단된다. 이러한 갯봄맞이의 생육습성은 원예작물로서의 개발가능성을 시사하는 측면이라고 사료된다. 한편, 온대지역인 우리나라에서의 갯봄맞이 자생지는 해안가 염습지에 분포하고 있어 충분한 습도를 유지하고 있는 곳인데, Khan and Gul(2006)은 습도가 높은 온대지역 염생식물(halophyte)의 경우 저온처리가 종자발아를 향상시키는 경향이 있다고 보고한 바가 있는데 이는 갯봄맞이 종자의 자생지외 발아 유도에 참고할 만 하다.
희귀 식물의 인공 번식과 보전을 위한 효과적이 방법의 하나로 기내배양기술을 들 수 있다. Eucalyptus phylacis, Dianthus superbus ssp. superbus, Hemiandra rutilans 및 Symonanthus bancroftii 등의 다양한 희귀 식물 종의 번식, 보전 및 복원에 실제 기내배양 기술이 활용되고 있다(Mikulik, 1999; Bunn et al., 2011; Pence, 2011; Pence, 2013). 그러나 본 연구의 재료인 갯봄맞이에 관한 연구문헌은 주로 생태적 정보를 다루고 있어(Rozema and Riphagen, 1977; Rozema et al., 1977; Jerling, 1988a; Jerling, 1988b; Jerling and Berglund, 1994; Druva-Lusite et al., 2008) 기내번식 체계 확립을 목적으로 하는 부분은 부족하다.
따라서 본 연구는 해안식물의 한 종인 갯봄맞이를 재료로 이용하는 다양한 심화연구를 수행하기에 앞서 개체군 밀도가 매우 낮아 유전적 다양성 또한 극히 협소한 것으로 추정되는 현 자생지의 교란을 최소화할 수 있도록 종자 유래 영양계를 확보하기 위한 목적으로 수행되었다. 이를 위해 갯봄맞이 종자의 발아에 미치는 저온처리의 영향을 파악하기 위한 실험이 병행되었다. 또한 향후 이 종의 서식지 복원에 필요한 기내증식 체계의 확립과 내륙에서 재배할 수 있는 원예종으로의 개발 가능성을 타진하기 위한 목적으로 기내 마디배양에 영향을 미치는 제조건을 탐색하였다.
재료 및 방법
식물재료
경상북도 포항시 해안지역(N36°06'24"/E129°57'49")에서 수집하여 사양토를 담은 화분에 심어 자연환경 조건에서 별도의 시비 없이 담수관수하며 재배되고 있던 갯봄맞이(Glaux maritima L.) 식물체를 2009년 분양 받아 2012년까지 본 연구의 실험재료로 사용하였다. 분양 받은 화분에는 줄기 부위와 꽃이 연한 붉은색을 띄는 개체와 줄기는 연한 녹색이고 꽃은 흰색을 띄는 개체가 혼식되어 있었는데, 본 연구에서는 전자를 ‘Red type’(RT)으로 후자를 ‘Pistachio type’(PT)으로 명명하고 각 실험에 구분하여 사용하였다(Fig. 1).

과실과 종자의 수확 및 특성 조사
갯봄맞이의 자가수정 또는 타가수정 가능 여부와 화분매개 방식 등 생식생리에 관해서는 알려진 바가 없다. 본 연구에서는 방임 수분을 통해 모식물체에 착생한 과실(삭과)의 과피 표면이 완전히 갈변된 시점인 8월 하순에 각 모식물체의 표현형 구분 없이 삭과를 수확하였다. 과피를 완전히 건조시킴으로써 탈종을 손쉽게 할 수 있도록 배양실(25 ± 2°C, 상대습도 50 ± 5%, 50μmolㆍm-2ㆍs-1, 16h 명/8h 암)에서 약 10주간 건조시켰다. 건조처리가 완료된 시점에 대부분의 열개되지 않은 과실을 대상으로 그 길이와 폭을 조사한 후 탈종하여 과실당 종자 수를 조사하였다. 또한 정선된 종자는 그 길이와 폭 및 천립중을 조사하였다. 종자의 특성 조사 후 실리카겔을 넣은 페트리접시에 담아 밀봉한 후 4°C 냉장고에서 암상태로 건식 보관하면서 다음 실험의 재료로 사용하였다.
기내파종
4°C 냉장고에서 약 4개월간 암상태로 건식 보관한 종자를 70% 에탄올에 1분간 침지한 후 멸균수로 3회 세척하였다. 계면활성제인 sodium dodecyl sulfate(Sigma-Aldrich, USA)가 0.2% 첨가된 25% CloroxTM(유효 염소 4% 이상) 용액에서 25분간 교 반한 후 멸균수로 5회 세척하여 종자의 표면을 살균하였다. 표면살균된 종자는 건조된 멸균 여과지에서 습기를 제거시킨 후 sucrose(Duchefa Biochemie, Netherlands) 30gㆍL-1, Plant agar(Duchefa Biochemie) 8gㆍL-1가 첨가되고 pH가 5.8로 조정된 MS 배지(MS: Murashige and Skoog, 1962) (Duchefa Biochemie)가 분주된 배양병에 5립씩 치상(1개 반복)하였다. 파종된 배양병을 4°C 냉장고(암 상태)에 넣은 후 2주 간격으로 일부를 꺼내 25 ± 2°C, 명배양(50μmolㆍm-2ㆍs-1, 16h 명/ 8h 암)하면서 발아를 유도하였다. 명배양 2주 후 종자의 발아율과 오염율을 조사하는 한편 발아된 실생의 줄기 색깔을 기준으로 표현형을 판정하였다.
단마디배양
기내 파종하여 8주간 저온처리한 후 명배양 조건으로 이동하여 7주간 생육시킨 유식물체의 표현형은 재료 식물체와 동일하게 줄기 색깔에 따라 ‘RT’와 ‘PT’로만 구분되었다. 각각의 표현형에서 마디가 5개 이상 포함 된 유식물체를 단마디 배양의 재료로 이용하였는데, 위로부터 정단, 제2절, 제3절, 제4절 및 제5절로 나누어 절편을 조제한 후 각각의 정단부와 마디의 액아로부터 신초를 유도하였다. 배지는 기내배양의 선행연구 결과에 관한 문헌을 찾을 수 없는 상황에서 하나의 대조배지로서 염스트레스 내성 검정에 Freipica and Ievinsh(2010)가 사용한 sucrose 30gㆍL-1, Plant agar 8gㆍL-1, 50mM NaCl(Sigma-Aldrich), Eriksson vitamin(Duchefa Biochemie)이 첨가되고 pH가 5.8로 조정된 1/2MS basal salt(Duchefa Biochemie) 배지(modified 1/2MS: Mod. 1/2MS)와 sucrose 30gㆍL-1, Plant agar 8gㆍL-1가 첨가되고 pH가 5.8로 조정된 MS배지(Full MS) 등 두 종류를 사용하였다. 각 절편은 25 ± 2°C, 명배양 조건에서 6주간 배양한 후 생장한 신초의 각 마디수, 길이, 발근 유무 및 근단부 갈변 유무 등을 조사하였다.
다신초 유도
마디배양의 결과를 바탕으로 각 표현형별 최적 위치의 마디를 사용하여 다신초 유도 실험을 수행하였다. 6-benzylaminopurine(BA; Duchefa biochemie) 0.5mgㆍL-1, indole-3-acetic acid(IAA; Fluka chemie AG, Switzerland) 0.5-3.0mgㆍL-1를 단용 또는 혼용하여 sucrose 30gㆍL-1, Plant agar 8gㆍL-1가 첨가되고 pH가 5.8로 조정 된 MS 배지에 치상하여 6주간 25 ± 2°C에서 명배양 하였다. 배양 6주 후에 유도된 신초의 수, 생장한 각 신초의 길이, 발근 유무, 근단부의 갈변 유무 등을 조사하였다.
유식물체의 기외 순화
다신초 유도로부터 얻어진 유식물체 clump 중 지상부와 근권부 생장이 양호한 것(Fig. 4의 ‘RT’ 표현형에서 IAA 단용 첨가구 제외)을 기외 순화의 재료로 사용하였다. 유식물체 clump의 근권부에 묻은 배지를 흐르는 수돗물로 완전히 세척한 후 상업용 상토(참사랑, Nongwoo Bio, Korea)가 담긴 10cm × 8.5cm 플라스틱 화분에 이식하였다. 유묘 clump가 옮겨 심긴 화분을 투명한 플라스틱 봉투에 넣고 배양실에서 기외 순화를 진행하였다. 배양실 환경은 25±2°C, 명배양 조건이었으며, 점진적으로 플라스틱 봉투에 구멍을 뚫어 유식물체를 저습한 외기에 적응시켰다. 순화 시작 2주 후 플라스틱 봉투를 완전히 제거한 후 화분을 무가온 플라스틱 필름 온실(좌표N36°22'46"/E128°08'51")로 옮겨 자연환경에서 생육시켰다. 관수는 육안으로 관찰하여 표토가 마르는 시점에 담수(수돗물)로 충분히 실시하였다.
통계분석
각 실험에서 조사 또는 측정된 값의 통계분석에는 SAS version 9.1.3 service pack 4(SAS institute Inc., Cary, NC, USA) 프로그램을 이용하였는데 처리간 평균의 비교는 Duncan’s multiple range test(DMRT)를 적용하여 p < 0.05 수준에서 실시하였다. 각 실험에서 처리당 반복수(n)는 다양하여 제시된 각 표에 직접 표시하였다.
결과 및 고찰
과실과 종자의 외형
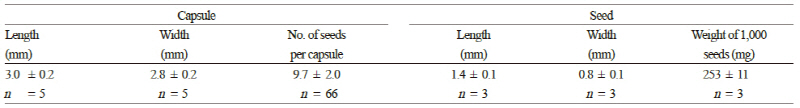
모식물체에 착생한 과실은 녹색에서 갈색으로 익어가는 만두모양의 삭과(capsule)였으며 열개하지 않았을 때의 삭과 크기는 길이와 폭이 각각 3.0mm와 2.8mm였고 삭과당 종자수는 9.7개였다(Fig. 2 and Table 1). 이러한 외형은 Strandkryp and Rannikki(2009)가 묘사한 것과 일치하는 것이다. 삭과는 후숙과 건조가 진행되면서 5개의 심피실 격벽을 따라 포간개열(septicidal dehiscence; Fig. 2C)하였는데 이러한 과실의 열개방식은 자연상태에서 종자의 확산을 돕는 현상 중 하나로 알려져 있다(Day, 2000). 탈종된 종자의 색은 짙은 갈색으로 길이는 1.4mm, 폭은 0.8mm였으며 천립중은 253mg으로 전형적인 미세종자의 특징을 나타내었다(Fig. 2D and Table 1). 주사전자현미경을 이용하여 앵초과와 자금우과 등의 종자를 대상으로 그 모양을 상세히 조사한 Morozowska et al.(2011)의 보고에서도 갯봄맞이 종자는 각이 져있고 표면은 그물 모양이며 길이와 폭이 각각 1.2mm와 0.9mm, 두께는 0.5mm, 그리고 제(hilum)는 좁다고 밝힌 바 있다.
| |
Fig. 2. Appearances of capsule and seed of Glaux maritima L. (A) Fruit (arrow) set on axil, (B) Mature capsule before dehiscence, (C) Capsule bearing seeds just after dehiscence, (D) Collected seeds. | |

저온처리가 종자 발아에 미치는 영향
본 연구의 채종방법(재료 및 방법 참조)에 따라 얻어진 갯봄맞이 종자의 발아에 미치는 저온처리의 효과를 확인하고자 저온처리 기간을 달리하여 발아율을 조사하였다. 탈종 후 4°C에 약 4개월간 건식 보관된 종자를 기내파종하여 곧바로 25°C 명상태 에서 발아를 유도하였을 때는 발아를 전혀 하지 않았지만 기내파종 한 후 4°C에서 2주 이상 처리하였을 때는 발아한 종자가 관찰되었다. 저온처리 기간에 따라 발아율에 차이를 나타내었는데, 파종 후 저온처리기간이 2주인 경우는 20.0%, 4주 이상에서는 처리기간 간에 차이가 없이 91.4-94.5%의 종자가 발아하였다(Table 2). 갯봄맞이를 염생식물로 구분할 수 있는지에 대해 학자에 따라 이견이 있을 수 있지만(Rozema, 1975; Khan and Gul, 2005; Flowers and Colmer, 2008), 언급한 실험 결과는 갯봄맞이 종자도 다른 다양한 염생식물의 경우와 유사하게 발아를 위해서는 필수적으로 수분을 흡수한 이 후 상당한 기간의 저온처리가 필요하다는 것을 나타내는 것이라 할 수 있다(Rozema, 1975; Khan and Gul, 2005). 한편, 파종 후 5.6 ± 1.5%(mean ± S.D., n = 4) 의 종자만이 미생물에 오염된 것으로 나타났으며 최종 발아율을 참조했을 때 CloroxTM 살균이 발아에 거의 영향을 미치지 않은 것으로 나타나(Table 2) 본 연구에서 실시한 종자의 표면살균 방법이 실용적 측면에서 활용 가능한 것으로 판단된다.

실생의 표현형 분리와 초기 생육
발아 3주 후 조사한 실생의 마디색은 개체별로 적색(‘RT’)과 연녹색(‘PT’)으로 중간 발색의 특성을 가진 개체 없이 뚜렷하게 구분되었다(Fig. 3B-3F). 본 연구에서 수회에 걸쳐 기내 파종한 종자로부터 유래한 전체 유묘를 대상으로 조사한 분리비는 약 82:18(‘RT’:‘PT’)로 본 실험에 사용한 종자집단은 ‘RT’가 우점하는 것으로 판단할 수 있다. 그러나 이러한 결과가 자생지 개체군 또는 집단의 구성을 절대적으로 반영하는 것인지, 더불어 색소발현의 유전양식이 어떠한지에 관해서는 좀 더 세밀한 연구가 필요한 것으로 사료된다. 기내 파종하여 충분한 저온을 거친 종자는 상온으로 이동된 후 약 5일경부터 표현형에 무관하게 유근을 출현시켰고, 발아 5주 후에는 마디가 3-4개, 초장이 약 1.5cm인 유묘로 생장하였다(Fig. 3).
실생에서 마디의 위치에 따른 증식 효율 검정
종자 파종 및 저온처리를 완료하고 상온으로 이동하여 발아를 유도한 후 7주가 경과한 실생을 다섯개 부위로 세절하여 두 종류의 배지에 각각 배양함으로써 마디의 위치와 배지종류가 증식효율에 미치는 영향을 검토하였다. 정단부위는 모두 한 개의 신초를 신장시켰고, 마디는 대생(opposite)한 액아로부터 대부분 2개의 신초를 신장시켰다. 정단부위를 제외하면, 표현형과 배지 의 종류에 무관하게 제3-4절까지 하위 마디일수록 새롭게 신장한 두 개 신초의 마디 수의 합(총 마디수)과 평균 신초 길이가 증가하는 경향을 나타내었다(Table 3). 액아 유래 신초의 총 마디 수를 기준으로 판단한 증식효율이 높은 제4절과 제5절을 Mod. 1/2MS배지에 배양하였을 때 ‘RT’와 ‘PT’ 표현형간 신초의 총 마디 수는 큰 차이를 나타내지 않았으나(제4절: LSD = 1.6, p < 0.05; 제5절: not significant), Full MS배지에 배양하였을 때는 뚜렷한 차이(제4절: LSD = 1.7, p < 0.001; 제5절: LSD = 1.3, p < 0.01)를 나타내어 ‘PT’의 신초 생장이 ‘RT’에 비해 우수하였다(Table 3). 한편, ‘PT’ 표현형의 경우 Mod. 1/2MS배지에 비해 Full MS배지에서 신초의 총 마디 수와 평균 길이 모두가 확연하게 우수하였는데 이는 Mod. 1/2MS배지에는 MS염뿐 아니라 50mM의 NaCl이 추가 첨가되었고 엽다육성 염생식물(leaf succulent halophyte)의 경우 70mM의 NaCl에서 발아 등이 억제된다는 보고와 무관하지 않은 것으로 사료된다(Rozema, 1975; Khan and Gul, 2005).


마디로부터 다신초 발생에 미치는 BA와 IAA의 영향
식물생장조절제 무첨가 배지에서의 마디배양 실험결과를 토대로 제4절과 제5절을 선택하고 각 마디로부터 다수의 신초를 유도하기 위해 BA와 IAA가 단용 또는 혼용 첨가한 Full MS배지에 두 표현형의 절편을 배양하였다. ‘RT’의 경우 IAA 단용 또는 BA와의 혼용배지에서 IAA농도가 증가할수록 신초의 분화 수가 줄어들었고 유식물체는 갈변을 동반하며 생장이 위축되는 경향을 보였다(Table 4 and Fig. 4A). 반면, ‘PT’의 경우는 IAA 단용 첨가 배지에서 IAA의 농도가 높아질수록 근소하게 신초의 분화 수가 증가하였는데 이러한 신초 분화 수의 증가는 BA를 혼용했을 때 더 뚜렷해졌다(Table 4 and Fig. 4A). 특징적인 것은 동일 종이며 수집 당시 ‘RT’와 ‘PT’가 생육 경쟁을 하는 한 화분에 혼식되어 있었음에도 불구하고(Fig. 1) 색소발현 양상이 다른 두 표현형이 동일한 기내배양 환경에서는 뚜렷하게 다른 반응을 보인다는 점이다. 이러한 현상은 갯봄맞이 기내배양에 관한 이전 연구 결과를 찾아볼 수 없는 현 실정에서 본 보고 이후에도 다방면의 접근을 통해 세부적으로 탐구해볼 부분이라 생각된다.
증식 유식물체의 기외 순화와 생활환 완성
마디배양으로부터 얻어지고 지상부와 근권부 생육이 모두 양호한 ‘RT’와 ‘PT’ 표현형 갯봄맞이 유식물체 clump(V0 세대)를 2011년 8월에 기외 순화시켰다. 상토가 담긴 화분에 이식하여 배양실과 온실에서 생육시킨 두 표현형의 유식물체는 모두 생존하였으며(Fig. 4B) 기외 이식 당년에는 개화하지 않고 영양생장만 지속하다가 한 겨울이 다가옴에 따라 지상부와 뿌리가 모두 고사하고 여름철 생장한 offshoot에 발생한 휴면아(hibernacle)만 생존하였다. 이듬해 봄에 지하부의 휴면아는 맹아하여 새로운 포기(V1 세대)로 생장하면서 하위 엽액에 꽃봉오리를 착생시켰다. 4월에 개화하였고 방임수분을 통해 정상적으로 삭과를 맺었으며(Fig. 4D), 동년 8월에는 지하부에서 이미 생장한 offshoot와 분화된 휴면아를 관찰할 수 있었다. 이러한 기내 증식 갯봄맞이의 생활환에 대한 관찰결과는 Jerling(1988a)이 자생지에서 관찰한 갯봄맞이의 생활환과 일치하는 것이며, 해안가 염습지가 아닌 내륙에서 담수관수를 통해서도 식물체의 정상 생육과 생활환 완성이 가능하다는 것을 나타내는 것이다. 염생식물(halophyte)의 기존 정의에 대한 모호성을 들어 Flowers and Colmer(2008) 및 Flowers et al.(1986)은 최소 200mM의 NaCl 조건에서도 생활환을 완성할 수 있는 것을 염생식물로 재정의한 바 있고 Khan and Gul(2005)은 Rozema(1975)의 보고를 참고하여 그 동안 염생식물로 알려진 갯봄맞이의 종자는 90mM의 NaCl에서 발아가 저해될 수 있다고 정리하였다. 본 연구의 목적인 갯봄맞이의 기내 증식 체계 개발과는 별개로, 언급한 관찰결과에서 보듯이 갯봄맞이가 혐염식물(glycophyte)과 다르지 않게 내륙지역에서 담수관수를 통해서도 정상적인 생활환 완성이 가능함에도 불구하고 갯봄맞이의 자생지가 해안가 또는 염습지에 한정해서만 분포하는지에 관한 생태적ㆍ생리적 연구가 필요한 것으로 사료된다.