

Introduction
Materials and Methods
Experimental site and materials
Experimental design and treatment
Samples and methods
Statistics and analysis
Results
Seed morphology and germination
Seedling growth characteristics
Chlorophyll fluorescence parameters of different passion fruit rootstocks to drought stress
Antioxidant enzyme activities of different passion fruit rootstock seedlings to drought stress
Evaluation of the comprehensive characteristics of different passion fruit rootstocks
Discussion
Conclusion
Introduction
Passion fruitis a type of perennial evergreen vine belonging to Passiflora L. of Passifloraceae. This genus contains more than 500 species, which can be divided into edible, medicinal, and ornamental according to the usage (He et al., 2020; Xia et al., 2021). Therein, a total of nine localized new varieties, such as ‘Jindu No. 3’, and ‘ZXMB’, of edible passion fruit have been cultivated since being introduced in China in 1901. Specifically, the fruits are rich in K, Ca and other mineral elements as well as bioactive substances, including amino acids, organic acids, Vc and flavonoids and are thus classified as medicinal and edible fruits (Ramaiya et al., 2021). Among them, Passiflora edulis f. flavicarpa enjoys high market acceptance due to its strong resistance and high sugar degree, and its economic benefits exceed those of staple grain crops (e.g., rice, corn, soybean) and economic crops (e.g., citrus, Chinese medical herbs, edible fungi).
At present, passion fruit is grown in Guangxi, Fujian, Yunnan, Hainan, and Guizhou provinces in China, and the planting area in Guizhou has been stabilized at over 13,300 ha for many years. Due to continuous large-scale planting over many years, rot-inducing pathogens in passion fruit stems, such as Fusarium oxysporum f. sp.Passiflorae and F. solani, have accumulated in the soil (Salazar et al., 2016; Wang et al., 2023b). This issue is often managed by pesticide disinfection, crop rotation or interplanting, soil improvements, and the breeding of resistant varieties. However, the extensive use of pesticides causes soil deterioration and water pollution, threatening the health of animals and humans (Wang et al., 2021). Rotation or interplanting provides insects with more places for activity and with more food, thus increasing the viral transmission risk among passion fruit plants, such as rotation or interplanting of passion fruit with Cucurbitaceae plants (e.g., watermelon and cucumber) and Solanaceae plants (including tomato and potato). Soil improvements require considerable manpower and cost inputs, with only limited economic benefits. The exploration of resistant resources and the breeding of resistant varieties are the most economical and feasible strategies, and the grafting of resistant rootstocks and excellent cuttings is one of the most common technologies (Chen et al., 2021). Therefore, obtaining high-quality rootstock is an important task for high-quality seeding production.
Drought, a major type of abiotic stress, is featured by higher frequencies, longer durations and greater hazard levels compared to relevant data over the past decades, as its risk is aggravated by greenhouse gas emissions and global warming (Ault et al., 2020). In particular, drought in early spring and summer can be fatal to passion fruit plants. Guizhou Province, located in Southwest China, is characterized by complex terrain and climate conditions, where seasonal drought has gradually become a common phenomenon. Total precipitation levels in Congjiang and Rongjiang counties, the main production areas of passion fruit, amounted to only 70.5 mm and 120.4 mm, respectively, in the third quarter of 2022, showing decreases of 76.6% and 69.5% compared to those of the corresponding period. Persistent high temperatures and little rainfall have resulted in severe agricultural drought, leading to the withering of leaves, developmental arrest of flower buds, as well as fruit wilting and abscission in passion fruit plants. Plants can evolve specific root structures and physiological strategies to respond to complex environments such as drought. For example, they will increase the underground biomass and root distribution space to promote the absorption of more water and fertilizer, thus meeting the requirements for their physiological metabolic activities (Hura et al., 2022). Moreover, they will change the thickness of the leaf palisade and spongy tissues, as well as the number and size of the stomata (Coelho et al., 2022), and will transmit stress signals to promote the synthesis and secretion of proteins with antioxidant enzyme activities to maintain the stability of the plant cell structure (Liu et al., 2022). Therefore, breeding drought-tolerant rootstocks is of great importance to alleviate the decline in the yield and quality of passion fruit caused by drought and other unfavorable conditions.
Passion fruit rootstocks from different regions have complex genetic backgrounds, with substantial differences in growth, development and drought-tolerant physiology. With regard to evaluations of passion fruit rootstocks, the germination characteristics and drought tolerance capacity can serve as important indexes. A higher germination potential can ensure the uniformity of seedlings, which is conducive to synchronous management, while an excessively low germination rate will not only increase seed consumption and management costs but will also delay the farming time, thus negatively affecting income. Better drought resistance ability of rootstocks can assist passion fruit in adapting to different production environments and can provide sufficient water and nutrition for plant growth. However, the commercially available rootstocks in China have mixed provenances and chaotic pedigrees, lacking stability and consistency at present. For this reason, it is urgent to conduct multi-index evaluations to screen rootstocks that have good germination characteristics and preferable suitability for local soil and climate conditions.
In this study, two of the most commonly used rootstocks in China (R1 and R2) and one rootstock from field breeding (R3) are utilized as research materials. Multi-index evaluations were used to compare the seed germination, growth and drought-resistant physiology characteristics among passion fruit rootstocks of the three genotypes, and a comprehensive evaluation incorporating a correlation analysis and the membership function method was implemented, aiming to lay a genetic foundation for cultivating rootstocks suitable for Guizhou and other drought-prone areas.
Materials and Methods
Experimental site and materials
The experiment was carried out on three genotypes of Passiflora edulis f. flavicarpa (R1, R2 and R3) in the Plant Germplasm Resource Conservation Center of the Guizhou Academy of Sciences (26°55′22″N, 106°49′25″E, Elevation: 1051.3 m) in October of 2022. Among them, R1 was purchased from Taiwan (China), R2 was collected from Guangxi (China), and R3 was selected from elite individual plants in Rongjiang, Guizhou (China). Following collection, the seed materials were cleaned, dried, packed in 80-mesh seed bags and stored in a dry and shady room [temperature (T) = 20–25°C and relative humidity (RH) = 40–50%].
Experimental design and treatment
Two experiments, a seed germination experiment and a stress experiment, were carried out in this study. (1) The seed germination experiment was conducted by means of a spot seeding method. Specifically, after soaking, identically sized seeds with high plumpness were selected, disinfected in 75% alcohol for 5 min, and rinsed twice with distilled water. They were then soaked in warm water at 45°C for 30 min, wrapped in wet gauze, placed in a beaker, and transferred to an incubator (T = 30°C, RH = 80%) for 48 h of germination acceleration. The substrate used here was prepared using peat, coconut bran, pearlite and vermiculite at a volume ratio of 2:1:1, fumigated with formaldehyde, and put into pots (13 cm × 15 cm). Thereafter, one seed was sown in one pot, with 100 pots for each genotype. The experiment was repeated three times. (2) The stress experiment was implemented under the conditions of natural drought and potted plants. In brief, for each genotype, 70 pots of plants with consistent growth potential [plant height = (12.5 ± 0.5) cm, and stem diameter = (2.4 ± 0.2) mm] were randomly selected on day 90 after sowing and then watered to reach a saturation moisture level. Subsequently, they were placed in a plastic greenhouse with automatic temperature and humidity control equipment for natural drought conditions (T = 18–25°C, and RH = 50–60%).
Samples and methods
(1) Seed characteristics: Full seeds were randomly selected, followed by a measurement of 1000 seeds weight with a balance, the detection of the seed length and width with Vernier calipers, and the calculation of the seed length-width ratio, with all processes repeated five times. The germination potential and germination rate were calculated on days 20 and 45 after sowing based on the following computational formula:
Formula 1: Seed germination potential/rate = M1 / M × 100%
where M1 represents the number of germinated seeds on days 20 and 45 after sowing and M denotes the number of seeds for the test.
(2) Seedling growth characteristics: A total of 30 individual plants were randomly sampled on day 90 after sowing. Next, Vernier calipers were used to measure the plant height (from the substrate to the top growth point) and stem diameter (2 cm above the substrate), and the number of true leaves was obtained by counting. In addition, the fresh weight of the whole plant, the aerial part and the subterranean part were measured using a balance, and the root-shoot ratio and seedling index were calculated based on Formula 2 (Bai et al., 2014). Additionally, the plant root analyzer (LA-S, WSeen, China) was utilized to measure the total root length, root surface area, root volume, and root tips number of the plants.
Formula 2: Seedling index = stem diameter/plant height × fresh weight × 10
(3) Chlorophyll fluorescence parameters for dark reaction: five individual plants were randomly sampled and subjected to 30 min of dark treatment after stress 21 days. Then, three mature leaves (the second to the fourth leaves from top to bottom) were selected from each test plant to measure the constant fluorescence [initial fluorescence (F0)], maximum fluorescence (Fm) and variable fluorescence (Fv) by a closed multispectral fluorescence imaging system for plants (FC00-C/1010GFP, FluorCam, Czech Republic), after which the maximum light energy conversion rate (Fv/Fm) was calculated.
(4) Physiological response to drought stress: Individual plants (n = 9 per day) were randomly sampled on days 7, 14, 21, 28 and 35 after stress, from which five mature leaves (the second to the sixth leaves from top to bottom) were taken, quickly frozen in liquid nitrogen, and stored in a refrigerator at –80°C.
Hydrogen peroxide (H2O2) was extracted as follows: Leaves (0.5 g) had 1 mL of acetone precooled at 4°C added with 0.2 g of quartz sand and were then ground into a homogenate. After washing, the homogenate was transferred to a centrifugal tube for 10 min of centrifugation at 3,000 r/min. Thereafter, 250 µL of the supernatant was collected, 25 µL of stronger ammonia water and 50 µL of a 5% (W/V) titanium sulfate solution were added, and this was centrifuged at 3,000 r/min for 10 min following precipitation. Afterwards, the supernatant was discarded, while the precipitate was repeatedly washed with acetone four times. 250 µL of a 2 mol/L sulfuric acid solution was added, and this was followed by 5 min of standing at room temperature for subsequent testing.
The extraction of superoxide dismutase (SOD) was done. In brief, leaf tissue (0.5 g) was put into a precooled mortar, with 2 mL of precooled 0.05 mol/L phosphate buffer (pH 7.8) added, and was ground into a homogenate on ice. The homogenate was then washed with a buffer four times, followed by centrifugation at 4,000 r/min for 15 min (precooled to 4°C in advance). Thereafter, the supernatant was collected for subsequent testing.
Catalase (CAT) extraction was implemented as follows: In total, 0.5 g of leaf tissue was taken, 2 mL of 0.2 mol/L phosphate buffer (pH 7.8) and 0.2 g of quartz sand were added for grinding, and this was followed by washing with a buffer four times. Next, the resulting product was transferred to a centrifuge tube for centrifugation at 4,000 r/min for 15 min and the supernatant was harvested for subsequent testing.
Peroxidase (POD) was extracted. Specifically, 0.5 g of leaf tissue was weighed, 2 mL of 0.05 mol/L phosphate buffer (pH 5.5) and 0.2 g of quartz sand were added, and this was ground and then centrifuged at 3,000 r/min for 10 min. The supernatant was then transferred to a 25 mL volumetric flask, and the precipitate was extracted with 5 mL of phosphate buffer twice. Afterwards, all of the supernatant was transferred to the volumetric flask, diluted to 25 mL, and stored at a low temperature for subsequent testing.
The optical density (OD) was measured with a microplate reader (Multiskan SkyHigh, USA) at 450 nm, and the sample concentration was calculated in accordance with the instructions for ELISA kits for plant H2O2, SOD, CAT and POD levels (Shenzhen Ziker Biological Technology Co., Ltd., China).
Statistics and analysis
In this study, the experimental data were statistically analyzed by Excel 2020 and SPSS 22.0 software, and with Duncan’s test (p < 0.05) employed for a variance analysis and a multiple comparison assessment. Spearman’s correlation analysis was conducted using Origin 2021, and a comprehensive evaluation was carried out by the membership function method. For drought resistance indexes with positive and negative correlations, the corresponding membership degrees were calculated according to Formula 3 and Formula 4, respectively. Finally, the drought resistance levels of the different genotypes are characterized by the corresponding average membership value (AMV, Formula 5).
Formula 3: Ri = (Xi–Xmin) / (Xmax–Xmin)
Formula 4: Ri = 1–(Xi–Xmin) / (Xmax–Xmin)
Formula 5: AMV = (∑ Ri) / N
where Ri is the membership degree of the ith index, Xmax and Xmin correspondingly refer to the maximum and minimum values of the ith index of the test material, and N is the total number of indexes used for the evaluation.
Results
Seed morphology and germination
The seeds of rootstocks with the three genotypes were all egg-shaped and dark brown. Among them, the seeds with R1 were zygomorphic, evenly distributed, with small and shallow dimples on the surface, whereas the seeds with R2 and R3 had a metallic surface with concave dimples as well as curved corners at the top germination opening (Fig. 1A). The seed lengths displayed no significant differences among the seeds with R1, R2 and R3, while the seed width of R1 was significantly larger than that of R2 and R3 (Fig. 1B and 1C). The seed length-width ratio was 1.45–1.59, and the seeds with R1 had a significantly decreased length-width ratio in contrast to the seeds with R2 and R3, with respective decreases of 8.81% and 9.36% (Fig. 1D). These results revealed that the seeds with R1 had the greatest 1000-seed weight, which was 23.2% higher than that of the seeds with R2. the R3 exhibited the greatest germination potential of 87.9% and a germination rate as high as 93.3%, while the germination rates of the seeds with R1 and R2 were only 64.4% and 65.6%, respectively (Fig. 1E-1G).
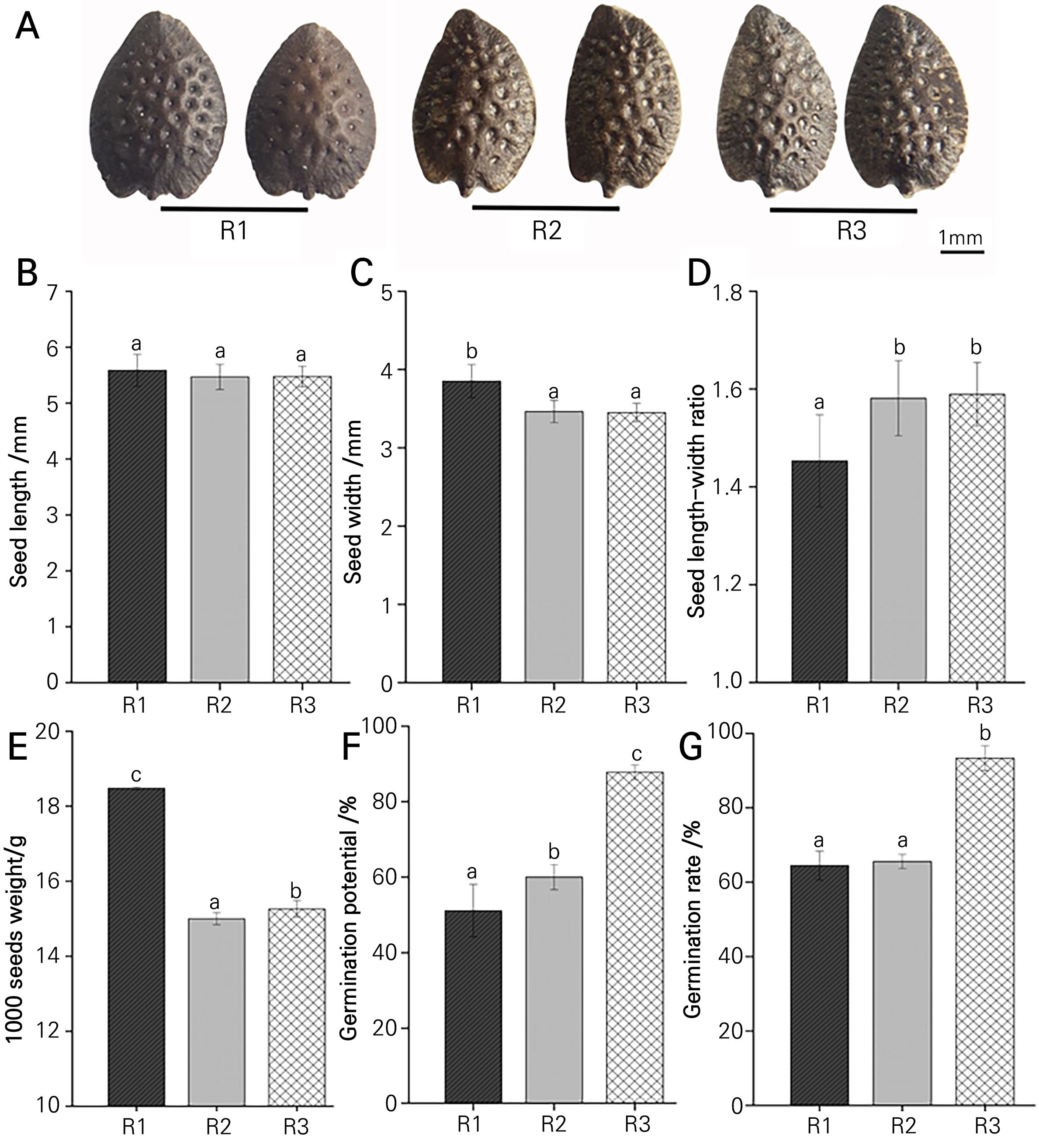
Seedling growth characteristics
The growth indexes of different rootstock seedlings were observed in this research. The plant height, stem diameter, fresh weight of the whole plant and the aerial part, and the seedling indexes of both R1 and R2 were significantly lower than those of R3, with decreases of 10.05% and 13.97% in the plant height, 6.85% and 8.61% in the stem diameter, 13.85% and 15.93% in the fresh weight of the whole plant, and 24.37% and 25.42% in the fresh weight of the aerial part, as well as reductions of 25.68% and 24.00% in seedling index, respectively (Table 1). However, the fresh weight of the subterranean part, the total root length and the root surface area presented no significant differences among the seedlings of the three rootstocks. The number of true leaves was lowest in the seedlings with R1, with obvious decreases of 25.35% and 8.04% compared to those in R2 and R3, respectively. The root-shoot ratio was 0.51–0.65, and it was significantly higher in seedlings with R1 and R2 than in seedlings with R3. It should also be noted that the seedlings with R2 had more abundant roots, reaching 1,107.14 in number, an outcome that was 2.82 and 2.51 times those in the seedlings with R1 and R3, respectively. However, by enlarging the root length and thickness, the seedlings with R1 and R3 had the same root surface area and root volume as the seedlings with R2.
Table 1.
Growth and development characteristics of three passion fruit rootstock seedlings on the 90th day after sowing
| Genotype | PH/mm | SD/mm | LN/N | TFW/g | SFW/g | RFW/g |
| R1 | 125.38 ± 10.09 b | 2.48 ± 0.20 b | 7.18 ± 0.98 b | 3.90 ± 0.70 b | 2.38 ± 0.47 b | 1.52 ± 0.31 a |
| R2 | 121.07 ± 11.70 b | 2.44 ± 0.14 b | 8.33 ± 0.65 a | 3.83 ± 0.57 b | 2.36 ± 0.36 b | 1.47 ± 0.28 a |
| R3 | 137.98 ± 10.38 a | 2.65 ± 0.18 a | 9.00 ± 0.74 a | 4.44 ± 0.62 a | 2.96 ± 0.44 a | 1.48 ± 0.28 a |
| Genotype | RSR | SI | TRL/cm | RSA/cm2 | TRV/cm3 | RT/N |
| R1 | 0.65 ± 0.13 a | 0.74 ± 0.07 b | 440.37 ± 87.06 a | 235.42 ± 38.26 a | 13.50 ± 2.43 b | 393.29 ± 85.98 b |
| R2 | 0.63 ± 0.09 a | 0.75 ± 0.08 b | 496.95 ± 91.24 a | 245.84 ± 47.55 a | 14.40 ± 2.13 ab | 1107.14 ± 238.95 a |
| R3 | 0.51 ± 0.09 b | 0.93 ± 0.15 a | 417.29 ± 84.52 a | 236.61 ± 47.50 a | 16.72 ± 3.45 a | 440.90 ± 82.68 b |
The table shows the mean ± SD of the test indicators, where identical lowercase letters in the column indicate no significant difference between the variables according to Duncan’s test at a probability of 5%. PH: plant height; SD: stem diameter; LN: leaf number; TFW: total fresh weight; SFW: shoot fresh weight; RFW: root fresh weight; RSR: root-shoot ratio; SI: seedling index; TRL: total root length; RSA: root surface area; TRV: total root volume; RT: root tips.
Chlorophyll fluorescence parameters of different passion fruit rootstocks to drought stress
An obvious difference was found in the leaf morphology among passion fruit rootstocks with the three genotypes. Leaves with R1 were broadly ovate with a wide bottom, those with R2 were lanceolate with a large aspect ratio, and those with R3 were oval with a wider middle part (Fig. 2A). Following drought stress, stress symptoms were first observed on the leaf edges and veins of the seedlings. However, after the drought treatment, except for the Fv/Fm ratio, which was markedly higher in the seedlings with R2 than in the seedlings with R1, the F0, Fm and Fv outcomes showed no significant differences among the seedlings of the three rootstocks (Fig. 2B-2E).
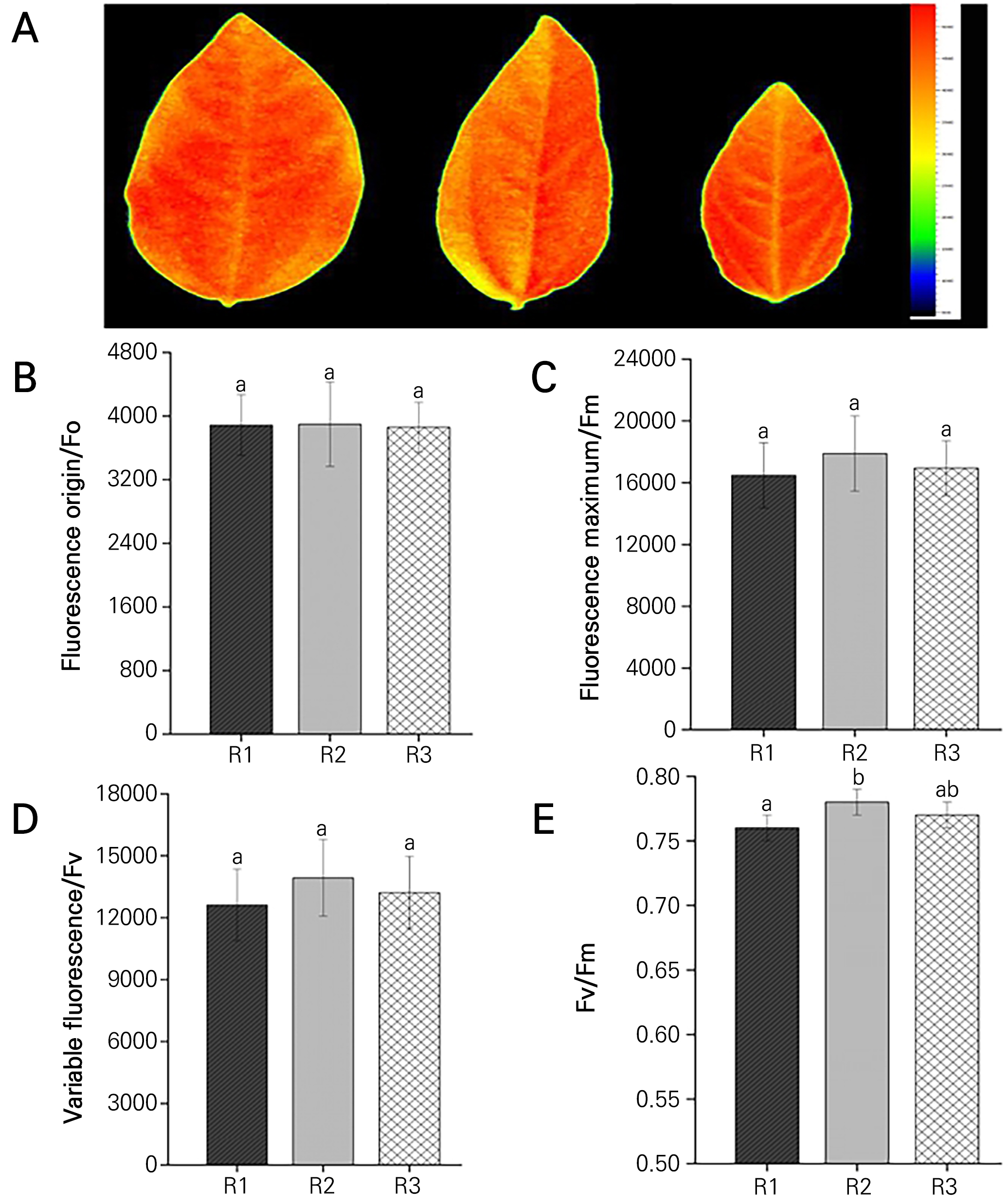
Antioxidant enzyme activities of different passion fruit rootstock seedlings to drought stress
With an increase in the number of stress days, the content of H2O2 in the seedling leaves of the three passion fruit rootstocks showed a trend of first decreasing and then increasing. Then, the H2O2 of R3 reached the lowest point at the 3rd week, only 0.41 µmol/g, which was significantly lower than that at the 1st, 2nd, 4th and 5th week (Fig. 3A). SOD is able to catalyze superoxide anion radicals to generate O2 and H2O2, which play a vital role in the antioxidant balance of plants. Under drought stress, SOD in the leaves of the different passion fruit rootstocks showed a change trend identical to that of H2O2 (Fig. 3B), signifying that SOD contributes much to H2O2 synthesis in plant leaves after stress. CAT increased first and then decreased, with the peak value detected in week 4 (Fig. 3C). POD reached the lowest level in week 4 after stress, which was significantly lower than that in week 1, but the POD activity showed no significant differences among the rootstock seedlings with different genotypes after the same number of stress days (Fig. 3D).
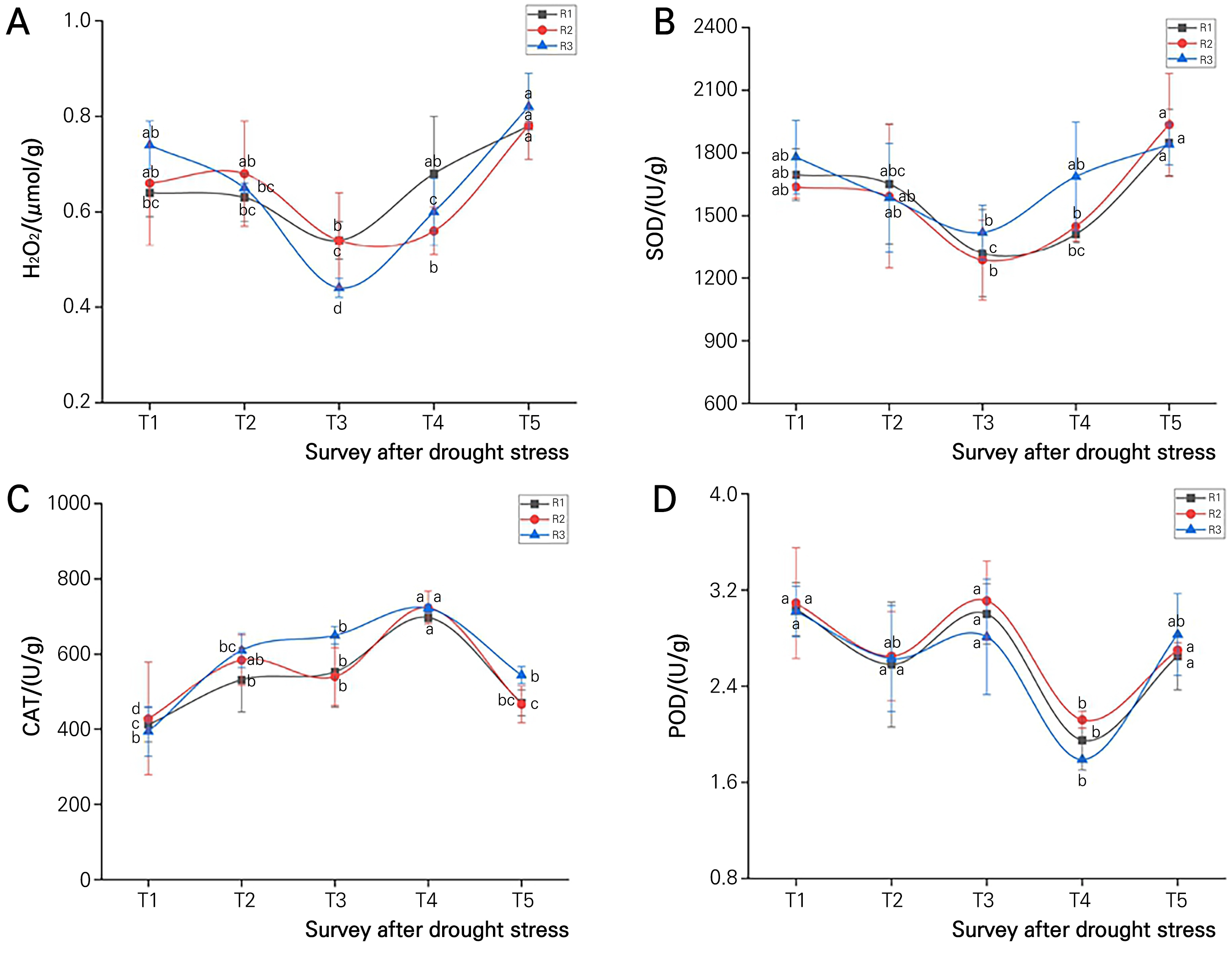
Fig. 3.
Changes in the H2O2 content and antioxidant enzyme activities in different passion fruit rootstock seedlings under drought stress. T1-T5 correspond to weeks 1, 2, 3, 4 and 5 under plant stress. A, B, C and D denote the changes of H2O2, SOD, CAT and POD with the duration of drought in three passion fruit rootstocks. Identical letters indicate no significant differences between the variables according to Duncan’s test at a probability of 5%.
Evaluation of the comprehensive characteristics of different passion fruit rootstocks
A correlation analysis of the growth characteristics and photosynthetic physiology of the passion fruit rootstock seedlings was conducted. The results showed that the plant height had an extremely significantly positive correlation with the stem diameter, fresh weight of the whole plant and the aerial part (Fig. 4), a markedly positive association with the number of leaves, the root fresh weight, and the total root volume, and a significantly negative correlation with the root-shoot ratio (p < 0.05). The seedling index displayed an extremely significantly positive correlation with the fresh weight of the whole plant, the fresh weight of the subterranean part, and the root-shoot ratio, meaning that it can serve as an important reference index of seedling quality. The plant root length exhibited a significantly positive relationship with the root surface area and root volume. Increases in the root length and volume are conducive to an increased soil contact surface, allowing the plant to obtain more space for nutrition and water absorption. Meanwhile, the number of root tips was positively correlated with Fm and Fv, and an abundance of root tips was beneficial to reduce the effect of the drought environment on the photosynthetic physiology of the plants. It should be noted that the Fv/Fm ratio was positively associated with the stem diameter and root length of the plants. It was therefore speculated that the physiological basis for passion fruit rootstocks to obtain drought tolerance may be a larger stem diameter and a longer root length. The comprehensive potential of the passion fruit rootstocks with the three different genotypes was evaluated by the membership function method, which can reflect the growth and development of plants as well as their adaptability to drought. The comprehensive potential outcomes of the passion fruit rootstocks with the three genotypes were ranked based on the average membership (Table 2), which was R3 > R2 > R1 in order of priority. Therefore, R3 can be considered as a rootstock resource suitable for development and utilization in Guizhou and other arid areas.
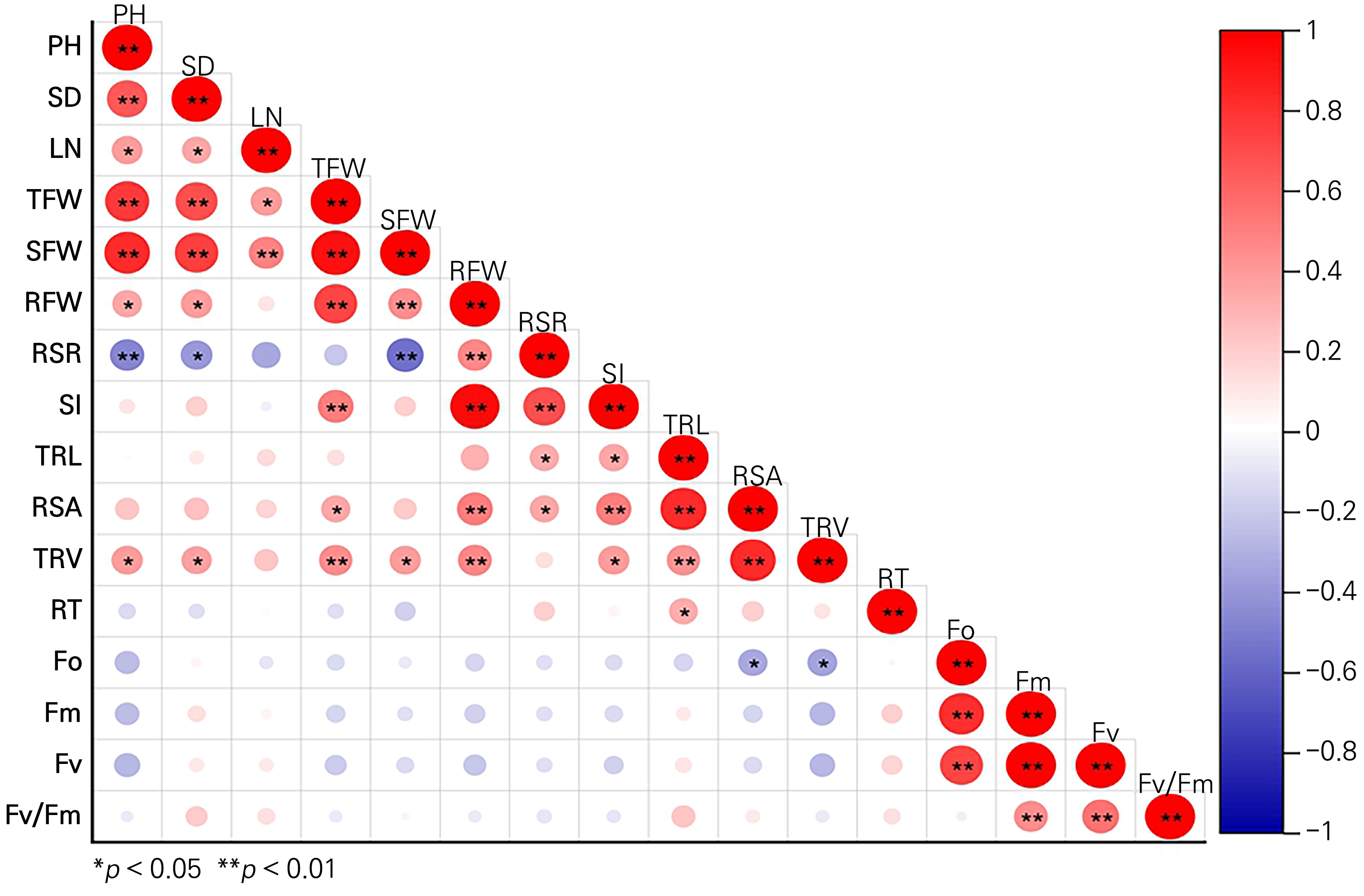
Fig. 4.
Correlation analysis of seedling growth and development of different passion fruit rootstocks. Spearman’s method is used for the correlation analysis, where * denotes significance (p < 0.05) and ** denotes extreme significance (p < 0.01). PH: plant height; SD: stem diameter; LN: leaf number; TFW: total fresh weight; SFW: shoot fresh weight; RFW: root fresh weight; RSR: root-shoot ratio; SI: seedling index; TRL: total root length; RSA: root surface area; TRV: total root volume; RT: root tips.
Discussion
The seeds of three passion fruit rootstocks showed great differences in the appearance and morphology (Fig. 1). To be specific, seeds with R2 and R3 showed a closer genetic relationship, most likely attributable to the long-term geographical isolation of R1, which came from an island area in eastern China. The ranges of the vertical and horizontal diameters of the seeds of the passion fruit rootstocks with the three genotypes were 5.47–5.58 mm and 3.45–3.85 mm, respectively, in line with the results of the third group in research on the germplasm quantitative characteristics of 35 portions of passion fruit conducted by Xu (Xu et al., 2022), whereas these values were lower than those from a study on the seed characteristics of commodity fruits by Wang (Wang et al., 2023a), with decreases of 19.4% (R1), 22.0% (R2) and 21.0% (R3) in the vertical diameter. The 1000-seed weight reflects the size and plumpness of seeds, serving as an important index to test the quality of seeds. It was found that the range of the relative standard deviation of the 1000-seed weight of the rootstock seeds with the three genotypes was 0.24–1.44%, meeting the requirements of seed production standards. This result is slightly lower than that reported by Veiga-Barbosa (Veiga-Barbosa et al., 2013), which is ascribed to the seed production conditions, storage time, drying time and preservation environment. Maldevelopment, overly long storage and drying times, and overly high preservation temperatures of seeds will all reduce the 1000-seed weight. The experimental results showed that the germination rate of rootstock seeds with the three genotypes exceeded 60%, greater than those of P. alata, P. cincinnata, P. coriacea, P. foetida, P. giberti, P. morifolia, P. mucronata and P. nitida. It was also reported that the germination rate can be further elevated by damaging the seed shell with liquid nitrogen, concentrated sulfuric acid, and other such substances (Kakihara and Hondo, 2013), but further studies should be conducted on the effects of the degree of damage on the germination rates of passion fruit seeds.
Rootstocks are characterized by easy reproduction, a high growth potential, a developed root system and strong stress resistance. Passion fruit rootstocks are mainly reproduced through seeds (Rosado et al., 2020), and each fruit usually contains over 100 seeds, showing strong reproductive ability. In this study, the growth, development and root system of seedlings of passion fruit rootstocks with the three genotypes were mainly compared. The results revealed that seedlings with R3 had significant advantages in terms of the plant height, stem diameter, number of leaves and the aboveground biomass (Table 1). seedlings with R2 had a stronger root meristem ability, gaining more water and nutrients by increasing the number of root tips and thereby impeding the metabolic disorder caused by drought and other adverse environments and avoiding such phenomena as wilting of the leaves, developmental arrest of flower buds, and dropping of flowers and fruits. The root system, the main organ for absorbing water, plays a vital role in the overall water balance of plants. Elongation of the root system will be repressed by drought and other adverse situations. However, there are considerable differences in the root length among different species of Passiflora L. For instance, Lima discovered that the total root length of P. mucronate, a local passion fruit resource in Brazilian, can reach 30 m in a hypersaline environment (Lima et al., 2019), a length six times than that of R2. This is attributable to long-term domestication under adverse situations, such as drought, high salt levels, high temperatures and fertilizer deficiencies. It also shows that the stress tolerance of plants can be improved through continuous stress and screening activities.
As a raw material of plant photosynthesis, water accounts for only 1% of root absorption and leaf transpiration. A lack of water in plants will reduce the stomatal opening and thus affect CO2 absorption, increase the hydrolase activity, reduce sugar accumulation, and destroy the photosynthetic mechanism to reduce electron transfers, thus affecting plant photosynthesis. Chlorophyll fluorescence is an important index reflecting plant photosynthetic ability. The complete set of the fluorescence parameters of leaves can be obtained using a fluorescence imager, without causing any damage to plants. Accordingly, this device is widely used for abiotic stress evaluations of plants (Banks, 2017; Baker and Rosenqvist, 2004). An uneven PSII running efficiency was found in passion fruit rootstocks with the three genotypes upon exposure to the drought environment, especially in the leaf veins and edges. This is attributed to the fact that the chlorophyll content differs between the leaf veins and mesophyll cells. At the same time, there is accelerated water loss in the leaf edges of passion fruit with more processes distributed, giving rise to cell dehydration and affecting the photosynthetic efficiency. It is also important to note that no significant differences were observed in F0, Fm and Fv among the three genotypes, whereas the Fv/Fm ratio was significantly higher in R2 than that in R1, implying that R1 underwent obvious membrane damage during drought stress compared to R2, possibly leading to thylakoid dysfunction in these plants (Gomes et al., 2012).
As one of the important indicators of reactive oxygen species (ROS), H2O2 can be produced during such physiological processes as photorespiration, fatty acid metabolism and plant senescence, while it will be rapidly synthesized and decomposed due to certain environmental stimuli, especially during drought and other abiotic stress (Manzer et al., 2015). The results of this study showed that H2O2 in the leaves of passion fruit rootstocks with the three genotypes presented the same trend, reaching the lowest level in week 3 and then rising rapidly (Fig. 3). This suggests that the initial saturation moisture may cause a certain degree of waterlogging and that the soil moisture content is suitable for the growth of passion fruit after two weeks of natural evaporation, whereas H2O2 is synthesized rapidly in leaves with an increase in the number of drought days. The climate phenomenon of waterlogging to drought is especially common in Southwest China. As a contributor to the volume of underground parts, rootstocks should have the ability to adapt to various adverse situations. As previously reported, exceeding the total oxidation potential of cells will induce oxidative stress, affecting plant growth (Qi et al., 2023). To reduce the oxidative damage of ROS to mitochondria, chloroplasts, nuclei and other membrane structures of organelles, plants have evolved a set of antioxidant systems to remove ROS, with SOD, CAT and POD as the most typical representatives. SOD can generate O2 and H2O2 by catalyzing superoxide anion radicals, which play a crucial role in the antioxidant balance of plants. Results have revealed that SOD was highly consistent with the level of H2O2 in terms of the change trend, indicating that SOD in passion fruit rootstocks may make a very useful contribution to H2O2 synthesis after drought stress. It should also be noted that H2O2 in leaves can be accurately regulated through the complex regulation of CAT mRNA (Luna et al., 2004) and that H2O2 can reduce the activity of POD, resulting in the accumulation of more H2O2 in plants (Song et al., 2016). The results here also confirmed that the activity of CAT and POD was reduced after the drought stress treatment, leading to the continuous accumulation of H2O2 and thereby causing irreversible damage to the plants.
A correlation analysis also was conducted to characterize the correlations among the variables (Schober et al., 2018), finding that the plant height of the passion fruit seedlings exhibited a significantly positive correlation with the stem diameter, number of leaves, the biomass (fresh weight of the whole plant, the aerial part, and the subterranean part) and the root volume, consistent with the growth and development of C3 plants. However, it is important to note that the seedlings with R3 had the smallest root-shoot ratio but displayed more prominent performance in terms of the plant height, stem diameter, number of leaves, fresh weight of the whole plant, fresh weight of the subterranean part, and the seedling index. It was hence speculated that R3 has a superior water absorption rate and better utilization efficiency. A comprehensive analysis can be done using the membership function evaluation method combined with multiple indicators, avoiding the one-sidedness of a single indicator, which has been applied to the assessments of the stress resistance and germplasm characteristics of passion fruit, grapes, kiwifruit and others (Dong et al., 2015; Chen et al., 2022; Wang et al., 2022). According to a report by Dong et al. (2015), passion fruit can be divided into a high-resistance type (AMV≥0.55), a medium-resistance type (0.45≤AMV≥0.55) and a low-resistance type (AMV≤0.45) based on the average membership degree. Therefore, R3 could be considered as a drought-resistant germplasm resource (Table 2).
Table 2.
Membership function values of different genotypes of three passion fruit rootstocks
| Genotype | PH | SD | LN | TFW | SFW | RFW | RSR | TRL | RSA | TRV |
| R1 | 0.000 | 0.000 | 0.000 | 0.000 | 0.253 | 0.253 | 1.000 | 0.000 | 0.000 | 0.000 |
| R2 | 0.901 | 0.237 | 0.905 | 0.095 | 0.000 | 0.000 | 0.730 | 1.000 | 0.529 | 0.581 |
| R3 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 0.000 | 0.578 | 1.000 | 1.000 |
| Genotype | RT | F0 | Fm | Fv | Fv/Fm | H2O2 | SOD | CAT | POD | AMV |
| R1 | 0.016 | 0.692 | 1.000 | 1.000 | 1.000 | 0.021 | 0.250 | 0.326 | 1.000 | 0.359 |
| R2 | 1.000 | 1.000 | 0.500 | 0.500 | 0.500 | 0.000 | 0.000 | 0.000 | 0.500 | 0.473 |
| R3 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1.000 | 1.000 | 1.000 | 0.000 | 0.609 |
The study found that there is a large difference in the affinity between different rootstocks and cuttings, which may affect the yield and quality (Karimi and Nowrozy, 2017). Among them, the incompatibility of rootstocks to cuttings easily triggers an impedance reaction, resulting in poor connections between the conducting tissues and vascular bundles and causing disordered nutrient absorption. In addition, another important function of passion fruit rootstocks is to resist damage from stem rot, a type of soil-borne disease. Therefore, using R3 as a drought-resistant parent, various excellent rootstock materials can be obtained through hybridization, molecular breeding, and other means.
Conclusion
Differences were found in the seed morphology, germination characteristics, growth and development, chlorophyll fluorescence parameters and antioxidant enzyme activities among passion fruit rootstocks from different sources and backgrounds. The results of this study showed that the passion fruit rootstock of R3 is a genotype with high drought resistance and that it may also possess a more efficient water metabolic capacity. Hence, R3 can serve as a drought-tolerant resource in breeding to acquire new cultivars with useful drought tolerance, high yields, good disease resistance and other excellent characteristics.
