

서 언
식물의 종자로부터 유묘로 발달하는 과정은 향후 개체군의 생존과 정착을 결정하기 때문에 생활사에서 매우 중요한 단계이다(Harper, 1977). 종자가 발아하고 유묘가 출현하는 시점은 종 특이적인데, 이는 식물의 생활환, 서식지의 조건 및 지리적인 분포에 따라 다르다(Baskin and Baskin, 1998; Nikolaeva, 1999). 종자가 모식물체에서 탈리되어 유묘가 출현하기까지는 일정기간이 필요한데, 이 기간 동안 자연조건에 많은 영향을 받는다. 따라서 종자는 다양한 휴면습성을 가지며, 이는 유묘의 생존 확률을 높일 수 있는 중요한 전략이다(Baskin and Baskin, 1998).
온대지방에 자생하는 상당수의 숙근성 식물 종자는 배가 충분히 성숙하지 못한 상태로 모체에서 탈리되는 것으로 알려져 있다(Martin, 1946). 이러한 식물의 종자는 배가 발아하기 전에 일정 크기 이상 자라야 하기 때문에 미숙배라고 지칭한다(Baskin and Baskin, 1998; Walk et al., 1999). 그러나 Drosera anglica의 경우, 종자의 크기에 비해 배의 크기는 작지만, 유근이 돌출하기 전에 추가적인 배의 신장은 일어나지 않기 때문에 완전히 성숙한 배로 판단한다(Baskin and Baskin, 2005).
미숙배 종자의 경우 적절한 환경조건에서 배의 신장과 유근의 돌출이 30일 이내에 이루어지는 경우를 형태적 휴면(morphological dormancy, MD)이라 한다(Baskin and Baskin, 1998; Walk et al., 1999). 그러나 배의 신장과 유근이 돌출되기까지 1개월 이상의 시간이 소요되고, 저온습윤, 고온습윤 또는 복합처리를 해야만 발아가 촉진되는 유형을 형태생리적 휴면(morphophysiological dormancy, MPD)이라 한다. 형태생리적 휴면은 첫째, 종자휴면을 타파시키고 발아를 촉진시키기 위한 온도, 둘째, 배가 발달하는 동안의 온도, 셋째, gibberellic acid(GA)의 휴면타파 여부에 따라 여덟 가지로 분류할 수 있다(Baskin and Baskin, 1998, 2004).
온대지방에 자생하는 많은 백합과(Liliaceae)와 미나리아재비과(Ranunculaceae) 식물들은 미숙배 종자를 탈리시키는 것으로 알려져 있다(Baskin and Baskin, 1998; Kondo et al., 2002; Martin, 1946; Niimi et al., 2006). 백합과의 Erythronium japonicum(Kondo et al., 2002), E. grandiflorum(Martin, 1946), Trillium camschatcense(Kondo et al., 2011), 그리고 미나리아재비과의 Helleborus niger(Niimi et al., 2006), Hepatica nobilis(Nomizu et al., 2004), Aconitum lycoctonum (Vandelook et al., 2007) 등의 종자는 미숙배로 탈리되고 형태생리적 휴면을 나타낸다. 그러나 한반도 자생종의 경우, 형태적 종자휴면에 관한 연구는 거의 없었는데, 최근에 와서야 모데미풀(Megaleranthis saniculifolia), 깽깽이풀(Jeffersonia dubia), 복수초(Adonis amurensis), 한계령풀(Leontice microrhyncha) 등에서 이에 대한 연구가 진행되고 있다(Lee et al., 2003, 2011, 2012; Rhie et al., 2010). Heloniopsis와 Thalictrum속의 경우 종자의 내부 형태와 휴면에 관한 구체적인 문헌이 거의 없다. Baskin and Baskin(1998)과 Copete et al.(2011)이 Takahashi(1984)의 문헌에 근거하여 Heloniopsis orientalis가 형태적 휴면(MD)이라 하였다. Thalictrum속의 경우 미국에 자생하는 Thalictrum mirabile의 종자가 미숙배로 탈리되며, 수개월 동안 발아가 지연되는 형태생리적 휴면(MPD)이라고 보고되었다(Walk et al., 1999).
남한에 자생하는 Heloniopsis속 식물로는 처녀치마(Heloniopsis koreana)와 숙은처녀치마(H. tubiflora)가 대표적인데, 두 종 모두 특산식물로 분류되어 있다(Fuse et al., 2004). 처녀치마는 산속의 습한 음지에서 자라는 상록성 다년초로서 이른 봄에 개화하는 것이 특징이며, 지상부의 잎이 늦가을까지 녹색을 유지하여 관상용, 지피용으로 유망한 자생식물이다. Thalictrum속의 경우 남한에 약 16종이 자생하고 있는데(Jeon et al., 2007), 이 중 금꿩의다리(Thalictrum rochenbrunianum)는 관상용으로 재배되고 있다. 또한 자주꿩의다리(T. uchiyamai)는 특산식물이고 연잎꿩의다리(T. coreana)는 멸종위기 식물로 지정되어 있다(Hammer and Khoshbakht, 2005; Korea National Arboretum, 2012). 따라서 본 연구에서는 몇 가지 자생 관상용 Heloniopsis와 Thalictrum속 식물들을 대상으로 첫째, 종자가 형태적으로 미숙한지, 둘째, 만약 미숙배 종자라면 어떠한 형태인지, 셋째, 형태적 휴면 또는 형태생리적 휴면 중 어떤 휴면 유형을 보이는지를 조사하여, 추후 이러한 종자휴면의 생태・생리적 연구를 위한 기초자료를 제공하고자 수행하였다.
재료 및 방법
종자 채종
백합과의 처녀치마, 미나리아재배과의 금꿩의다리, 자주꿩의다리 및 연잎꿩의다리 종자는 경기도 용인소재 한택식물원에서 각각 2012년 5월 20일, 9월 20일, 9월 30일, 9월 19일에 채종하였다. 숙은처녀치마 종자는 전라북도 무주에 위치한 덕유산 향적봉에서 2012년 6월 18일에 채종하였다. 채종한 종자 또는 과실은 지퍼백에 담아 실험실(25-30°C, 20-30µmol・s-1・m-2)로 가져와 7-15일 정도 건조, 후숙과정을 거친 후 5°C cold-lab chamber(DS-91, Dasol Scientific Co., Hwaseong, Korea)에 실험에 사용될 때까지 건조 보관하였다. 보관하던 종자를 꺼내어 충실한 종자들을 정선하여 실험에 사용하였다.
종자의 내・외부 형태 관찰
채종한 종자들은 곧바로 실험실로 가져와 외부사진을 촬영하였다. 채종 약 보름 후에 충실한 종자를 반으로 잘라 해부현미경(KSZ-1B, Samwon Scientific Co., Ltd., Seoul, Korea)으로 내부 형태를 관찰하였다. 반으로 자른 종자를 해부현미경의 작업대에 올려놓고 micrometer가 장착된 접안렌즈로 관찰하면서 종자와 배의 길이를 측정하였다. 종자와 배의 길이를 측정한 후 miview USB digital microscope (MV 1302U, CosView Technologies Co., Ltd., Shenzhen, China)를 이용해 60-200배율로 종자 내부를 촬영하였다. 종자 내부의 배의 크기는 종자의 크기가 작을수록 상대적으로 작은 경우가 많다. 따라서 배의 발달은 종자 내부의 배유조직에 대한 배의 상대적인 크기로 표현해야 정확하다 할 수 있다. 본 연구에서는 미숙배의 신장을 좀더 명확하게 표현할 수 있는 배(embryo)와 종자의(seed)의 비율인 E:S ratio (Vandelook et al., 2007)를 계산하였다. 특히 연구 대상종 중 꿩의다리속 종자는 수과(achene)로서 과피 내부의 실제 종자부위를 종자의 길이로 측정하였다. 또한 미숙배인지 여부를 판단하기 위해서 유근이 돌출되기 직전에 배의 길이를 측정하였다.
발아실험
정선한 종자들을 90 × 15mm disposable petri dish에 여과지(Whatman No. 2) 2매를 깔고 benomyl 수화제(500mg・L-1)를 분무하여 약 1-2시간 정도 소독하였다. 소독 후 증류수로 세척한 후 30립 3반복 완전임의 배치하여 multi-room incubator (DS-13MCLP, Dasol Scientific Co., Ltd., Hwaseong, Korea)에서 배양하였다. 배양환경은 주・야 25/15°C(12h/12h), 광도는 약 20µmol・s-1・m-2였다. 배양 중에 곰팡이에 오염된 부위가 발견될 때마다 petri dish를 꺼내어 benomyl 수화제로 소독하였다. 배양 후 일주일 간격으로 총 8주 동안 발아율을 조사하였고, 조사 시 부패하여 미발아된 종자들은 발아율 계산에서 제외하였다. 배양 중 처녀치마, 숙은처녀치마, 금꿩의다리, 자주꿩의다리 및 연잎꿩의다리 종자의 부패율은 각각 40%, 17%, 3%, 30% 및 8% 정도였다.
결과 및 고찰
백합과의 처녀치마와 숙은처녀치마는 종자의 형태와 크기가 비슷하였다(Fig. 1). 미나리아재비과의 금꿩의다리, 자주꿩의다리 및 연꿩의다리는 모두 수과(achene)로서 과피가 종자를 둘러싸고 있었다. 꿩의다리 세 종의 경우 종자의 크기와 과피의 모양은 달랐으나 과피 내부의 종자는 비슷한 모양이었다. 그러나 연구 대상 다섯 종 모두 배의 크기가 작다는 것을 확인할 수 있었다. Martin(1946)은 종자를 내부 배의 형태에 따라 크게 기저형(basal), 지협형(peripheral, 주변형) 및 중축형(axile)의 세 가지 분류군으로 나누었다. 일반적으로 기저형 종자는 배가 작고 종자의 하단부에 위치하며, 종자 크기의 50%를 넘지 않는다. 지협형 종자는 배가 길고 크며, 종피를 따라 휘어 있는 형태이다. 중축형 종자는 배가 작은 것에서부터 긴 것까지 다양하지만, 백합과의 경우, 작고 길쭉한 모양의 배가 종자의 중앙부에 위치한 것들이 많다. 기저형에서는 타원형의 작은 배가 종자의 하부에 위치한 미발달형(rudimentary type)을 그리고 중축형에서는 작고 길쭉한 형태의 배가 종자의 중앙부에 위치한 직선형(linear type)을 종자의 대표적인 형태로 보고하였다. 백합과의 처녀치마와 숙은처녀치마는 미발달형 배를 가지고 있었고, 두 종 모두 심장형 배(heart-shaped embryo)보다는 더 발달된 어뢰형 배(torpedo-shaped embryo)를 가지는 것으로 판단된다. 미나리아재비과의 세 종 모두 미발달형 배로 판단되나, 배의 너비에 비해 약간 긴 형태를 보였고 어뢰형 배의 형태를 보였다. 백합과 종자들의 배는 직선형이 대부분인 것으로 알려져 있는데(Martin, 1946), 한반도 특산 처녀치마 두 종은 미발달형 배를 가지고 있어, 몇 안 되는 예외 종으로 판단되었다. 미나리아재비과 종자들은 대부분 미발달형 배를 가지는 것으로 보고되었는데(Martin, 1946), 본 연구의 꿩의다리 세 종은 미발달형과 직선형의 중간형태로 판단되었다.
본 연구에서는 종자의 길이, 너비 및 배의 길이를 조사하였다(Table 1). 또한 같은 종이더라도 종자의 크기가 다양하고 배의 길이도 차이가 있기 때문에, 종자의 길이에 대한 배의 상대적인 크기를 좀더 명확하게 표현할 수 있는 E:S ratio(Vandelook et al., 2007)를 제시하였다. 종자의 길이는 백합과의 처녀치마와 숙은처녀치마, 미나리아재비과의 금꿩의다리가 가장 길었고, 5mm 이상이었다. 그러나 처녀치마와 숙은처녀치마는 종자의 가장자리의 하얀색 날개부위를 제외하면 실제 배유조직을 포함하는 종자부위는 2mm 미만이었고, 종자의 너비도 0.8mm 미만이어서 미세 종자였다. 꿩의다리 세 종 가운데 금꿩의다리 종자가 가장 길었다. 배의 길이는 꿩의다리 세 종이 백합과의 처녀치마 두 종보다 더 길었다. 백합과 두 종의 E:S ratio는 0.10 내외여서 배의 비율이 약 10% 정도로 비슷하였다. 그러나 꿩의다리 세 종 가운데 금꿩의다리와 연잎꿩의다리는 E:S ratio가 0.20 정도여서 배의 배율이 약 20% 정도였지만, 자주꿩의다리는 0.15 이하여서 15% 정도로 꿩의다리 세 종 중에서 가장 작았다. Walk et al.(1999)은 꿩의다리와 동일 속인 Thalictrum mirabile의 종자휴면 연구에서 초기 배의 길이가 약 0.56mm 정도라고 하여 남한의 자생종인 금꿩의다리 및 연잎꿩의다리와 비슷하였으나, 자주꿩의다리의 경우 약 0.3mm 정도였기 때문에 같은 속이라도 종마다 배유 내에서 배가 차지하는 영역이 다양함을 알 수 있었다.
|
Fig. 1. Morphology of flowers and seeds in two genera (Heloniopsis and Thalictrum) native to Korea. emb, embryo; end, endosperm; per, pericarp. |

발아하기 직전의 배의 길이를 조사하였다(Table 1). 백합과의 처녀치마 두 종 모두 배가 일정 크기까지 신장한 후 유근이 돌출되었다(결과 미제시). 미나리아재비과의 금꿩의다리, 자주꿩의다리 및 연잎꿩의다리의 배는 초기 배의 길이에 비해 각각 270%, 500% 및 277% 정도 증가되었다(Table 1). 꿩의다리 세 종 모두 종자 길이의 70% 정도까지 배가 신장한 후 발아하였다. 따라서 본 연구의 5종 모두 미숙배인 것을 확인하였고, 이는 발아하기까지 일정시간을 지연시키는 요인으로서 형태적 종자휴면을 보이는 것으로 판단되었다. 미국 자생종인 T. mirabile 종자도 발아 전에 미숙배가 약 300% 정도 신장했다고 하여, 본 연구 결과를 뒷받침 해준다(Walk et al., 1999).
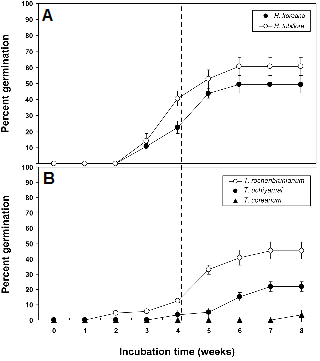
파종 후 8주 동안의 발아율은 연구대상 5종 간에 극명한 차이를 보였다(Fig. 2). 처녀치마 두 종은 2주까지 발아하지 않다가 3주째부터 발아하기 시작하였고(Fig. 2A), 이어 4주째 각각 22.7%, 40.7%, 8주째는 각각 49.4%, 60.7% 발아하였다. 꿩의다리 세 종은 차이가 컸는데, 금꿩의다리는 2주째부터 발아가 시작되었고, 4주째 12.5%, 8주째 45.4% 발아하였다(Fig. 2B). 자주꿩의다리는 4주째부터 발아하였고, 8주째 21.9% 발아하였다. 연잎꿩의다리는 7주까지 발아하지 않다가 8주째가 되어서야 3.3% 발아하여 가장 저조하였다. Baskin and Baskin(2004)의 분류체계에 따르면, 미성숙 배를 가지는 종자들은 형태적 휴면(MD) 또는 형태생리적 휴면(MPD)을 보이는데, 아무런 휴면타파 처리 없이 30일 내에 배의 신장과 발아가 이루어지는 유형을 형태적 휴면이라 하고, 저온습윤, 고온습윤 또는 복합처리를 해야만 배가 신장하고 발아되는 유형을 형태적인 휴면과 생리적인 발아억제 요인이 복합적으로 작용한다 하여 형태생리적 휴면이라 하였다(Baskin and Baskin, 1998, 2004; Walk et al., 1999). 따라서 미숙배 종자의 발아소요일수가 약 30일 이상일 경우, 형태생리적 휴면으로 판단한다.
본 연구에서 처녀치마와 숙은처녀치마는 미성숙 배를 가지고 있지만, 30일 내에 각각 22.7%, 40.7%가 발아하였기 때문에(Fig. 2A), 각 종자의 약 22%, 40% 정도가 형태적 휴면으로 판단되었다. 따라서 두 종의 나머지 약 78%, 60% 정도는 형태생리적 휴면으로 판단되었다. 일본 자생종인 Heloniopsis orientalis의 경우, 30일 내에 거의 대부분의 종자가 발아하였다고 보고되었다(Copete et al., 2011; Takahashi, 1984). 따라서 자생지의 환경과 지리적인 위치에 따라서 휴면성의 깊이는 차이가 있음을 확인할 수 있었다.
금꿩의다리, 자주꿩의다리 및 연잎꿩의다리의 종자는 30일 동안 각각 12.5%, 3.3% 및 0%가 발아하였기 때문에(Fig. 2B), 종자의 약 87%, 96% 및 100%가 형태생리적 휴면임을 알 수 있었다. 따라서 미숙배가 발아지연의 첫 번째 원인이며, 추가적인 생리적 요인으로 인해 발아가 다소 지연되는 것으로 보인다. 또한 한 종 내에서도 형태적 휴면과 형태생리적 휴면을 동시에 보였는데, 이는 개체군 내에서 종자마다 휴면의 깊이를 달리하여 적절한 환경이 주어져도 발아소요일수를 달리하기 위해 적응한 결과라 할 수 있다(Adams et al., 2005; Baskin and Baskin, 2004). Lee et al.(2012)은 한반도에 자생하는 네 종의 미나리아재비과, 두 종의 백합과의 춘계단명 종들의 종자가 모두 형태적 또는 형태생리적 휴면을 보였다고 보고하였다. 또한 외국종의 경우 미나리아재비과의 Helleborus niger(Niimi et al., 2006), Aconitum lycoctonum L.(Vandelook et al., 2009), 백합과의 Trillium camschatcense(Kondo et al., 2011), Erythronium albidum (Baskin and Baskin, 1985) 등에서도 형태생리적 휴면이 보고되고 있어서 본 연구결과를 뒷받침 해준다.
형태생리적 휴면은 배발달 온도가 상대적 고온인 simple- type MPD와 저온인 complex-type MPD의 두 가지 분류군으로 나눌 수 있는데, 각각의 분류군 내에서도 생리적인 휴면의 깊이에 따라 다시 non-deep MPD, intermediate MPD, 그리고 deep MPD의 세 가지 유형으로 나눌 수 있으며, deep type으로 갈수록 GA의 효과는 떨어진다(Baskin and Baskin, 1998, 2004). 백합과 두 종의 경우 발아는 지연되었으나, 주간 25°C의 상대적 고온에서 발아율이 지속적으로 증가하는 경향이었다(Fig. 2A). 따라서 Heloniopsis속은 simple-type MPD로 판단되었다.
결론적으로 한반도에 자생하는 많은 수의 백합과, 미나리아재비과 종들의 종자는 모식물체에서 탈리되는 시점에 미숙배를 가지고 있는 것으로 판단되었다. 본 연구에서는 다섯 가지 자생종을 대상으로 한 기초특성을 바탕으로 하여 형태적 또는 형태생리적 휴면인지는 분류하였으나, 어떠한 유형의 형태생리적 휴면인지에 관해서는 구체적인 휴면의 생태・생리연구가 이루어져야 할 것으로 판단된다. 또한 발아가 지연되는 원인이 무엇인지, 그리고 촉진시킬 수 있는 방법이 무엇인지에 관해 추가적인 연구가 이루어져야 할 것이다. 지난 수십 년간 국내에서도 많은 숙근성 식물들의 종자휴면에 관해 연구가 진행되어 왔다. 특히, 저온(cold stratification), GA(gibberellic acid) 및 후숙(after-ripening) 처리 등을 통하여 휴면타파되는 생리적 휴면, 종피파상(scarification) 등으로 휴면타파되는 물리적 휴면에 관해서는 많은 연구(Kang et al., 2010; Kim et al., 2010; La and Jeong, 2008; Yeam et al., 1985)가 이루어져 왔으나 성숙한 종자가 미성숙한 배를 가지고, 단순히 저온습윤처리, GA 처리 등을 통해 휴면이 타파되지 않고, 발아가 억제되는 다소 복잡한 생리기작을 보이는 형태생리적 휴면에 관한 연구는 최근에 와서야 깽깽이풀(Rhie et al., 2010), 복수초(Lee et al., 2011), 한계령풀(Lee et al., 2012) 등에서 구체적으로 이루어지고 있다. 따라서 국내 자생종을 대상으로 한 이러한 연구들은 학술적인 측면에서 의의가 있을 뿐만 아니라 원예적인 측면에서 종자번식을 위한 중요한 기초자료가 될 것이다.
