

서 언
재료 및 방법
식물 재료
휴면단계 변화 탐색
휴면 심도에 따른 눈의 형태학적 변화
결과 및 고찰
발아 특성에 따른 휴면 단계 및 심도 판정
발아 특성에 따른 저온요구도 설정
휴면 심도에 따른 눈 발달 관찰
꽃눈과 잎눈의 외적 특성 비교
서 언
대부분의 온대과수는 봄에서 겨울까지의 4계절을 주기로 지속적인 환경변화를 겪으며 생존하고 있다. 그 중, 내재휴면은 계절적으로 식물 생장에 불리한 조건으로 전환되는 중에 생장을 억제하기 위한 눈의 내부적인 생리변화에 의해 나타나는 결과이다(Horvath et al., 2003). 내재휴면의 유도 및 타파에는 일장과 온도가 중요하게 작용하고 있으며, 그 중에서도 일장은 대부분의 다년생 목본식물에서 내재휴면 유도에 더 중요하게 작용한다(Heide, 2008; Okuba, 2000). 반면, M. Domestica나 P. communis와 같은 일부 장미과에 속하는 품종들은 광주기에 둔감하여 이와 관계없이 12°C 이하의 온도에서 내재휴면이 유도된다고 하였으며(Heide and Prestrud, 2005), 이와 더불어 저온의 축적은 내재휴면 타파를 위한 필수 요소이다(Atkinson et al., 2013; Mohamed et al., 2010; Takemura et al., 2013).
저온은 휴면과정뿐만 아니라 이듬해에 수체와 과실의 전반적인 생장에도 영향을 미치기 때문에 저온의 축적 정도를 평가함으로써 휴면심도와 휴면타파 이후에 진행되는 과정들을 예상하고자 하는 연구들이 수행되었다. 또한, 저온 축적의 평가는 발아라는 가시적인 결과를 통해 눈이 가지고 있는 휴면심도를 결정할 수 있는 이점이 있으나, 지금까지 저온 축적 및 발아에 관련된 연구들은 낙엽 직후의 가지를 채취하여 인위적인 저온 처리를 통해 휴면개시 및 타파시기를 간접적으로 추정한 연구들이 대부분이었기 때문에 실제 재배포장의 기온 및 눈의 생리학적 발달 정도를 반영한 방법이 확립되어 있지 못한 상태이다.
이와 같이 온대과수의 휴면에 관한 이전의 연구들은 주로 휴면의 유도와 타파에 요구되는 기상 조건들을 밝히는 데 중점을 두었으며, 최근에는 지구온난화와 관련하여 변화하는 기상 환경에 대한 휴면 기작의 반응 및 예상되는 문제점과 해결 방안 등에 관심이 집중되고 있다. 실제로 지난 133년간(1880-2012년) 연평균 기온이 0.85°C 상승하였고, 향후 지구 온난화에 따른 한반도 기온은 대표농도경로(RCP: Representative Concentration Pathway) 4.5 기준으로 1986-2005년 대비 21세기 중반(2046-2065년)에는 연평균 2.3°C, 21세기 후반(2081- 2100년)에는 3.0°C 상승할 것으로 예측되고 있으며(Chung et al., 2007; IPCC, 2013; KMA, 2012) 이러한 기상 조건의 변화는 국외뿐만 아니라 국내 주요 과수의 생물계절 및 재배적지 변화를 유도하고 있다(Chmielewski et al., 2004; choi et al., 2011). 이전까지의 재배적지에 관한 연구들은 생육기의 재배 적온의 부족으로 인한 북방한계선의 개념이 우선되었으나, 최근에는 휴면타파에 필요한 저온 부족의 가능성으로 인해 과수재배의 남방한계선 설정을 위한 연구들이 새롭게 요구되고 있다.
따라서 본 연구에서는 국내 주요 재배종인 ‘Fuji’ 사과를 대상으로 생육기부터 휴면타파 이후까지 시기를 세분화하여 발아 특성 관찰을 통해 국내 환경에 맞는 휴면단계 변화 및 내재휴면 타파와 저온 사이의 관계를 알아보고자 하였다. 또한, 눈 발달을 관찰하여 발아와 휴면과의 관계를 보다 객관적으로 평가하여 직면한 기후 변화에 따른 변화들을 예상하고 대처하는데 필요한 기초 자료를 제공하고자 하였다.
재료 및 방법
식물 재료
충청북도 충주시 주덕읍 소재 사과원에서 M26과 M7에 이중 접목한 25년생 ‘Fuji’ 사과나무를 대상으로 하였으며, 2011년 7월부터 2012년 2월까지 15일 간격으로 발아 후 당해 년도 형성되는 액아를 가진 1년생 가지 중 20-30cm 정도 자란 가지를 채취하였다. 가지는 재식된 나무 중 수세가 비슷한 20여 나무를 선발하여 시기별 무작위로 20개의 가지를 채취하였으며, 그 안에 포함되어 있는 꽃눈을 사용하였다.
휴면단계 변화 탐색
일반적으로 수체 및 눈의 휴면심도는 수삽을 통한 발아율 및 발아 소요 일수를 조사하는 방법으로 추정하고 있으며(El-Agamy et al., 2001; Kim et al., 2006; Takemura et al., 2013), 절지를 발아에 적합한 조건의 온실 또는 기내에서 발아시킬 때 15일 이내 발아하면 내재휴면이 완료된 것으로 평가하고 있다(Hauagge and Cummins, 1991; Kim et al., 2006). 따라서 본 연구에서도 이와 동일한 방법으로 ‘Fuji’ 사과 품종의 휴면심도를 평가하였다.
채취한 가지에서 눈이 충실하게 발달된 것으로 생각되는 중간 부분을 절단하여 수삽에 사용하였다. 가지에 남아있는 잎을 제거한 뒤, 70% 에탄올로 1차 소독한 후 증류수에 수삽하였으며 증류수는 매일 교환해주었다. 또한 agar 등에 의해 물에 잠긴 기부의 물관부 손상 및 부패 방지를 위하여 7일에 한번씩 기부의 1-1.5mm 정도를 잘라내었다. 발아 조건은 온도 23 ± 2°C, 상대습도 30% 및 광주기 16/8(day/night)로 일정하게 유지시킨 후부터 지속적인 육안관찰을 통해 최초 발아까지 소요된 일수와 시기별 발아율 및 최종 발아율을 조사하였다.
휴면 심도에 따른 눈의 형태학적 변화
가지의 길이는 결과모지로부터 10번째 마디까지의 평균 길이를 산출하였고, 가지의 굵기는 각 마디 사이의 직경을 버니어 캘리퍼스(CD-15CP, Mitutoyo Corp., Tokyo, Japan)를 이용하여 측정하였다. 눈의 크기는 기부의 첫 번째 눈부터 열 번째 마디의 눈까지 종경과 횡경을 각각 측정하였으며, 예리한 칼(ST-300 stainless blade, Dorco, Korea)을 이용하여 액아를 종축으로 절단한 뒤, 실체 현미경(SZX10, Olympus, Tokyo, Japan) 관찰을 통해 시기별로 눈의 발달 양상 및 괴사유무를 조사하였다. 또한 가지의 종류에 따라 과대지(bourse shoot)의 액아와 단과지(spur)의 정아로 구분하여 각각 형태에 따른 눈의 꽃눈 분화 여부를 구분하였다.
통계분석은 PASW Statistics 18(SPSS Inc., USA) 프로그램을 이용하여 수집된 데이터의 Duncan 다중검정(p = 0.05) 및 T-검정을 실시하였다.
결과 및 고찰
발아 특성에 따른 휴면 단계 및 심도 판정
만개 후 150일과 165일에 채취한 가지에서 각각 수삽 후 7일과 15일부터 액아의 일부가 발아를 시작하였으며, 만개 후 165일의 경우에는 최종 발아율 또한 70%로 높게 조사되었다(Table 1). 기존의 보고에 따르면 발아 소요 일수가 15일을 초과하는 경우를 내재 휴면 기간으로 정의하고 있는데(Hauagge and Cummins, 1991; Kim et al., 2006), 만개 후 165일까지는 모두 15일 이내에 발아를 시작하였고, 발아율 또한 높게 유지되어 이 시기는 내재휴면이 시작되기 전인 것으로 판단되었다. 만개 후 180일에는 최초 발아까지 소요되는 기간이 증가하여 만개 후 165일에서 180일 사이에 내재 휴면이 시작된 것으로 생각되었으며, 만개 후 240일까지 발아 소요 일수가 길고, 발아율이 20% 이하로 낮게 유지되었다. 만개 후 255일에는 내재휴면이 시작되기 전인 165일과 유사한 발아 소요 일수 및 발아율을 보였으며, 이후에도 20일 이내에 발아가 시작되었고, 60% 이상의 높은 발아율을 보여 만개 후 255일에는 내재휴면이 타파된 것으로 생각되었다. 따라서, 중부 지역 ‘Fuji’ 사과의 내재휴면 기간은 만개 후 165일부터 255일 사이로, 10월 말부터 이듬해 1월 초까지 약 70일 정도만 내재휴면 상태를 유지하는 것임을 확인하였다. 또한 만개 후 240일에는 발아가 전혀 진행되지 않은 것으로 볼 때, 내재휴면이 시작된 후 휴면심도가 서서히 깊어져 내재휴면기 후반부인 12월 말경에 이르러 최대에 도달한 후, 급격하게 휴면타파가 진행되는 것으로 판단되었다.
낙엽과수의 내재휴면은 가지의 생장 및 정아의 형성이 완료된 후 시작되는데, 수분과 양분의 공급 상태, 온도, 일장 등의 환경 조건에 따라 차이를 보이고, 동일한 나무 및 가지 내에서도 위치에 따라 다르게 나타나 휴면 개시기를 정확하게 판정하기 어려운 것으로 보고된 바 있다(Kim et al., 2006; Kweon et al., 2013). 또한, 내재휴면 기간을 정의한 연구들에서도 발아 소요 일수 15일을 기준으로 판단하고 있으나(Hauagge and Cummins, 1991; Kim et al., 2006), 이러한 연구들은 대부분 온실 내에서 실험을 진행하였으며, 휴면 개시와 같이 타파 시기는 동일 품종 내에서도 눈의 종류와 위치, 나무의 수령, 재배 지역 등에 따라서도 다르게 나타날 수 있다고 보고되었다(Faust et al., 1995; Hauagge and Cummins, 1991; Kweon et al., 2013). 본 실험에서는 이러한 애로 사항들을 개선하기 위해 발아 소요 일수와 함께 발아율을 조사하여 최초 발아까지 20일 이내, 발아율 60% 이상을 기준으로 외재휴면에서 내재휴면으로, 다시 내재휴면에서 환경휴면으로 전환되는 시점을 찾을 수 있었다. 또한 내재휴면이 타파된 것으로 판단된 시점 이후에도 발아 소요 일수가 기존의 기준보다 5일 정도 늦은 20일로 유지된 것으로 보아 실제 포장에서 채취한 가지를 대상으로 수삽한 경우, ‘Fuji’ 사과나무의 발아 소요 일수 20일을 기준으로 내재휴면기를 판단하여도 무방할 것으로 생각되었다.
발아 특성에 따른 저온요구도 설정
내재휴면의 타파에 있어 0-7.2oC 사이의 온도가 저온 축적에 유효하다는 CH 모델(Weinberger, 1950)이 밝혀진 이래로 세분화된 온도 범위의 점수화 및 최적 온도의 수정 등의 과정을 거친 Utah 모델(Richardson, 1974), North Carolina 모델(Shaltout and Unrath, 1983), Dynamic 모델(Fishman et al., 1987) 등이 개발되었다. 그 중 CH 모델과 Utah 모델은 최근까지도 일반적으로 사용되고 있는 저온 산출 방법이라고 할 수 있다. CH 모델의 경우, 해당 범위의 온도가 관찰된 시간을 단순 적산한 방식으로 계산이 용이하다는 장점을 가지고 있으며, Utah 모델은 기준한 범위 내에서 온도 등급에 따른 유효도를 판정하여 적산하는 방식으로 실제 포장에서 나타나는 결과들과 가장 유사한 값을 보이는 모델로 평가되고 있어 두 모델을 이용하여 내재휴면의 타파에 필요한 저온요구도를 산출하였다.
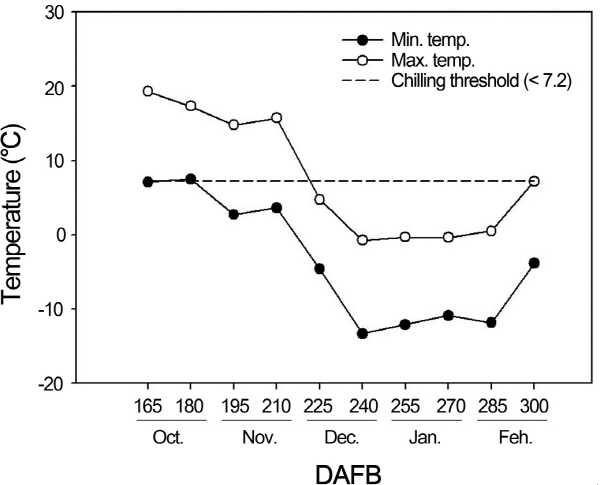
일 최고 및 최저기온을 조사한 결과(Fig. 1), 10월 2일부터 CH 모델의 기준인 0-7.2 범위의 온도가 처음 관찰되었고, 11월 21일에는 최초로 일 최고기온이 7.2 이하로 낮아졌으며, 만개 후 255일 경인 12월 중순 이후부터는 최고온도가 기준 이하로 유지되었다. CH 모델과 Utah 모델 모두 10월 14일 이전에도 각 모델의 저온적산에 유효한 온도가 관찰되었으나, 이 시기는 아직 내재휴면이 시작되기 전으로 내재휴면이 시작된 10월 14일까지 관찰된 유효온도는 내재휴면의 타파에 필요한 저온요구도 계산 시 제외해야 하는 것으로 판단하였다.
|
Fig. 1. Fluctuation of maximum and minimum temperatures during the sampling time in 2011-2012 seasons. |
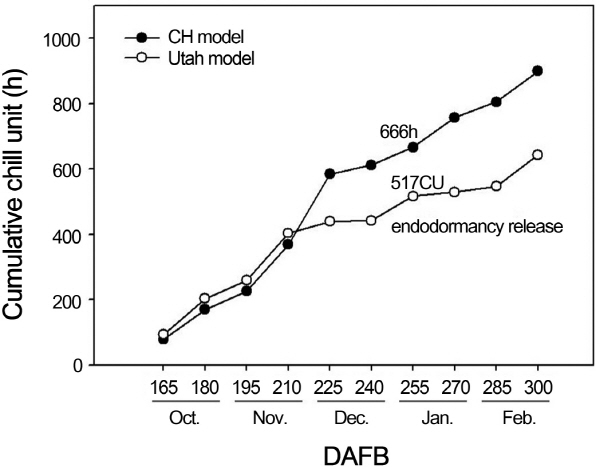
발아 소요 일수 및 발아율 조사를 통해 도출된 내재휴면기의 저온을 산출한 결과, 내재휴면 타파기인 만개 후 255일까지 축적된 저온은 CH 모델의 경우 666h, Utah 모델은 517CU으로 조사되었다(Fig. 2). Utah 모델의 517CU의 저온요구도는 선행 연구들의 결과에 비해 다소 높은 수치로, ‘Fuji’ 사과는 ‘Braburn’, ‘Dorsett Golden’, ‘Gala’, ‘Pink Lady’와 함께 200-400CU의 낮은 저온요구도를 가지는 품종으로 보고되었으며(Andersen and Crocker, 2009; Herter et al., 1988), Hauagge와 Cummins(1991)가 90여개의 사과 품종을 대상으로 저온요구도를 산출한 연구에서 동일한 품종 및 포장에서도 해에 따라 차이를 보인다고 보고하였다. 게다가 본 연구를 수행한 2011년은 2000년대 들어 평균 기온이 가장 높은 해로 조사되어(KMA, 2012), 내재휴면 타파에 소요되는 기간이 길어졌을 가능성이 있는 것으로 예상되었다. 또한, 본 연구에서는 15일 간격으로 시료를 채취하여 내재휴면기로 판단되는 최장 기간을 산출하였는데, 시료 채취 시기인 만개 후 255일과 270일 사이에 휴면이 타파되었다고 가정하였을 때에는 400CU에 보다 근접한 저온요구도를 보일 것으로 생각되었다. 따라서, 본 연구를 수행한 2011-2012년 사이의 기온은 CH 모델의 666h, Utah 모델의 517CU의 저온요구도를 충족시키는 데 충분하였으나, 선행 연구 결과의 400CU와 비교하였을 때, 저온요구도가 충족되는 기간을 41일 정도 지연시켰다. 뿐만 아니라 앞에서 기술한 바와 같이 2011년이 지난 10년간(2001-2010년) 평균기온이 가장 높았고, 향후에도 지구온난화로 인해 계속적으로 겨울철 평균기온이 상승할 경우, 재배적지의 이동이 불가피 할 것으로 생각되었으며, 예상되는 기후변화 시나리오에 적합한 남방한계선의 설정이 요구되었다.
이미 국내에서 CH 모델을 이용하여 사과나무의 저온요구도를 계산하고, 지구온난화에 따른 재배 가능성을 예측한 보고들이 있으나(Kweon et al., 2013; Yun et al., 2001), 생리적으로 정확한 내재휴면 개시기를 파악하지 못했으며, 인위적인 저온 처리를 통하여 얻은 결과들로 본 연구처럼 실제 포장에서 얻은 값과는 많은 차이를 보였다. 또한, 국외의 많은 연구들에서 CH 모델의 저온 기준인 7.2°C보다 높은 온도 조건에서도 휴면타파에 유효한 저온 축적 효과가 있다고 보고되었으며(Dokoozlian, 1999), ‘Budagovski 9’, ‘Malling 9’, ‘Alnarp 2’ 등의 사과 대목 품종의 경우, 12oC까지도 휴면타파에 유효한 온도로 인식하는 것으로 보고되었다(Heide and Prestrud, 2005). 뿐만 아니라 Luedeling et al.(2009)은 캘리포니아 내 동일지역에서 나타나는 온도 변화를 4개의 일반적인 chill 모델에 적용해 본 결과, 1,950과 2,050 사이 범위에 속하는 CH가 측정되었다고 보고하였는데, 이는 기후변화에 대한 모델의 반응 감도가 상당히 다르게 나타날 수 있음을 시사하는 결과로 볼 수 있다. 본 연구의 결과 역시, 두 모델에서 서로 다른 저온요구도가 산출되어 국내 환경에서의 저온요구도 설정에 더 적합한 모델의 선발이 이루어져야 할 것으로 생각되었다.
내재휴면이 타파된 이후 식물체는 환경휴면에 들어가는데, 내재휴면과 환경휴면을 육안으로 혹은 외부적으로 구별하는 것은 거의 불가능하다고 할 수 있다. 그러나 식물체 내부적으로 내재휴면기에는 이를 타파하기 위한 저온을 감지하는 반면, 환경휴면기에는 환경휴면을 종료하고 발아 및 생장을 준비하여 고온을 감지한다(Asakura, 2011; Cesaraccio et al., 2004). 이와 같이 저온요구도와 반대되는 개념인 growing degree hour(GDH)는 기온이 4.5°C 이상으로 상승하면 적산되기 시작하며, 개화기 및 과실의 생장을 예측하는 데 사용될 수 있다. 사과나 포도의 경우, 복숭아, 배에 비해 상대적으로 높은 GDH를 갖고 있기 때문에 내재 휴면이 일찍 타파되어도 개화가 늦은 경향을 보이는 것이라 할 수 있다(Ben Mimoun and DeJong, 1999; Cain, 2006; Hauagge and Cummins, 1991; Lopez et al., 2007). 결과적으로 저온요구도와 GDH의 결합은 생물계절학적 측면에서 각 과종 및 품종의 내재휴면과 환경휴면의 기간을 보다 정확하게 예측할 수 있을 것으로 예상되었다. 반면, 내재휴면이 타파된 이후에 진행되는 과정들을 예측하고자 하는 모델이나 방법들은 내재휴면기, 특히 내재휴면 타파기를 기준으로 하고 있기 때문에 이를 정확하게 파악하는 것이 우선되어야 할 것이다.
휴면 심도에 따른 눈 발달 관찰
사과나무의 꽃눈 형성에는 개시와 발달의 두 가지 과정이 관여하고 있다(Jackson and Sweet, 1972). 세포 수준에서 볼 때, 눈의 분열 조직 내에서 발생하는 개시 과정은 비교적 짧은 1-2주의 시간만을 필요로 하며, 바로 뒤이어 이루어지는 발달 과정은 눈에 존재하는 화기 구조 생성을 위해 오랜 시간을 필요로 한다. 이 중 발달 과정은 가을과 겨울 즉, 휴면 기간 동안 느리게 진행되며 이듬해의 성공적인 개화와 높은 생산량 확보를 위한 매우 중요한 과정이다. 또한 눈의 내재휴면 유지 및 타파는 눈 자체의 내부 구조와도 밀접한 관계를 갖고 있기 때문에(Ramina et al., 1995) 휴면기와 휴면타파기 동안의 ‘Fuji’ 사과나무 눈의 발달 양상을 관찰하였다.
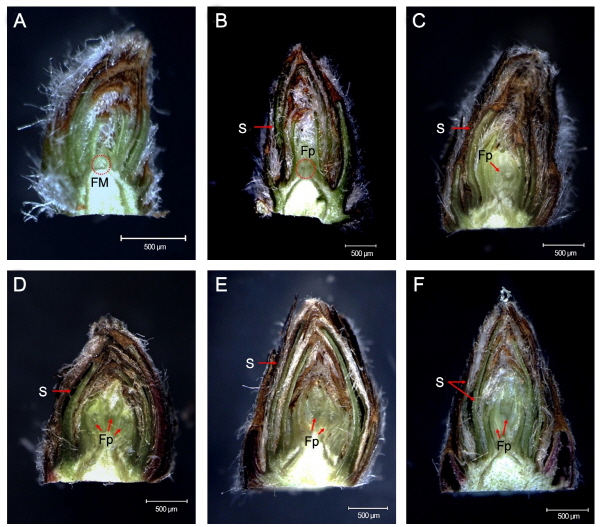
7월 중순경부터 분열조직이 비대하여 돌기모양으로 솟아오른 형태가 관찰되어 이 시기에 영양생장으로부터 생식생장으로의 전환이 이루어지는 것을 알 수 있었으며(Fig. 3A), 9월에는 중심화의 화원기가 뚜렷하게 관찰되었다(Fig. 3B). 일반적으로 사과나무는 하나의 눈 속에 4-6개의 꽃이 형성되며, 꽃눈 분화를 위한 자극이 전달되면 약 10일 정도 후에 중심화의 화원기가 만들어지고, 15-20일 후에는 제 1측화가 분화되며 제 4, 5측화는 40-50일이 지나야 분화된다고 알려져 있다(Bergh, 1985; Foster et al., 2003). 본 연구에서도 기존의 연구 결과(Bergh, 1985)와 동일하게 중심화의 원기가 관찰된 후 약 90일 정도 지난 12월에 최소 3개 이상의 화원기가 형성되어 있었으며(Fig. 3D), 중심화의 화원기에서는 뚜렷하지 않으나 수술이 관찰되어 화기의 대부분이 완성되어 있었다. 또한 12월은 수삽 실험 시 전혀 발아하지 않았기 때문(Table 1)에 내재휴면 심도가 최대에 도달한 시기임이 밝혀져 내부의 구조적인 변화가 거의 없을 것으로 생각되었으나, 내부적인 발달은 지속되고 있음을 알 수 있었다(Fig. 3F). 따라서 ‘Fuji’ 사과의 내재휴면은 꽃의 원시세포(flower primordium)가 형성되는 시기에 시작되어 수술을 포함한 꽃의 내부 구조물이 형성된 이후 타파됨을 확인하였으며, 이를 통해 휴면과 눈의 내부 구조와의 유기적 관계를 밝힐 수 있었다. 또한 화원기가 형성된 후, 눈에 여러 겹의 인편 사이로 털이 촘촘하게 발달하였으며, 가장 바깥쪽의 인편은 시기가 진전될수록 더 두껍고 단단해져 추위를 견디기 위해 발달하는 것임을 확인하였다.
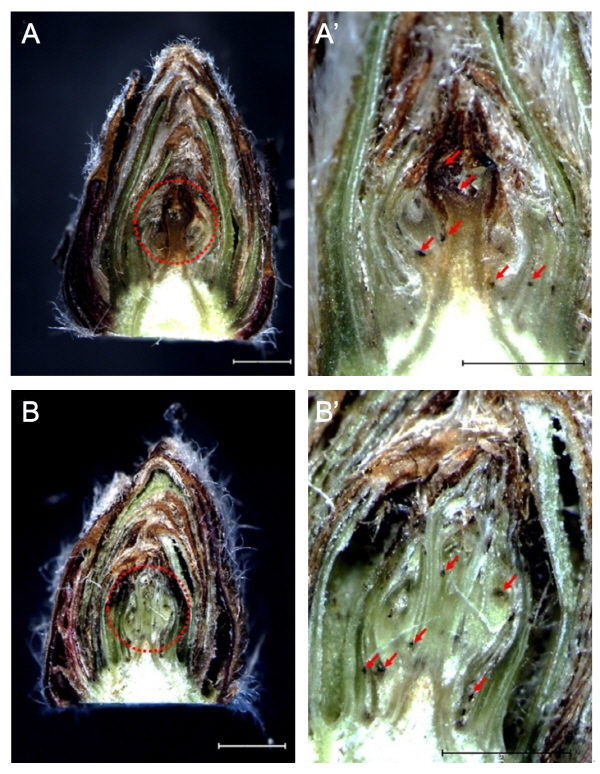
휴면 기간 동안의 눈 발달을 관찰하는 과정에서 대부분의 눈이 건전하게 생존해 있음을 확인하였지만 일부에서 화원기의 괴사 현상이 나타났다(Fig. 4). 꽃눈 괴사 시 냉각 또는 서리에 의한 피해는 저온에 직접적인 영향을 받기보다는 주로 얼음결정 형성에 의해 발생하며, 얼음 결정이 발생하는 위치에 따라 세포의 피해 유무가 달라진다고 알려져 있다. 대부분의 식물 조직에서 얼음결정의 생성은 물관 또는 인편과 인편 사이, 세포 간극 등의 세포외공간(extracellular space)에서 시작되고(Guy, 1990), 생성된 얼음결정은 세포 내부의 수분을 세포 밖으로 이동시켜 삼투 농도를 급속하게 상승시키는 원인이 된다(Burke et al., 1976). 이러한 세포외동결의 효과는 세포의 점진적인 탈수를 유도해 세포내동결을 방지하는 것이나, 극심한 탈수로 인해 원형질의 응고(coagulation)가 발생할 수 있으며, 얼음 결정에 의해 세포막과 다른 세포 구성 요소 사이를 붕괴시켜 세포 괴사를 가속화시킬 수 있다(Burke et al., 1976; Rodrigo, 1999; Steponkus, 1984). 세포의 붕괴는 일반적으로 영향을 받은 조직의 변색으로 구분할 수 있으며, Fig. 4에서도 동일하게 변색 여부로 세포의 붕괴, 즉 괴사 여부를 판단할 수 있었다. 특히, 주목할만한 사실은 건전하게 살아 있는 세포들 사이로 부분부분 검은 점의 형태가 발견된 것으로(Fig. 4A’, B’), 이 검은 점을 중심으로 주변을 향해 퍼져나가는 수침 및 갈변 현상이 관찰되었으며, 이미 괴사가 완전히 진행된 화원기 안에서도 동일하게 검은 점이 발견되어(Fig. 4, arrow) 선행 연구에서 언급된 얼음결정의 형성으로 인해 일부 조직의 세포가 먼저 피해를 입고, 여기로부터 괴사가 진행되고 있음을 확인할 수 있었다.
|
Fig. 4. Cross section of floral bud necrosis in ‘Fuji’ apple trees at 255 (A, A’) and 300 (B, B’) days after full bloom. Arrows indicate necrotic spots. Scale bars = 500 μm. |
꽃눈과 잎눈의 외적 특성 비교
눈의 외적 특성을 잎눈과 꽃눈으로 구분하여 비교한 결과, 잎눈은 관찰 시기에 따른 큰 차이가 없는 것으로 조사되었으나 꽃눈은 9월 이후 꽃눈 분화의 영향으로 눈의 크기가 다소 증가하였다(Table 2).
시기별로 관찰한 결과를 토대로 눈의 외적 특성만을 이용해 잎눈과 꽃눈을 명확하게 구분하는 방법을 찾고자 잎눈과 꽃눈을 비교 분석하였을 때, 잎눈과 꽃눈은 가지의 직경과 눈의 종경, 횡경 모두에서 차이가 있었다(Table 3). 이듬해 생산량과 직결되는 꽃눈은 잎눈에 비해 평균적으로 종경이 1.30 ± 0.06mm, 횡경이 0.97 ± 0.07mm 더 컸으나, 꽃눈이 위치한 가지의 직경은 잎눈보다 1.02 ± 0.14mm 정도 더 가는 것으로 조사되어 많은 꽃눈을 분화시키기 위해서는 가지의 직경을 7.0mm 이하로 생장시키는 것이 긍정적인 효과를 보일 것으로 생각된다. 이는 ‘Campbell Early’와 ‘Hongisul’ 포도에서 각각 8.0과 8.5mm를 기준으로 가지 굵기에 따른 꽃눈의 괴사율의 차이가 나타나 가지 굵기와 눈괴사 현상에 밀접한 연관이 있다는 기존의 보고(Choi et al., 2007; Kim et al., 2011)와도 일맥상통하는 결과이다. 이러한 경향은 가지의 수세가 강할수록 생육이 활발해 눈으로 전달되는 양분이 부족해지기 때문에(Collins and Rawnsley, 2005; Scagel et al., 2010) ‘Fuji’ 사과의 꽃눈 발달이 저해되는 것으로 생각되며, 재배 시 수세가 너무 강해지는 것은 피해야 할 것으로 판단된다.
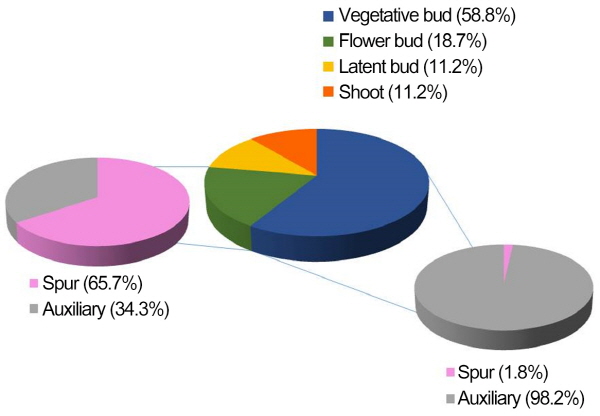
또한 꽃눈은 과대지의 액아와 단과지의 정아(Fig. 5)에서 각각 34.3과 65.7%의 비율로 나타나 단과지의 정아에서 더 많이 형성되었다. 그러나 모든 단과지의 정아에서 꽃눈 분화가 이루어지지 않았기 때문에 다시 단과지 내에서 잎눈과 꽃눈을 구분하여 비교한 결과, 잎눈과 꽃눈 차이는 오로지 크기에 의한 것으로 꽃눈은 잎눈보다 평균 종경이 1.4 ± 0.32mm, 횡경은 0.78 ± 0.12mm 더 컸다(Table 5). 따라서 단과지의 발달을 유도시키는 것이 꽃눈 확보에 보다 유리할 것으로 판단된다.
