

서 언
재료 및 방법
식물재료
재배포장에서 착과율을 이용한 교배친화성검토
화분관 신장 검경을 통한 교배친화성 검토
자가불화합 유전자 밴드패턴 비교
동계전정 가지를 이용한 인위적 착과조건에서의 교배친화성 검토
결과 및 고찰
재배포장조건에서 P. pyrifolia와 다른 종간의 교배친화성
기내 화분관신장에 따른 P. pyrifolia와 다른 종간의 교배친화성 검토
S allele를 확인을 통한 P. pyrifolia와 다른 종간의 교배친화성 검토
항온조건에서 P. pyrifolia와 다른 종간의 교배친화성 검토
서 언
Pyrus 속은 24개의 기본종과 자연적으로 발생된 6개의 종간잡종이 있는 것으로 보고되고 있다(Hancock and Lobos, 2008). 배의 원산지는 P. pyrifolia와 P. ussuriensis가 유래된 중국, P. communis, 그리고 P. communis와 P. bretschneideri 간의 종간잡종이 유래된 중앙아시아와 서아시아, 그리고 P. communis의 자생종이 유래된 코카서스 산맥과 소아시아의 세 개 지역으로 추정되고 있다(Vavilov, 1951).
동아시아 지역을 중심으로 분화된 동양배는 육질이 아삭하고 감미가 높은 과실의 특성을 가지고 있고 유럽에서 주로 재배되는 서양배는 동양배에서 많이 나타나는 흑성병에 대한 내성을 가지고 있거나 동양배에서는 볼 수 없는 붉은 과피색을 나타내는 품종이 있어 새로운 형질을 도입하거나 유전연구를 위한 집단작성은 유전적으로 거리가 먼 종간교배 방법이 대안으로 인식되고 있다. 이러한 이유로 세계 각국은 새로운 형질을 보유한 품종 개발을 위해 인위적으로 종간교배 방법을 이용하여 품종육성을 시도하고 있다(Bell et al., 1996; Kwon et al., 2017).
뉴질랜드에서는 서양배의 풍미와 표주박 형태를 가지며 동양배 타입의 아삭하고 부드러운 육질을 갖는 품종 또는 보구력이 길고, 유년성이 짧으며 병해에 강한 유용형질을 도입하기 위해 서양배와 동양배의 종간교배를 시도하고 있다. 그 결과, ‘Max Red Bartlett’과 ‘Huobali’로부터 적색유전자를 도입하여 품종을 육성하여 품종보호출원 중이며, 상업적 가능성이 높은 우량 계통들을 선발하여 그 특성을 정밀검토 중에 있다(White and Brewer, 2002).
미국에서는 P. calleryana와 P. kawakami Hayata 등을 경관 조성용 품종 육성소재로 활용해 왔다. P. calleryana에 속하는 ‘Bradford’나 P. kawakami는 기후 및 토양에 적응성이 좋아 관상용으로 적합한데 fire blight에 감수성을 보여 재식에 제약요인이 되고 있다. 이 때문에 종의 다양성에 기초한 종간교배 방법에 의해 기존 품종의 단점을 보완할 수 있는 병 저항성 경관조성용 품종을 개발 중에 있다(Pittenger, 2010).
이탈리아에서는 1978년 과실 품질을 향상시키고, 적색 과피, psylla, fire blight 저항성 등을 목표로 서양배와 동양배의 종간잡종을 이용하였고, 1991년 이후로는 유전적 다양성을 높이고 시장의 다양한 요구를 반영하는 새로운 형태의 과실을 육종하기 위해 종간 잡종을 이용하고 있다(Sasavini et al., 2002).
일본에서는 지금까지의 교배육종이 소수의 품종이나 계통을 중복 사용함에 따라 유전적 다양성이 저하되는 것이 문제점으로 지적되면서 이러한 근친교배를 막기 위해 P. bretschneiderie와 종간교배를 함으로써 그 후대에서 ‘Oushu’가 육성되었다(Kotobuki et al., 2004).
국내에서도 병 저항성이 뛰어난 것으로 알려진 야생종 배의 유용형질 도입을 위해 주요 형질을 지배하는 유전자를 찾거나 분자 표지에 근거한 유전자 연관 지도가 매우 효율적으로 이용될 수 있기 때문에 한국 야생배 P. ussuriensis와 P. pyrifolia의 종간잡종 96개체를 대상으로 RAPD, SSR, AFLP 마커를 이용하여 genetic linkage map을 작성해 보았으나 개발된 유전자 지도의 연관군 수가 배의 염색체 17개와 일치하지는 않았다(Choi et al., 2010).
형태적, 유전적으로 차이가 큰 두 개 종을 교배친으로 이용하면, 새로운 형질을 보유한 품종선발 및 유전 연구소재 확보가 용이할 것으로 생각된다. 추후 상업적으로 시장성이 있으면서 다양성이 높은 차별화된 후대를 얻기 위해 P. pyrifolia와 P. communis의 종간교배가 요구되고 있다.
본 연구는 종간의 유효형질을 보유한 품종을 육성하기 위하여 시도되고 있는 종간교잡과정에서 포장조건과 항온조건에서의 착과량, 생체 내에서의 화분관 신장, 자가불화합 유전자 밴드패턴을 확인하여 종간교잡의 실효성을 평가하기 위하여 수행하였다.
재료 및 방법
식물재료
P. pyrifolia ‘Niitaka’와 주요 종에 속하는 품종들과의 교배친화성 검토를 위해 2013-2015년 나주시 금천면에 소재한 농촌진흥청 배연구소 포장에 재식되어 있는 P. betulaefolia에 속하는 ‘OPR114’, ‘OPR260’ 2품종, P. bretschneideri에 해당하는 ‘Dangshansuli’, ‘Xuehuali’, ‘Yali’ 3품종, P. calleryana인 ‘OPR125’, ‘OPR195’, ‘OPR249’ 3품종, P. communis에 속하는 ‘Bartlett’, ‘Bosc’, ‘Canal pear’, ‘Max Red Bartlett’ 4품종, 그리고 같은 P. pyrifolia에 속하는 ‘Chuhwangbae’ 1품종을 부본으로 P. pyrifolia ‘Niitaka’를 모본으로 이용하였다.
2013년 4월 포장에서 교배친화성을 검정하고 2014년 3월에 실내조건에서 교배친화성을 검정하였으며 2015년 PCR방법을 이용하여 자가불화합인자를 확인하였다.
재배포장에서 착과율을 이용한 교배친화성검토
재배포장에서 ‘Niitaka’를 종자친으로 하여 5종 13품종의 꽃가루를 인공수분하였다. 교배를 위한 제웅은 화뢰기에 꽃받침과 꽃잎, 수술을 동시에 제거하는 화판 인발법을 이용하여 실시하고, 제웅 다음 날 준비된 꽃가루를 이용하여 교배하였다(Kang et al., 2008). 착과율 조사는 교배 1달 후 유과가 충분히 비대되어 육안으로 착과가 확인되었을 때 수행되었다.
화분관 신장 검경을 통한 교배친화성 검토
화분관 신장 검경을 위해 교배 후 72-96시간에 꽃을 채취하여 FAA(formalin: acetic acid: ethyl alcohol(80%) = 5 : 5 : 90 v/v/v) 용액에 고정하였다. 연화는 1N NaOH 용액이 들어있는 용기에 시료를 넣고 60°C의 수조에서 40분간 실시하였으며, 그 후 상온에서 aniline blue 염색액에 24-48시간 염색하고 squash법으로 슬라이드를 제작하였다(Kho and Baer, 1968). 화분관 신장은 invert type 형광현미경(Diaphot 300, Nikon, Japan)을 이용하여 관찰하였다.
자가불화합 유전자 밴드패턴 비교
S-allele을 확인하기 위해 PCR을 실시하여 양친의 자가불화합 유전자 밴드패턴을 비교하였다. 어린잎에서 DNeasy Plant Mini Kit(Qiagen, Germany)를 이용하여 genomic DNA를 추출하고, 배 S-allele 특이적 primer(FTQQYQ, anti-(I/T)IWPNV)를 이용 PCR을 수행하였다. PCR 조건은 94°C 5분간 반응시킨 다음 94°C에서 1분, 54°C에서 2분, 72°C에서 3분 30cycle을 반복한 후 72°C에서 7분간 반응하였다. 그 후, 자동전기영동분석장치(Multina, Shimatsu, Japan)를 사용하여 PCR 산물을 확인하였다.
동계전정 가지를 이용한 인위적 착과조건에서의 교배친화성 검토
재배포장조건에서 나타날 수 있는 환경에 의한 교배불친화성 요인을 배제하고자 휴면이 타파된 동계전정 가지를 3월에 채취하고 18°C의 실내조건에서 수삽하여 인위적으로 개화시킨 후 재배포장조건에서 수행했던 동일한 조합으로 교배를 실시하여 종간의 교배친화성을 검토하였다. 착과율 조사는 교배 2주 후 씨방이 부풀어 오른 정도와 씨방 내 종자의 발달 정도를 기준으로 판단하였다. 착과율은 교배화수에 대한 착과수를 조사하여 비율로 산정하였다.
결과 및 고찰
재배포장조건에서 P. pyrifolia와 다른 종간의 교배친화성
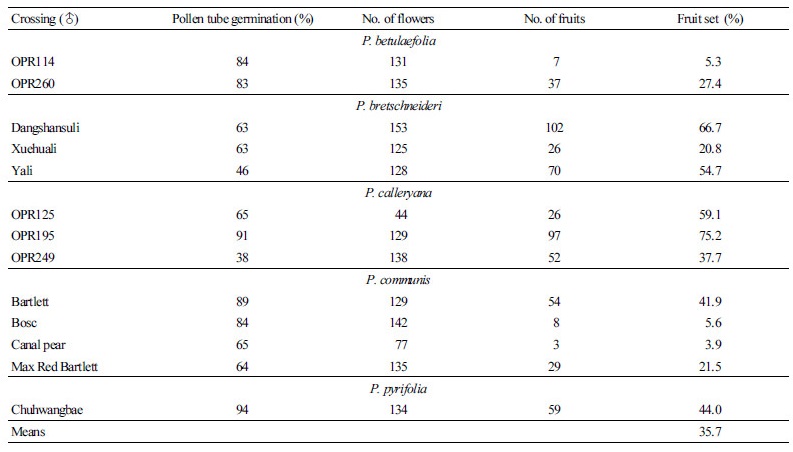
재배포장조건에서 P. pyrifolia ‘Niitaka’와 주요 종에 속하는 품종들과의 교배친화성을 착과율을 기준으로 검토한 결과 품종간 차이는 있었지만 부본으로 P. bretschneideri와 P. calleryana를 사용했을 때 P. betulaefolia와 P. communis를 사용한 경우보다 P. pyrifolia ‘Niitaka’의 착과율이 높은 경향을 나타내었다. 착과율은 P. calleryana ‘OPR195를 교배한 경우 가장 높았고 P. communis ‘Canal pear’를 교배한 경우 가장 낮았다(Table 1).
결과적으로 포장조건에서 P. pyrifolia ‘Niitaka’는 P. betulaefolia 에서는 ‘OPR114’, P. communis에서는 ‘Bosc’과 ‘Canal pear’과 교배불친화성을 나타냈다. 이는 배는 주요한 자가불화합 장벽이 없어 모든 종은 교잡이 가능하다는 결과(Folta and Gardener, 2009)와 상이하여 추가적인 확인이 필요하였다.
기내 화분관신장에 따른 P. pyrifolia와 다른 종간의 교배친화성 검토
재배포장 조건에서 착과율이 저조한 교배조합의 원인 탐구를 위해 포장에서 교배한 주두 내 화분관신장 정도를 현미경을 이용하여 검경하였다.
착과율이 높았던 ‘Dangshansuli’ 화분을 교배한 ‘Niitaka’의 주두에서 화분관 신장은 활발하게 이루어지는 것을 확인할 수 있었다(Fig. 1A). 또한, 착과율이 높았던 ‘OPR125’ 화분을 교배한 ‘Niitaka’의 주두에서 화분관 선단이 비대하는 현상이 일부 관찰되기도 하였다(Fig. 1B).
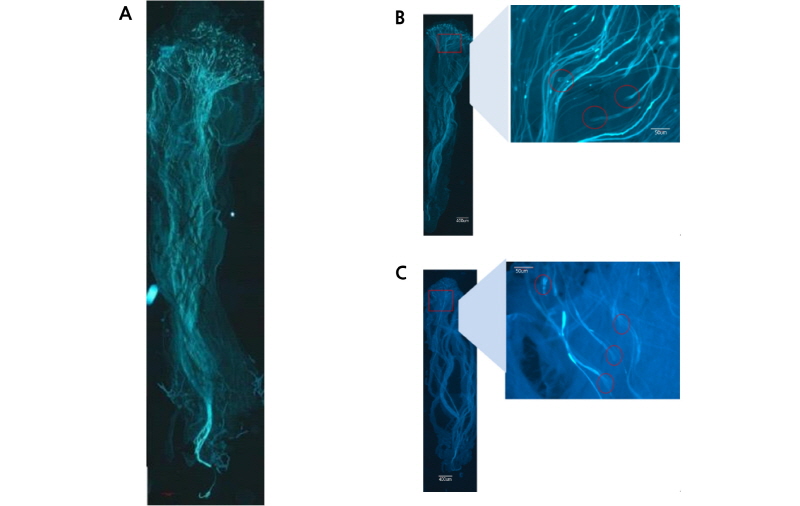
Fig. 1. Micrographs of pollen tube elongation in squashed styles of ‘Niitaka’ pear after artificial pollination in field conditions. A, ‘Niitaka’ × ‘Dangshansuli’; B, ‘Niitaka’ × ‘Xuehuali’; C, ‘Niitaka’ × ‘OPR125’. Scale bar = 200 µm. Circle shows a hypertrophied pollen tube tip in the amplified region (demarcated by a rectangle).
그런데 재배포장에서 착과율이 낮았던 ‘Xuehuali’ 화분을 교배한 ‘Niitaka’의 주두에서 화분관 선단이 비대하는 현상이 관찰되기도 하였지만 자방까지 도달한 화분관도 관찰되었다(Fig. 1C).
결과적으로, 주두로 침입한 화분관의 선단이 비대하여 신장을 멈추는 자가불화합성 작물에서 나타나는 전형적인 현상(Hiratsuka et al., 2001)이 있었지만 자방까지 도달한 화분관도 관찰되어 포장조건에서 나타난 교배불친화성에 대한 유전적 환경적 검증이 필요하다고 판단되었다.
Table 1. Effects of pollen parent cultivar on fruit set in ‘Niitaka’ pear after artificial pollination in the field conditions
 |
S allele를 확인을 통한 P. pyrifolia와 다른 종간의 교배친화성 검토
포장조건에서 나타난 교배불친화성을 S allele 확인을 통하여 검정하였다.
S allele 확인을 위해 특이 primer를 사용하여 PCR을 한 결과, 특정 제한효소로 절단하지 않아도 차이점이 확인될 정도로 PCR product의 크기는 다양하였다. ‘Niitaka’의 경우 자가불화합 인자는 S3S9로 PCR product는 380bp, 1,300bp의 크기로 확인되었다. ‘Xuehuali’, ‘OPR114’, ‘OPR260’, 그리고 ‘Chuhwangbae’에서는 ‘Niitaka’의 380bp와 유사한 크기인 360bp의 PCR product가 얻어져 부분불친화성을 확인할 수 있었지만 ‘Dangsanshasuli’, ‘Yali’, ‘Bartlett’, ‘Bosc’, ‘Canal pear’, ‘Max Red Bartlett’, ‘OPR125’, ‘OPR195’, 그리고 ‘OPR249’ 화분에서는 ‘Niitaka’와 동일한 PCR product가 확인되지 않았다(Fig. 2).

Fig. 2. Self-incompatibility gene pattern of ‘Niitaka’ compared to 13 other cultivars. PCR was carried out with SI-specific primers. The lanes show, from left to right, L, 1 Kb ladder; 1, ‘Niitaka’; 2, ‘Dangsanshasuli’; 3, ‘Xuehuali’; 4, ‘Yali’; 5, ‘Bartlett’; 6, ‘Bosc’; 7, ‘Canal pear’; 8, ‘Max Red Bartlett’; 9, ‘OPR125’; 10, ‘OPR195’; 11, ‘OPR249’; 12, ‘OPR114’; 13, ‘OPR260’; 14, ‘Chuhwangbae’. Each product was loaded by an automatic electrophoresis system. Dotted lines indicate PCR product sizes from ‘Niitaka’ (upper line; 1,400 bp, lower line; 380 bp).
유전적 장벽에 의한 교배불친화성은 자가불화합 인자가 모두 같을 경우에만 해당하므로 식물체가 갖는 자가불화합 인자 두 개 중 하나가 다르면 교배친화성에는 문제가 없는 것으로 알려져 있다(Hiratsuka and Zhang, 2002). 따라서, 배에서 종간교배 품종 작성 시 자가불화합 인자에 의한 교배불친화성은 문제가 없는 것으로 판단되었다.
항온조건에서 P. pyrifolia와 다른 종간의 교배친화성 검토
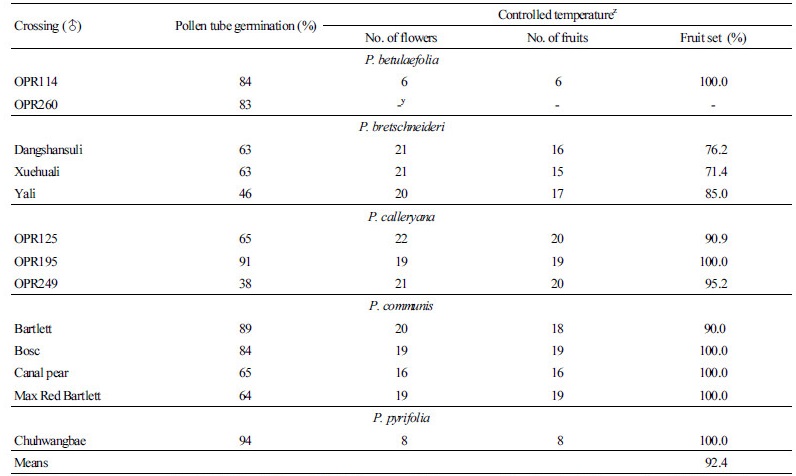
포장조건에서 나타난 교배불친화성을 극복하기 위하여 18°C의 실내조건에서 수삽된 가지를 이용하여 인공수분으로 교배친화성을 검토하였다.
‘Niitaka’에 교배된 P. bretschneideri ‘Dangshansuli’, ‘Xuehuali’, 그리고 ‘Yali’의 착과율은 각각 76.2%, 71.4%, 85.0%를 나타내었지만 P. betulaefolia ‘OPR114’와 P. calleryana ‘OPR125’, ‘OPR195’, ‘OPR249’, P. communis ‘Bartlett’, ‘Bosc’, ‘Canal pear’, 그리고 ‘Max Red Bartlett’ 및 P. pyrifolia ‘Chuhwangbae’를 이용한 교배조합에서는 90% 이상의 착과율을 나타내 재배포장에서 확인되었던 교배불친화성을 확인할 수 없었다(Table 2). 이러한 결과는 P. communis를 교배모친으로 활용하였을 때 종간교잡에는 자가불화합 장벽이 없었다는 결과(Folta and Gardener, 2009)와 일치하였다.
결과적으로 재배포장 조건에서 P. pyrifolia ‘Niitaka’와 주요 종에 속하는 품종들과의 교배불친화성이 확인되었지만 기내 화분관 신장을 관찰한 결과 신장되는 화분관의 선단이 비대되는 현상이 일부 관찰되기도 하였지만 자방까지 도달한 화분관이 도달되는 것을 관찰할 수 있었고, S allele 확인을 위해 특이 primer를 사용하여 PCR을 한 결과에서도 교배불친화성을 설명할 수 있는 결과를 확인할 수 없었다. 그런데, 18°C의 실내조건에서 수삽하여 개화시킨 후 재배포장에서 수행했던 동일한 조합으로 교배를 실시한 결과 모든 종에서 교배친화성이 확인되었다.
따라서, 종간의 유효형질을 보유한 품종을 육성하기 위하여 시도되고 있는 종간교잡 과정에서 나타나는 자연포장에서의 교배불친화성은 수분과 수정과정에서의 환경조건에 의한 것이라고 판단되었다.
