

서 언
재료 및 방법
시험 온실, 작물, 재배 관리
온실 내부 환경 요소 측정
식물체 광합성 요소와 과실 비대 측정
통계분석
결과 및 고찰
온실 내부 환경 변화
식물체의 광합성 요소
과실 비대
서 언
수박의 품질과 생산성을 증대시키기 위하여 토양수분함량(Huh et al., 2020), 토양시비관리와 연작장해 방지(Ahn et al., 2010a, 2010b) 등에 집중되어 연구되어 왔다. 최근에는 정식 후 생육에 영향을 미치는 묘의 품질에 대한 환경 요인의 영향이 연구되었다(Kwack and An, 2021). 하지만 파프리카, 토마토와 같은 온실 작물에 비하여 재배 기간 중 환경이 작물 생육에 미치는 영향에 대한 연구가 많이 진행되지 않았다. 특히, 현재 수박 재배는 광이나 기온의 제어가 되지 않아 외부 환경 변화에 대한 영향을 직접적으로 받는다. 작물의 생육은 광합성 대사를 기반으로 이루어지고 있어 광 환경은 많은 시설 재배 작물에서 과실 품질과 수확량에 영향을 미치는 것으로 알려져 있다(Heuvelink et al., 2004; Jeong et al., 2009a, 2009b; Marcelis et al., 2004). 온실 내부에서 광 환경은 지역 및 계절, 온실 형태(Choi et al., 2013; Kim, 2001; Jeon et al., 2022), 피복재 (Choi et al., 2006; Yoon et al., 2020) 및 차광 소재(Salazar-Canales et al., 2021), 그리고 온실 방향과 내부에서의 위치(Roberts, 1998; Seo et al., 2021) 등 다양한 조건에 따라 차이를 나타낸다. 특히, 온실의 방향은 일중 해의 고도 변화에 따라 내부 수광량의 차이를 나타내며, 단동 온실에서 더욱 차이가 크다(RDA, 2018). 토마토 재배에서 온실 방향에 따라 기온, 환기횟수, 조도 등과 미기상의 차이로 인하여 남북방향에서 숙기가 빠르고 수확량이 많지만, 참외는 생육 및 수량에 있어 온실 방향의 영향을 받지 않는다고 보고되었다(Choi et al., 2008). 이러한 온실 방향에 따른 광 환경 차이는 광합성 생리에 대한 차이를 가져온다. 식물체에 광이 조사되면 내적 요인인 abscisic acid, auxin, phytochrome B 등이 기공 열림(Shimadzu et al., 2019)에 관여하는데 특히, blue와 red 파장의 강도가 기공의 직경 확장에 크게 영향을 미치는 것으로 알려져 있다(Lurie, 1978). 이러한 기공 열림 정도는 기공전도도(Verma et al., 2020)와 수증기압포차(Lee et al., 2012)의 차이를 유발시키고 증산량과 광합성에 영향을 준다. 파프리카 생육 시 증산 속도와 수증기압포차는 일사량과 높은 정의 상관을 나타내는 것으로 보고되었다(Tai et al., 2010). 잎의 온도는 대기 온도의 영향을 크게 받으며(Kim et al., 2020; Yun et al., 2020; Lee et al., 2021), 잎의 수증기 함량은 내부와 외부의 수분포텐셜로 인해 변화하고 이로 인한 엽온의 변화는 증산 속도에 영향을 미친다(Bange, 1953). 국내 첨단 온실 작물에서는 이러한 내부 환경 요인 변화에 따른 생육 모델들이 세밀히 연구되고 있지만 주로 단동형 온실과 토양에서 재배되고 있는 수박에서는 온실방향에 따른 내부 환경변화, 그리고 이에 따른 작물의 생리적 변화에 대한 연구가 미흡한 실정이다. 따라서 본 연구는 동일한 위치 및 외부환경 조건 하에 씨 없는 수박이 재배되고 있는 동서(동 → 서, EW) 및 남동북서(남동 → 북서, SENW) 방향의 온실 간 내부 환경 차이, 그리고 이에 따른 과실 비대 차이를 알아보기 위해 수행되었다.
재료 및 방법
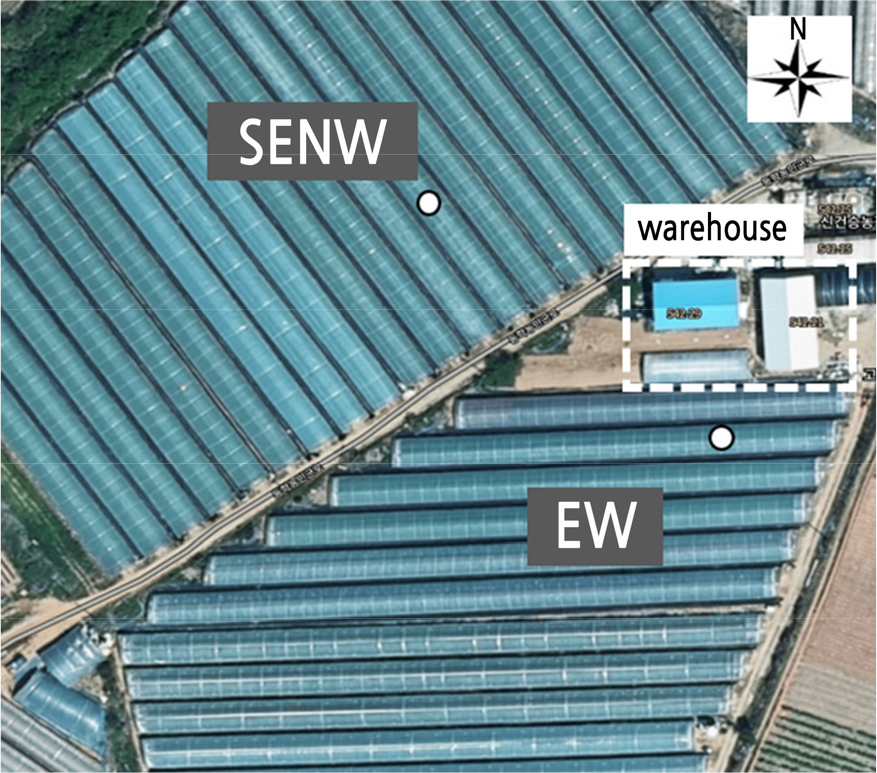
시험 온실, 작물, 재배 관리
본 연구는 2021년 7월 8일부터 8월 5일까지 전북 고창군 내 동일한 위치(방위 N 35°24'42.1", E 126°32'27.2")에서 수박이 재배되고 있는 설치 방향이 다른 온실을 대상으로 수행되었다. 두 온실은 모두 무가온 단동형 온실(폭 × 측고 × 길이 = 6m × 2.5m × 100m)이며 동일한 구조와 피복재를 사용하고 있었다. 두 온실의 설치 방향은 각각 동쪽에서 서쪽(EW), 남동쪽에서 북서쪽(SENW) 이었다(Fig. 1). 시험 품종은 씨 없는 수박 ‘쓰리엔플러스(THREEN PLUS, Pan Pacific Seed PPS Co., Korea)’로 6월 5일에 정식이 되었고 본엽 7장이 발생할 때 5장을 남기고 적심이 되었다. 적심 후 발생한 측지(아들줄기) 중 3줄기를 남기고 나머지를 제거하였다. 이후 각 줄기의 15–20마디에서 발생한 암꽃에 수꽃을 이용하여 인공수정을 실시하였다. 수정 완료 후 온실 방향별로 10주를 선발하여 3줄기 중 직경 10cm 정도로 비대한 과실 1개를 남기고 나머지를 적과하였다. 2021년 7월 28일 이후에는 지나친 내부 온도의 상승을 막기 위해서 온실 외부에 35% 차광막을 온실의 피복재에서 40cm 위에 설치하였다.
온실 내부 환경 요소 측정
2021년 7월 8일부터 8월 4일까지 온실 내부의 환경 요소인 광량, 기온, 상대습도는 각 요소의 센서가 장착된 환경계측기(aM-31, Wisesensing, Korea)를 30분 간격으로 자동 저장하도록 설정하여 온실 중앙의 1m 높이에 설치하였고 광도는 PAR 범위 파장의 광량자속밀도(photosynthetic photon flux density, PPFD)로 수집되었다. 수집된 환경 데이터들은 광량이 0인 시간을 야간, 이외 시간을 주간으로 분리하여 평균하였다. 광도와 기온의 일중 변화는 수집 데이터에서 매일 동일한 시각의 데이터를 평균하였다. 일 적산 일사량(daily integrated solar radiation, mol·m-2·day-1)은 일별로 30분 간격으로 측정된 주간의 PPFD 값에 1,800초(30분)를 곱한 후 합하였다. 식물체 주변의 환경 요소는 2021년 7월 8일부터 8월 4일까지 일주일 간격으로 오전 11시경에 온실별 10주 중 3주를 등간격으로 선발하여 측정되었다. 각 식물체별로 과실 착과 부위에 위치한 잎의 근접 PAR의 총 광도(PPFD) 및 파장별 광도는 광 스펙트럼 분석기(LI-180, LI-COR, USA)를 이용하여 측정하였다. 잎의 온도(엽온)는 착과 부위 잎들 중에서 그늘이 발생하지 않은 상부 잎을 형광 측정기(LI-600, LI-COR, USA)를 이용하여 측정하였다. 엽온과 기온의 차이(엽온 - 기온)는 형광 측정기에서 측정된 엽온과 환경계측기에서 측정된 동일 시각의 기온을 이용하였다.
식물체 광합성 요소와 과실 비대 측정
위의 식물체 착과 부위 잎들 중 그늘이 발생하지 않은 상부 잎을 대상으로 오전 11시경 형광 측정기(LI-600, LI-COR, USA)를 이용하여 기공전도도(stomatal conductance), 수증기압포차(vapor pressure deficit), 광계Ⅱ의 광량자이용효율(photon use efficiency, ØPSⅡ), 증산속도(transpiration rate)를 측정하였고 각각 식 (1), (2), (3), (4)에 의해 계산되었다.
기공전도도(stomatal conductance)
gsw : 기공전도도(mol·m-2·s-1)
gbw : 경계층 전도도(mol·m-2·s-1)
gtw : 총 전도도(mol·m-2·s-1)
수증기압포차(vapor pressure deficit)
VPDleaf : 수증기압포차(kPa)
VPleaf : 잎의 수증기압(kPa)
VPcham : 챔버(온실)의 수증기압(kPa)
광량자이용효율(quantum use efficiency)
Ø PSII : 광계(photosystem)Ⅱ의 광화학에너지 변환 유효광량자수율
Fm’ : 빛에 적응된 상태에서의 최대 형광강도
Fs : 빛에 적응된 상태에서의 최소 형광강도
증산속도(transpiration rate)
E : 증산 속도(µmol·m-2·s-1)
ui : 잎으로의 수증기 흐름 속도(µmol·m-2·s-1)
Wref and Wsam : 잎으로 들어오고 나가는 수증기의 몰 분율(mol·H2O·mol·air-1)
s : 잎의 면적(m2)
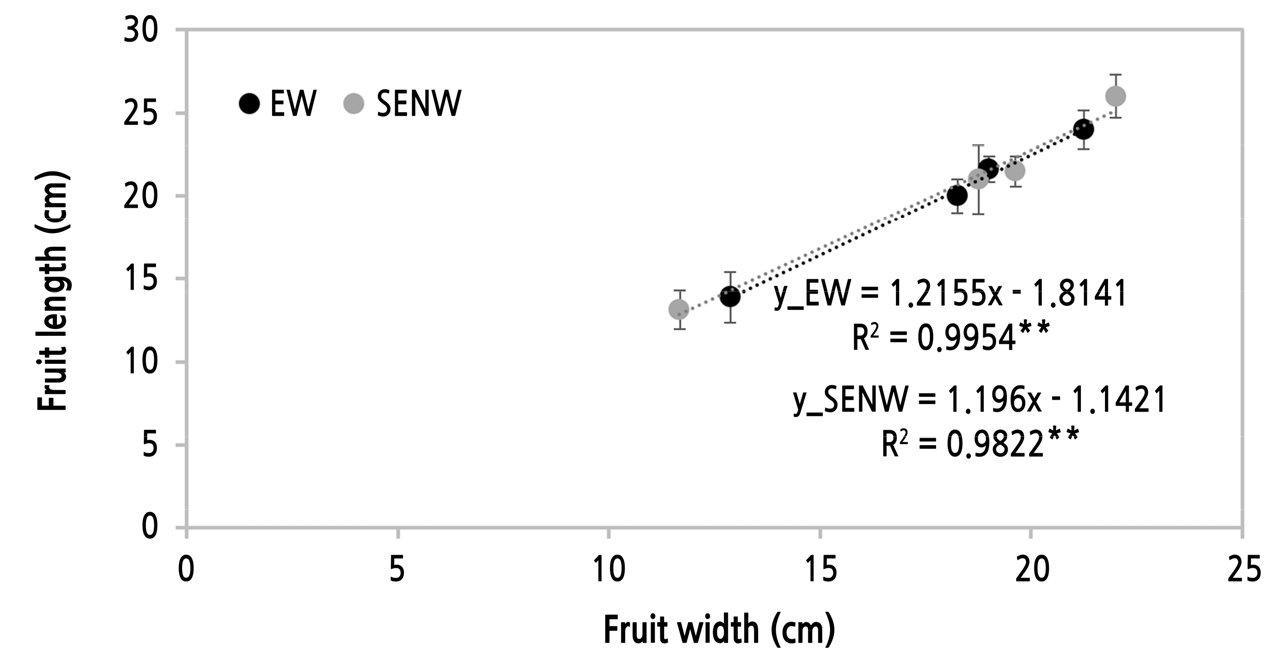
그리고 2021년 7월 15일부터 8월 4일까지 온실별 10주에 착과된 과실을 대상으로 체적량을 조사하였다. 2개의 30cm 자를 ‘ㄱ’자형으로 제작하여 일주일마다 과실의 최대 길이 및 너비를 측정하였다. 그리고 품종이 구형(Fig. 2)에 가까움이 확인되어 측정된 과실의 길이와 너비를 식 (5)에 적용하여 체적량(cm3·fruit-1)을 계산하였다.
과실 체적(fruit volume)
V : 과실 체적(cm3/fruit),
π : 3.14159265358….(= 3.14)
r : 반지름(cm) = (fruit length + fruit width)/4
통계분석
SPSS 프로그램(Version 19.0 SPSS Inc., Chicago, USA)을 이용하여 두 집단(EW, SENW 방향의 온실) 간 평균 차이 검증은 95% 신뢰수준(p < 0.05)에서 F-검정을 통한 이분산(Heteroscedasticity of variance) 또는 등분산(Homogeneity of variance) T-검정을 실시하였고, 주요 요소 간 관계는 회귀 분석(regression analysis)을 실시하였다. 그리고 회귀 그래프는Excel 2016(office 365, microsoft Crop., Canada)을 이용하여 작성하였다.
결과 및 고찰
온실 내부 환경 변화
재배 기간 동안 온실 내부의 일 평균 광도(PPFD) 및 기온을 측정하였다(Table 1). 30분 마다 측정된 값의 일 평균 광도, 일 평균 기온, 그리고 주간 평균 기온 모두 SENW 방향의 온실에서 유의하게 높았다. 일 평균 광도는 SENW 온실에서 363.2 µmol·m-2·s-1로 EW 방향 온실의 337.8 ± 293.14µmol·m-2·s-1 보다 약 25.4µmol·m-2·s-1 (7.5%) 더 높아 재배 기간 누적 광량 수준에서 큰 차이를 나타낼 것으로 판단된다. 이러한 온실 내 유입되는 광도의 차이는 기온의 차이를 유발시켰고, 이에 재배기간 동안의 적산 일사량 및 온도에도 영향을 주었을 것으로 판단된다.
온실 방향에 따라 일중 시각별 평균 광도와 재배기간 동안 일 적산일사량(Daily integrated solar radiation) 변화를 조사하였다. 일중 평균 광도는 EW보다 SENW에서 높았고 편차도 작은 경향을 나타내었다(Fig. 3A). 특히, 일출부터 14시경까지 차이를 나타내었는데, 이는 온실 방향과 태양 이동 방향 및 고도에서 오는 것으로 판단된다. Choi et al.(2008)과 Choi et al.(2010)의 토마토 온실 방향(남북, 동서)에 따른 겨울철 온실 내부의 일중 광도(PPFD) 변화 연구에서도 온실 방향에 따라서 오전 광도와 오후 광도의 변화가 월마다 차이를 나타내었다. 특히, 동지를 지나 3월로 들어올수록 남북 방향 온실에서 동서 방향 온실보다 오전 광도가 높아지고 오후 광도의 차이는 작아지는 경향이었다. 이를 고려하면 본 연구 시기인 7월에 온실 방향에 따라 EW 온실보다 SENW 온실에서 오전에 광도의 차이를 나타낸 결과도 이와 같은 결과로 생각되었다. 이는 방위각이 커짐에 따라 EW는 남쪽 측면 전체의 입사각이 작아지기 시작하는 반면 SENW는 태양과 직선으로 놓이는 시간이 많음으로써 측면 입사각이 커지기 때문으로 판단된다. 일 적산 일사량을 보면, 조사 기간 동안 SENW 온실에서 높았다. 차광을 실시한 7월 28일 이전 기간에는 평균 2.7 ± 3.2mol·m-2·day-1, 차광된 기간에는 평균 3.6 ± 3.3mol·m-2·day-1 더 높았다(Fig. 3B). 하지만 수박의 광 포화점은 80klux(약 550–600µmol·m-2·s-1)로 알려져 있는데(RDA, 2018) 이와 비교하면 본 시험 시기에는 광도가 낮았다.
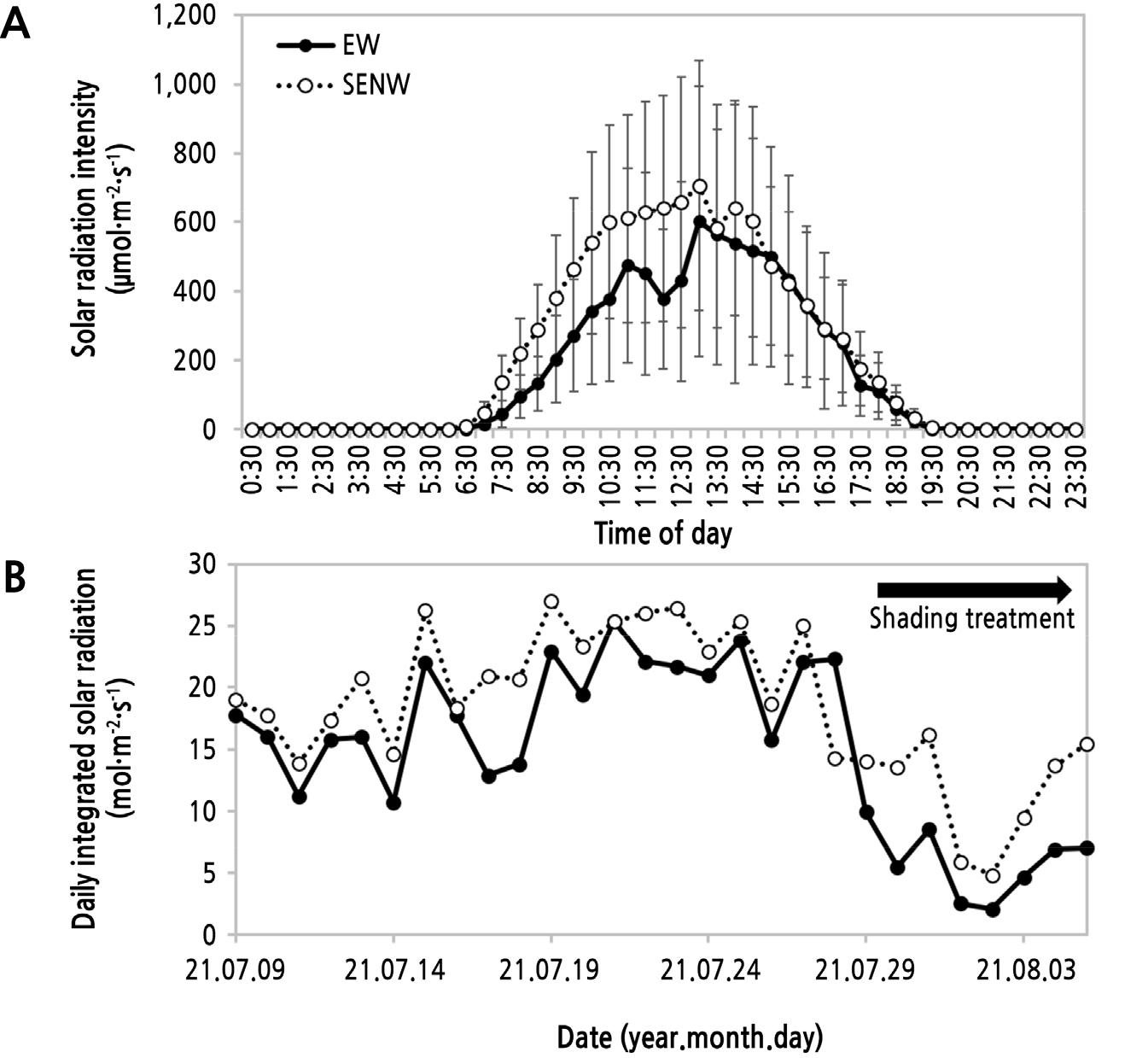
Fig. 3.
Change in internal mean solar radiation intensity as photosynthetic photon flux density during a day (A) and cultivating period (B) according to direction of single-span greenhouse planted seedless watermelon. EW, east-west; SENW, southeast-northwest. Bars are standard deviation (A, n = 28 days).
온실 방향별 기온 변화를 조사하였다. 조사 기간 동안 일중 시각별 기온변화를 보면 각 시각별로 두 온실 간에 뚜렷한 차이를 나타내지는 않았지만 EW에서 조사기간 동안 동일 시각 간 편차가 SENW보다 컸다(Fig. 4A).
기온은 광 파장에서 열원으로 작용하는 적외선에 의해 영향을 받는다. 온실 내부의 기온과 일 적산일사량을 고려하였을 때 주간온도의 차이가 뚜렷하게 나타나야 하지만 본 연구의 결과에서는 다른 결과가 나타났다(Table 1 and Fig. 4B). 시험장소인 전북 고창은 이 시기에 일몰 이후 기온의 하강에 따라 남풍이 부는 시기이며 주간에는 고온으로 인해 측창 전체를 열어 두는데 풍향을 고려하면 온실 전체 환기량은 EW 온실에서 좋았을 것으로 생각된다. 하지만 오히려 야간온도는 환기량이 많을 것으로 판단된 EW에서 높은 경향을 나타내었는데 이는 Fig. 1에서 보는 바와 같이 EW 온실 옆에는 선별 창고가 위치하고 있어 나가는 바람을 막아 야간의 환기량이 적어 온실 내부 기온의 하락이 적었을 것으로 판단된다. 본 온실과 같은 환경제어가 되지 않는 경우 내부 온도는 외부 환경에 의존적이다. 내부 온도 관리에 환기는 매우 중요한 요소로써 환경제어가 가능한 온실에서는 환기량에 대하여 외부 광량, 외부 온도, 풍량을 중요한 매개변수로 다루고 있다(Jung et al., 2020).
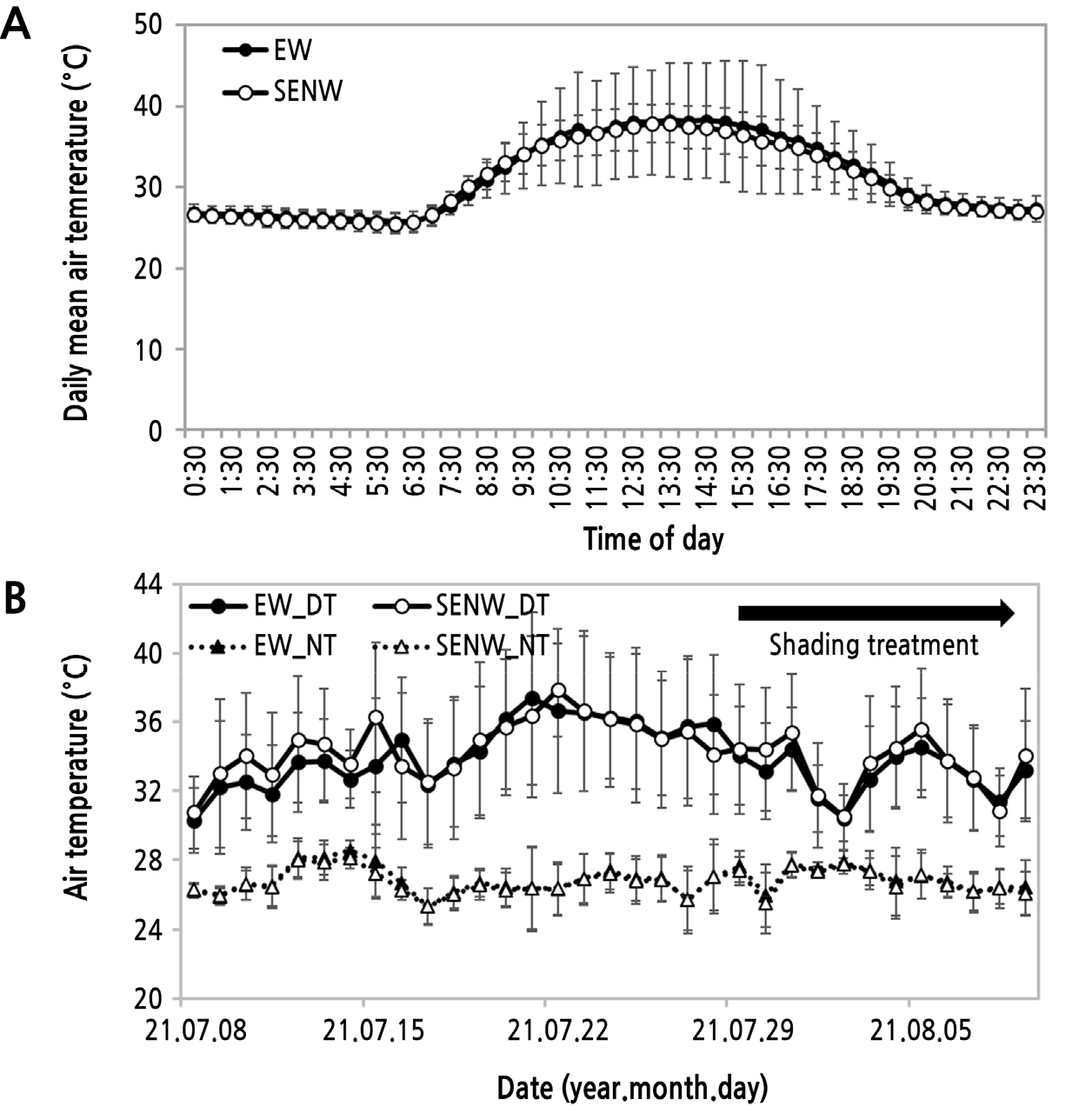
Fig. 4.
Change in internal mean air temperature during a day (A) and cultivating period (B) according to direction of single-span greenhouse planted seedless watermelon. EW, east-west; SENW, southeast-northwest; DT, daytime; NT, nighttime. Bars are standard deviation (A, n = 28 days; B, n = 22–25 times).
Table 1.
Intensity of internal solar radiation and air temperature according to direction of single-span greenhouse planted seedless watermelon
| Direction | Daily average internal radiation intensity (µmol·m-2·s-1) | Average internal air temperature (°C) | ||
| Daily | Daytime | Nighttime | ||
| EW | 337.8 ± 249.61z | 30.69 ± 4.46 | 33.84 ± 3.19 | 26.86 ± 1.19 |
| SENW | 363.2 ± 264.96 | 31.52 ± 5.33 | 34.21 ± 3.59 | 26.72 ± 1.11 |
| Significancey | * | * | * | NS |
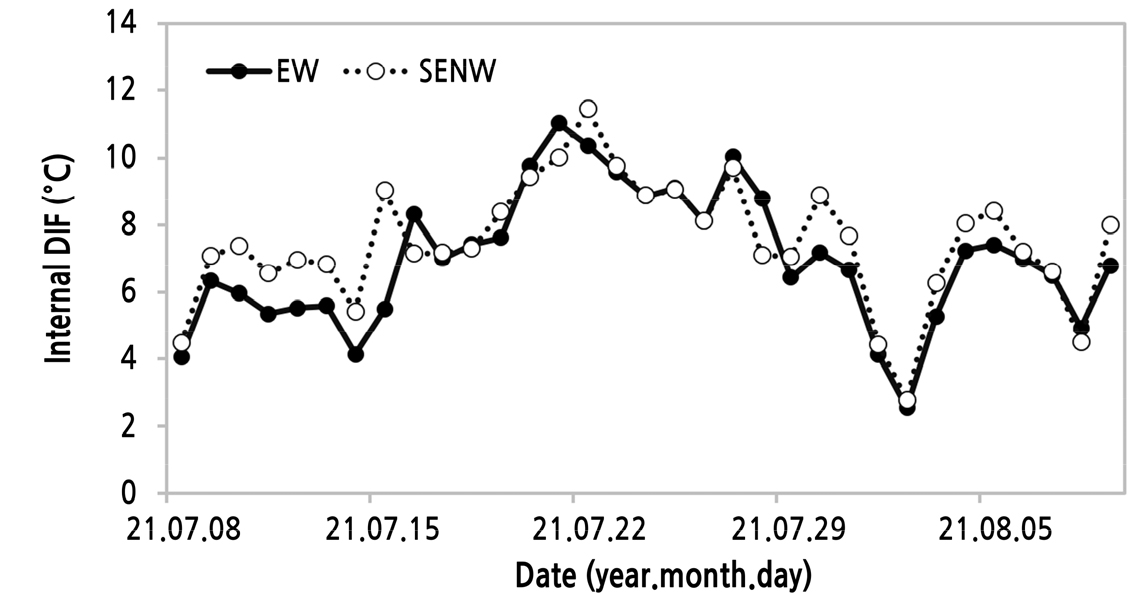
일평균 주간온도와 야간온도의 차이(주야간온도차, DIF)는 7월 상·중순에는 SENW에서 EW보다 높게 유지되었고, 이후부터 7월 하순에는 EW에서, 그리고 8월 상순에는 SENW에서 다소 높은 날이 많은 경향이었다(Fig. 5). 이러한 경향은 과실 체적 증가량(Table 7)의 결과와 유사하였다. 수박(Kwon et al., 1999)을 포함한 과채류에서 DIF에 대한 연구는 주로 육묘 과정에서 진행되었고, 정식 후 생육 기간 중 그 영향력에 대한 연구는 거의 진행되지 않았다. 파프리카의 경우에는 주간 및 야간 적온을 고려할 때 DIF가 약 5°C 내외로 판단되는데(Jeong et al., 2009a; Myung et al., 2012), 정식 후 초기 생육기에 수확된 과실 무게는 DIF 0–6°C 범위에서 차이를 나타내지 않았다고 보고되었다(Kim et al., 2013). 그리고 겨울철 토마토에서는 주간환기점을 27°C로 하였을 때 누적생산량은 야간 13–14°C 범위에서 최대라고 하였다(Kang et al., 2012). 이와 같이 작물 종류 및 시기에 따라 적합한 DIF는 다를 것으로 판단된다. 본 연구에서도 조사 기간 동안 DIF 범위는 두 방향 모두 2.0–12.0°C 범위로 시기별로 편차가 매우 심하였다. 그리고 두 방향 간에 DIF차이는 1.0–2.0°C로 큰 차이로 판단되지 않는다. 이들을 고려할 때 본 연구에서 두 온실 방향에 따른 DIF 차이 수준이 과실비대에 뚜렷한 영향을 주지 않았고 이를 구명하기 위한 면밀한 연구가 필요하다고 판단된다.
조사 기간 동안 일주일 간격으로 오전 11시경 온실 식물체 착과 부위 잎에 조사되는 광도 및 파장을 분석하였다(Table 2). 평균 PAR범위 광도(PPFD)는 EW에서 408.13, SENW에서는 825.83µmol·m-2·s-1으로 SENW 온실에서 2배 이상 높았다. 그 중 적색광은 각각 147.47과 308.16µmol·m-2·s-1, 청색광은 114.35와 221.89µmol·m-2·s-1로 PAR범위 평균 광도의 차이와 같은 차이를 나타내어 PAR 범위 광도와 높은 정의상관(R2 = 0.9962**)을 나타내었고(Fig. 6), 태양광 내 파장별 상대적 비율은 변화가 없었다. 또한, 두 온실 간 잎 주변의 광도 차이가 온실 내부 광도 차이보다 크게 나타났는데(Table 1), 이는 물체까지의 도달 시 거리에 따라 그 PPFD는 지수함수형태로 감소하기 때문으로 판단된다(Lee et al., 2014). 총 PPFD에 대한 적색광과 청색광의 비율은 두 처리 간에 차이가 없었으므로(Table 2) blue와 red 파장 강도가 기공 열림에 영향을 주었고(Lurie, 1978), 이에 따라 증산속도(Table 3)에도 영향을 주었을 것으로 판단된다.
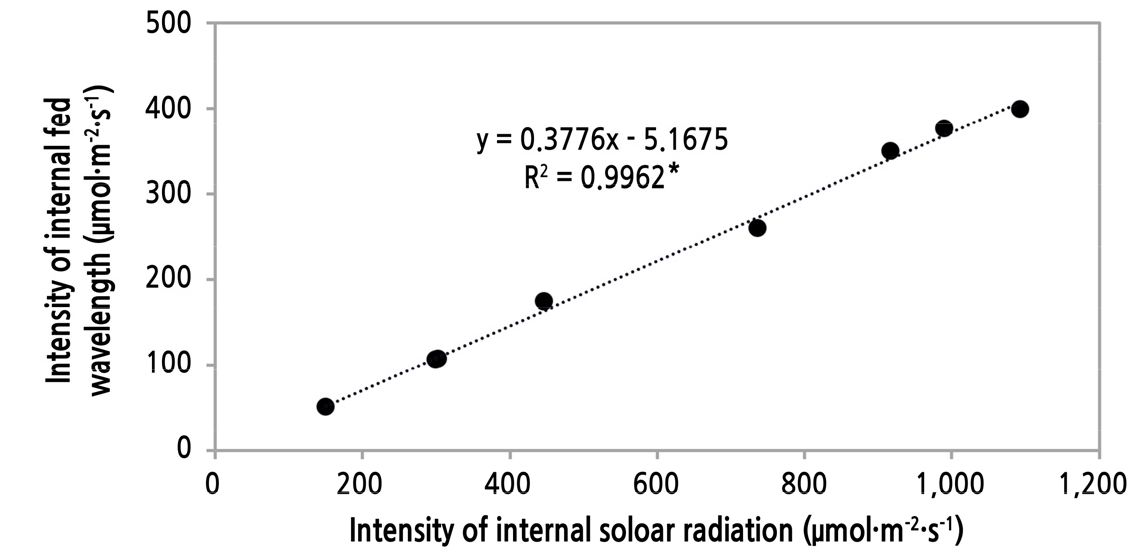
Table 2.
Total of internal solar radiation, amount and rate of red and blue wavelength in photosynthetic active radiation (PAR) range near leaves around fruits of seedless watermelon according to direction of single-span greenhouse
| Direction of greenhouse | Internal PPFD (µmol·m-2·s-1) in PAR range | |||||||
| Total of solar radiation | Red wavelength of | Blue wavelength | ||||||
| Amount | Rate (%) | Amount | Rate (%) | Amount | Rate (%) | |||
| EW (A) | 408.13 | 100 | 147.47 | 27.54 | 114.35 | 21.93 | ||
| SENW (B) | 825.83 | 100 | 308.16 | 28.23 | 221.89 | 20.71 | ||
| Significancez | * | * | - | * | - | |||
| B-A | 417.7 | - | 160.69 | 0.69 | 107.54 | 1.22 | ||
Table 3.
Average values of transpiration rate, stomatal conductance, vapor pressure deficit, and photon efficiency of leaves around fruits of seedless watermelon according to direction of single-span greenhouse from July 8 to August 4, 2021
| Direction of greenhouse |
Transpiration rate (mmol·m-2·s-1) |
Stomatal conductance (mol·m-2·s-1) |
Vapor pressure deficit (kpa) |
| EW | 4.75 | 0.68 | 0.91 |
| SENW | 6.53 | 0.64 | 1.25 |
| Significancez | * | NS | ** |
식물체의 광합성 요소
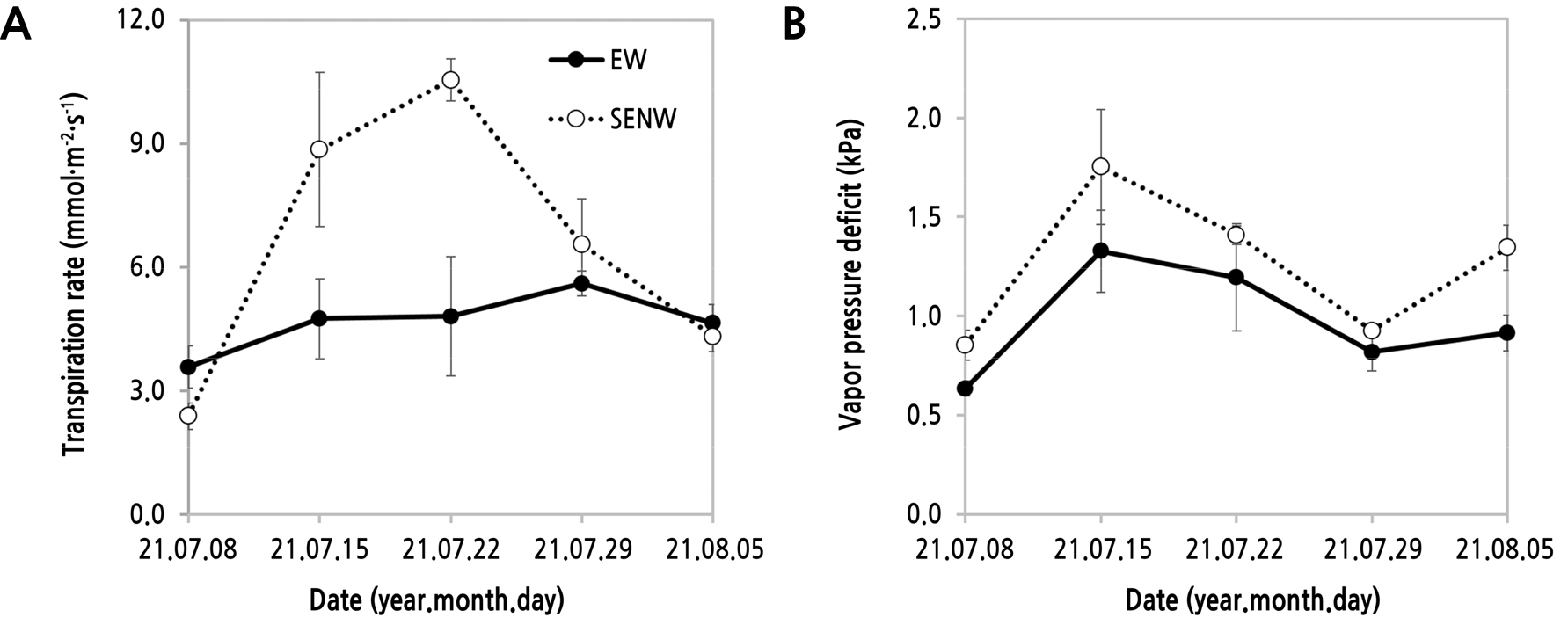
조사 기간 동안 일주일 간격으로 오전 11경의 잎의 기공전도도(stomatal conductance), 증산속도(transpiration rate), 수증기압포차(vapor pressure deficit)는 모두 Fig. 3와 Table 2에서 온실 내부 및 잎의 근접 광도가 높았던 SENW에서 EW 온실 식물체에 비해 높은 경향이었고, 잎의 기공전도도를 제외하고 증산속도(p < 0.05)와 수증기압포차(p < 0.01)는 통계적으로 유의한 차이를 나타내었다(Table 3). 특히, 증산속도는 조사 시작 2–3째주 조사(7월 15일과 22일)에서 EW 온실 식물체보다 SENW 온실의 식물체에서 각각1.86배와 2.19배 유의하게 높았다(Fig. 7). Tai et al.(2010)도 파프리카 식물체 잎의 증산속도와 수증기압포차는 일중 일사량 변화에 따라 높은 정의 상관을 보였다고 보고하였다. Shin et al.(2007)은 참외 재배 시 낮은 수증기압포차(VPD) 조건을 만들기 위해서는 기온과 광량을 낮추어야 하고, 이에 따라서 잎의 증산속도와 기공전도도도 낮아진다고 보고하였는데 본 연구에서도 유사한 결과를 보였다(Table 2 and 3). 잎에 직접 전달되는 일사는 엽온을 상승시키고 증산에 의한 잠열 손실에 의해서 다시 엽온이 내려간다(Idso, 1982; Park, 2011). 이에 대해 Bange(1953)의 연구 결과에 의하면 엽온이 내려가는 것은 내부와 외부의 수분포텐셜로 인한 잎의 수증기 함량의 변화, 즉 증산이 엽온 변화를 유발하여 이에 따라 증산속도에도 영향을 준다고 하였다.
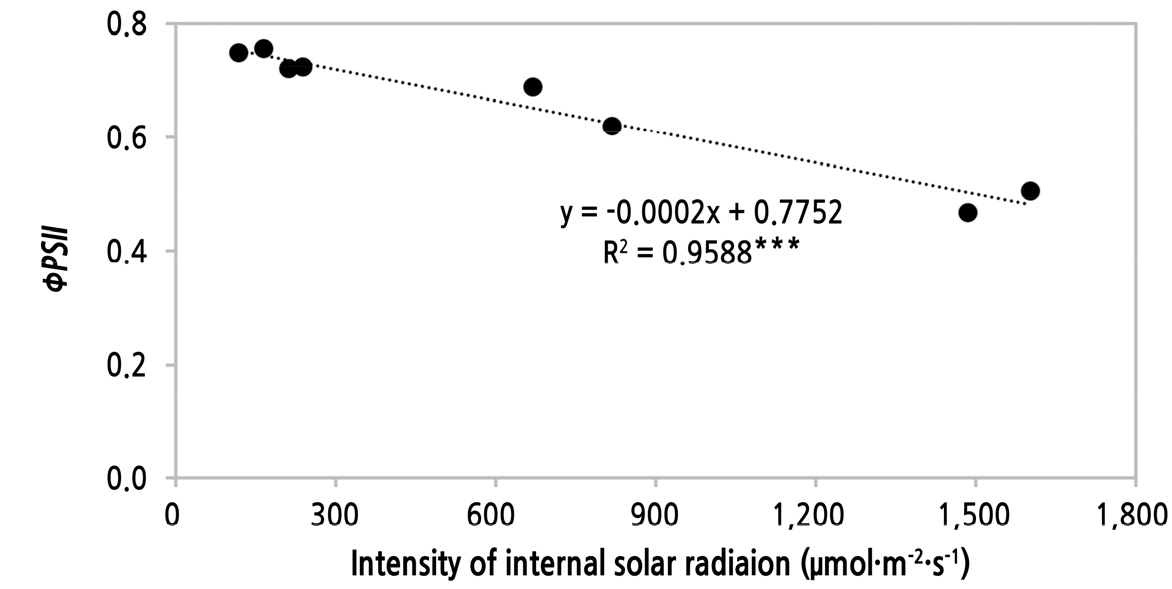
광에 적응된 상태에서 잎의 최대 형광강도(maximum fluorescence, Fm’)는 EW 온실 식물체에서 372.6 ± 22.39, 최소 형광강도(minimum fluorescence, Fs)는 SENW에서 134.2 ± 24.46로 유의하게 높았다(Table 4). 이에 따라 (Fm’–Fs)/Fm’에 의한 광계 Ⅱ에서 입사된 광량자의 광화학에너지로의 사용 효율인 ØPSⅡ은 광도가 높았던 SENW 온실에서 0.47–0.69의 범위로 측정이 되었으며 EW 온실에서 0.71–0.76의 범위로 SENW 온실보다 EW 온실의 식물체에서 유의하게 높았다(Table 1 and Fig. 3). Niinemets and Kull(2001)은 일평균 적산일사량(mol·m-2·day-1)이 증가함에 따라 광합성전자수송능력(capacity of photosynthetic electron transport, µmol·m-2·s-1)은 증가하나, PPFD가 높을수록 광계Ⅱ의 효과적인 광양자수율(ØPSⅡ)은 감소한다고 하였다. 본 연구의 수박의 잎에서도 광도가 높을수록 ØPSⅡ는 감소하는 높은 유의성의 선형 회귀모형(Fig. 8, R2 = 0.9588***)을 나타내었다.
일주일 간격으로 오전 11시경 측정된 기온과 엽온의 차이를 조사하였다(Table 5). 측정시점의 기온(p < 0.001), 엽온(p < 0.001), 그리고 엽온 - 기온의 차이(p < 0.05)는 온실 방향에 따라 유의한 차이를 나타내었다. 엽온과 기온의 차이가 SENW 온실에서 EW 온실보다 작았다. 엽온에서 기온을 뺀 값이 양의 값이면 기공전도도가 감소하고 증산이 억제된다는 연구결과가 있는데(Jackson et al., 1981; Idso, 1982), 본 연구에서는 음의 값을 나타내어 고온 작물로서 수분 스트레스가 없었던 것으로 판단되었다.
Table 4.
Average values of maximum fluorescence (Fm’), minimum fluorescence (Fs), and quantum use efficiency (ØPSⅡ) of leaves around fruits of seedless watermelon according to direction of single-span greenhouse from July 8 to August 4, 2021
| Direction of greenhouse | Fm’ | Fs | ØPSⅡ |
| EW | 373.80 | 100.05 | 0.73 |
| SENW | 322.50 | 134.22 | 0.57 |
| Significancez | ** | ** | * |
Table 5.
Leaf temperature (LT) of seedless watermelon and internal air temperature (AT), and the difference values between LT and AT (LT-AT, DV) according to direction of single-span greenhouse from July 8 to August 4, 2021
| Direction of greenhouse | EW | SENW | Significancez |
| LT (A) | 28.66 | 32.18 | *** |
| AT (B) | 37.12 | 38.34 | *** |
| DV (A-B) | –8.46 | –6.16 | * |
| Significancez | ** | ** | - |
착과 부위 잎 근처의 기온, 엽온, 엽온에서 기온을 뺀 값(DV), 광도(PPFD)와 광합성 요소인 증산속도, 수증기압포차, 기공전도도 간 상관분석 결과(Table 6), 엽온은 수증기압포차와 매우 밀접한 양의 상관성(R = 0.904***)을 나타내었고 DV와도 유의한 상관성(R = 0.0.799*)을 나타내었다. 그리고 잎에 근처에 도달되는 광도는 증산속도와 높은 양의 상관성(R = 0.856**)을 나타내었다. 광도는 증산속도 외에는 다른 요소와 유의성을 나타내지는 않았지만 엽온에 영향을 미쳤고(Fig. 9A, R2 = 6828**), 이에 엽온이 DV에 영향을 미친 것(Fig. 9B, R2 = 0.6227*)으로 보아 광 환경이 광합성 요소에 간접적으로 연관성을 나타낸 것으로 판단되었다. Espinosa-Calderon et al.(2012)은 Capsicum annuum 작물에서 챔버의 온도(23, 33, 39, 45°C) 및 광도(0–1,000µmol·m-2·s-1)을 달리한 조건에서 엽온과 기온의 차이를 연구한 결과, 챔버 온도 23°C과 33°C에서는 엽온이 챔버의 온도보다 낮았고 광도가 높아질수록 두 온도의 차이는 커지는 경향을 나타내었다. 하지만 챔버온도 39°C와 45°C에서는 엽온이 챔버온도보다 높았고 그 차이는 광도가 높아져도 초기 차이와 유사하게 유지되는 경향을 나타내었다. 그리고 엽온이 챔버온도보다 낮은 23°C과 33°C 처리에서 암반응 단계에서 CO2를 RuBP에 고정시키는 카르복실화효소인 Rubisco의 양이 일정 광도에서 증가하기 시작하여 광도의 증가에 따라 그 양도 증가하는 경향을 나타내었고 이러한 결과가 광합성율에 영향을 미쳤다고 보고하였다. 이 기존 결과를 고려할 때 두 온실 간의 광도 및 적산 일사량 차이(Fig. 3 and Table 2)가 증산속도나 수증기압포차 등 광합성 요소의 차이(Table 3)를 유발한 것으로 판단된다.
Table 6.
Correlation analysis between internal environmental factors near leaves around fruits and transpiration rate (TR), vapor pressure deficit (VPD), and stomatal conductance (SC) of the leaves of seedless watermelon in single-span greenhouse from July 8 to August 4, 2021
| Factors | TR (mmol·m-2·s-1) | VPD (kPa) | SC (mol·m-2·s-1) |
| Leaf temperature (A) | 0.540z | 0.904** | –0.223 |
| Air temperature. (B) | 0.487 | 0.422 | 0.070 |
| DV (A-B) | 0.299 | 0.799* | –0.329 |
| Solar radiation | 0.856** | 0.689 | 0.297 |
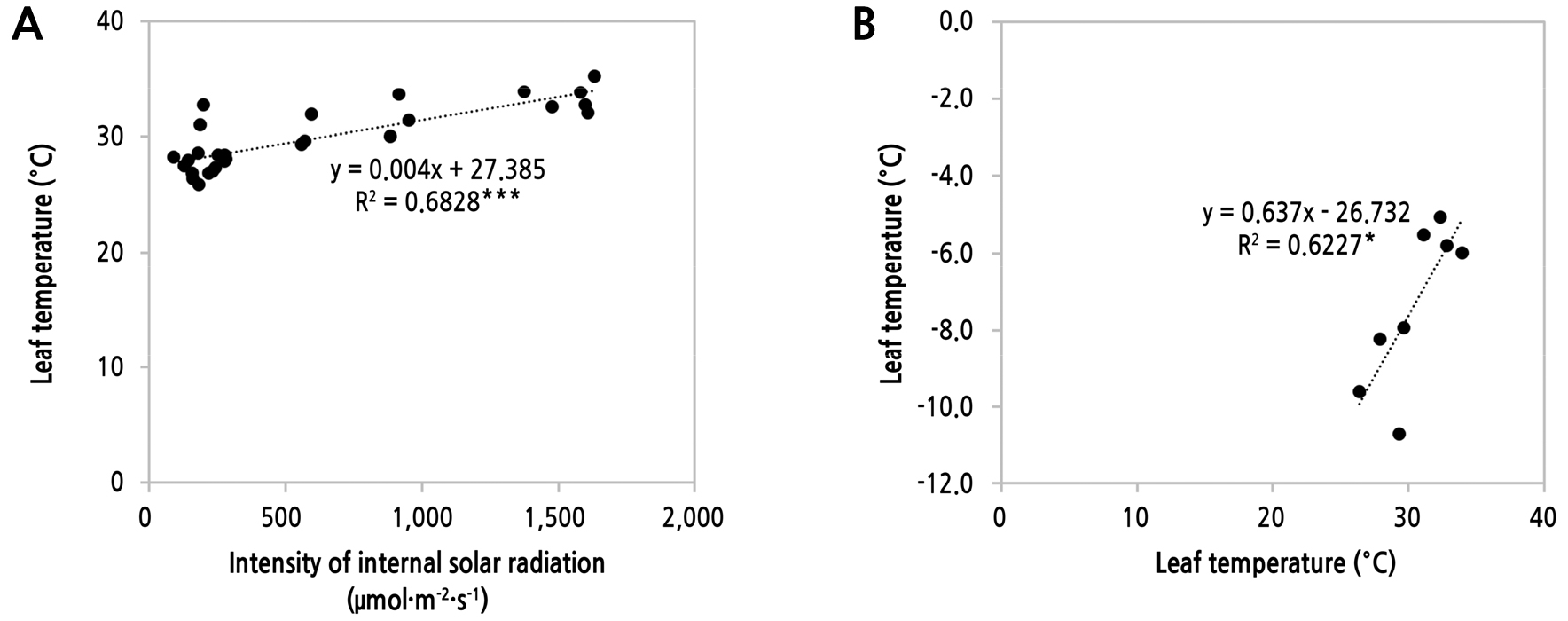
Fig. 9.
Linear regression between internal solar radiation intensity of greenhouse and leaf temperature of seedless watermelon (A), the leaf temperature and difference between the leaf temperaure and internal air temparature (DV) (B) according to direction of single-span greenhouse. *,**Significant at p < 0.05 and 0.01.
과실 비대
수박의 과실생장 곡선은 단일시그모이드곡선(single sigmoid curve)을 이루는 것을 고려하여 과실의 비대가 활발한 시기인 7월 15일부터 8월 5일까지 일주일 간격으로 과실의 직경과 길이를 조사해 체적으로 환산한 후 일주일간의 증가량을 조사하였다(Table 7). 조사 2주–3주차 기간동안 EW 온실에서 SENW 온실에 비해 342.5cm3·fruit-1 유의하게 더 많았던 것을 제외하고 다른 기간에는 SENW 온실에서 각각 794.2와 1,019.7cm3·fruit-1 더 많았다. 이에 따라 4주간 총 체적 증가량은 SENW 온실 과실에서 EW 온실 과실보다 평균 1,471.5cm3·fruit-1 더 많았다.
Table 7.
Increment of fruit volume in fruit enlargement of seedless watermelon by period according to direction of single-span greenhouse from July 8 to August 4, 2021
| Direction of greenhouse | Increment of fruit volume (cm3·fruit-1) per period | |||
|
1st–2nd (Jul. 15–21) |
2nd–3rd (Jul. 22–28) |
3rd–4th (Jul. 29–Aug.04) | Sum (1st–4th) | |
| EW (A) | 2,363.7 | 700.7 | 1,691.0 | 4,755.3 |
| SENW (B) | 3,157.9 | 358.2 | 2,710.7 | 6,226.8 |
| Significancez | * | ** | * | ** |
| A-By | –794.2 | 342.5 | –1,019.7 | –1,471.5 |
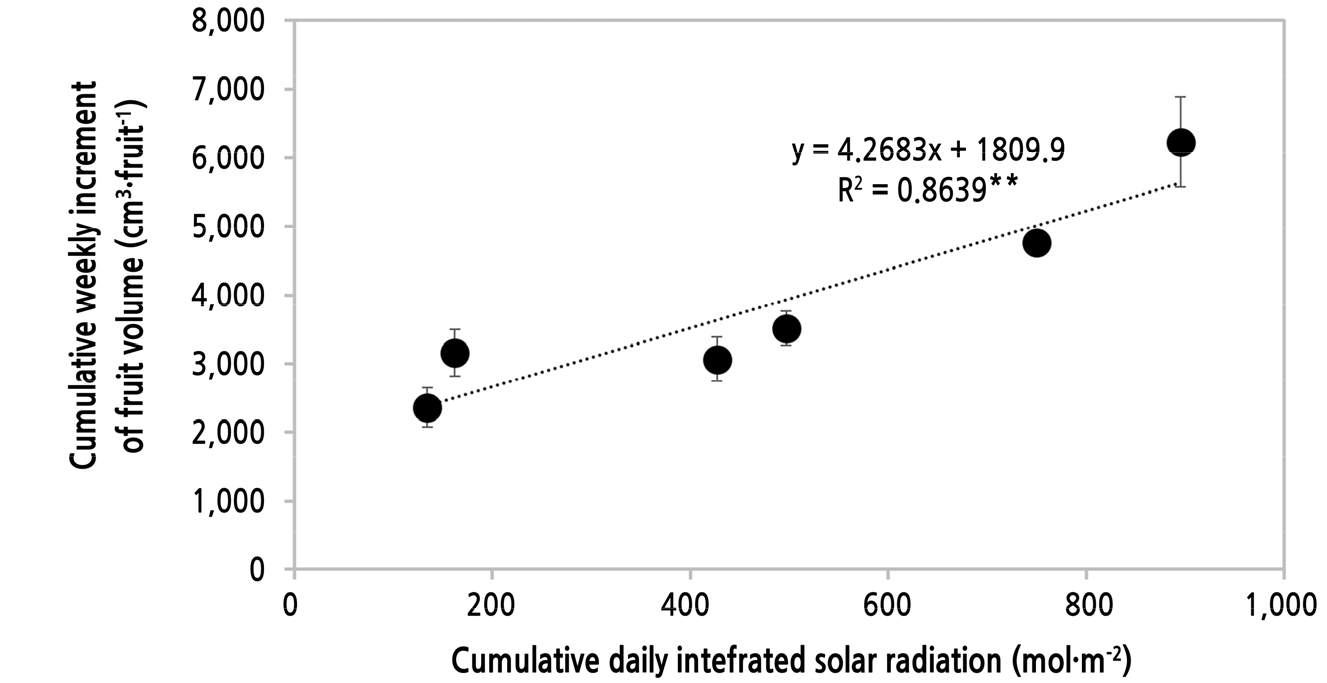
온실 방향에 따른 환경 요소, 광합성 생리 요소의 결과들을 고려할 때 수박의 비대 차이는 온도보다는 광 조건 차이로 유발된 광합성 생리 활성 정도 차이가 결국 과실 체적 증가량의 차이를 유발한 것으로 판단된다. 이에 따라서 과실이 비대하는 동안 조사된 주별 과실 체적증가량 및 주별 적산일사량의 누적값을 이용하여 선형 회귀분석을 하였다(Fig. 10). 수박 과실의 비대는 적산일사량의 증가에 따라 유의하게 영향을 받았으며(R2 = 0.8639**) 적산일사량이 1mol·m-2 증가할 때 4.2683cm3·fruit-1 증가하는 선형회귀식을 나타냈다.
본 연구의 결과를 종합하면 SENW 온실에서 조사 기간 동안 일중 광도 및 적산일사량이 높았으며, 이에 따라 증산속도의 차이를 유발하여 동화산물의 생산량 차이를 나타낼 수 있던 것으로 판단된다. 따라서 평지에서 고온 작물인 수박을 재배할 시 단동 온실을 동서보다는 남북 방향으로 짓는 것이 광 환경 및 비대 촉진에 유리할 것으로 판단된다.
