

서 언
재료 및 방법
실험 장소 및 재배 방법
재배환경 및 생육 측정
다중 선형회귀분석 및 통계분석
결과 및 고찰
온실 내·외부 환경 특성
생육 특성과 단위 면적당 생산량 변화
환경요인과 생산량 간 상관관계 분석
서 언
파프리카(Capsicum annuum L.)는 경기, 충청, 경남, 전라도 지역에서 8–9월에 정식하여, 다음 해 7월까지 수확하는 겨울 작형과 일부 고랭지와 강원도에서 3–5월에 정식하여, 5–12월 수확하는 여름 작형으로 생산되고 있어 연중 생산이 가능하다. 2020년 국내 파프리카 총수출량은 30.3천 MT로 매월 평균 2.5천 MT를 수출하였으나 1–3월과 8–10월의 수출량은 평균 1.5천 MT로 비교적 낮은 편이다(KaTi, 2022). 1–3월과 8–10월의 낮은 수출량은 저온, 저일조, 고온 등의 환경이 생산성 저하를 가져온 것으로 알려져 있다(Adams et al., 2001; Lee et al., 2014; Kim et al., 2021). 특히 여름 재배의 생산량이 겨울 재배의 생산량보다 20% 낮아(Cho et al., 2009), 8–10월의 생산량 증대 기술이 필요하다.
파프리카는 전 재배 기간에 걸쳐 영양생장과 생식생장이 동시에 이루어지는 작물로 광량, 온도, 습도, 이산화탄소 등 환경요인에 따라 생육상이 결정된다(Bakker and Uffelen, 1988). 착과기에는 생식생장을 유도하고, 과실비대기에는 영양생장을 유도하는 관리가 필요하고 생육상과 생산량이 환경에 예민하게 반응하기 때문에 균일하고 정밀한 환경관리가 필요하다(Marcelis et al., 2004; Heuvelink et al., 2002). 균일하고 정밀한 환경관리는 광, 온도, 습도 등의 지상부 환경요인과 근권 온도와 수분 등 지하부 환경요인이 복합적으로 상호작용하는 환경 관리를 말한다. 그 중 가장 큰 영향을 미치는 요인으로 광과 온도가 제시되었으며 이들 요인이 생산량에 영향을 미친다는 선행연구 결과가 보고되어 있다(Rylski and Spigelman, 1982; Myung et al., 2012; Woo et al., 2014; Myung, 2016).
지금까지 국내 파프리카 연구는 개별 환경요인이 파프리카의 생육과 생산성에 미치는 영향을 구명하여 온실 내부의 한계점을 극복하는 노력을 해왔다. 특히 여름철 온실 내부 환경은 장마와 잦은 강우로 일조량이 낮아지는(Jeong et al., 2009) 반면 온도는 30°C 이상의 고온이 지속되어 작물의 광합성량 부족과 낙화 또는 착과 불량, 과실 비대 발육 미숙이 발생한다(Marcelis et al., 2004; Cho et al., 2009; Myung, 2016). 이처럼 생육과 개화, 착과 등에 온도가 크게 관여하기 때문에 생육 단계별 온도 관리가 필요하여 정식 후 영양생장기에는 21–23°C, 착과 후 과실비대기에는 21°C로 관리한다(Bakker and Uffelen, 1988). 25°C 이상에서 작물은 유지호흡(maintenance respiration)을 증가시켜 낙화 또는 낙과율이 높아지고, 34°C 이상의 고온에서 개화 전에 낙화한다(Rylski and Spigelman, 1982). 낙과율은 주간보다는 야간 온도에 큰 영향을 받는 것으로 알려졌으며(Rylski and Spigelman, 1982; Bakker, 1989), 특히 야간 온도가 높고 일조가 낮을 때에는 꽃의 영양상태 불량으로 수분 및 수정이 불량해지고 결국 낙화 또는 낙과하게 된다(Marcelis et al., 2004; RDA, 2020).
국내에서는 시설 내부 환경관리를 위하여 여름철 차광(또는 차광제)과 시설 구조 또는 덕트와 같은 냉방설비를 통한 온도 조절과 겨울철 보광을 활용한 광량 조절을 시도하고 있다. Lee et al.(2014)은 LEP(Lighting Emitting Plasma)램프로 보광했을 때 유효 광량이 증가하여 생산성 증대가 가능하다고 보고하였으며, Ha et al.(2012)은 여름철 고온기에 30% 차광률의 차광제를 온실 외부 지붕에 살포하였을 때 온실 내부온도 감소 효과와 엽온 2°C 하강, 과실 온도 5°C 하강, 꽃의 온도 3°C 하강의 효과를 보고하였다. Kim et al.(2021)은 지열과 팬 앤 패드로 냉방이 가능한 반 밀폐형 온실로 시설의 구조를 변경하였을 때 하루평균 온도가 일반 플라스틱 온실보다 약 6°C 낮다고 발표하였다. Choi et al.(2013)은 공기 순환 덕트를 활용한 근권부 냉방 24시간 연속 냉방 하였을 때 온실 상부 4.4–5.1°C, 하부 2.1–3.1°C 하강하였고, 근권온도 22.8°C로 대조구 27.7°C에 비하여 현저히 낮다고 보고하였다. Lee et al.(2022)는 파프리카 수경재배에서 근권 냉방이 무냉방 보다 근권온도를 4–8°C 낮출 수 있으며 특히 15°C의 차가운 배양액을 급액 했을 때 효과가 우수하여 뿌리 건물중과 광합성률을 상승시킨다고 하였다. 그러나 상기한 방식은 광과 온도 개별로 관리하는 방식으로 복합적으로 상호작용하는 환경이 식물체에 주는 영향에 대한 이해가 부족하다.
최근에는 상호작용하는 온실 환경이 작물에게 미치는 영향에 관한 연구가 수행되고 있다. Myung(2016)은 파프리카 재배에서 이산화탄소와 일평균 온도, 재식밀도와 광량 등 지상부 환경과 EC농도와 같은 지하부 환경 동시 개선을 통한 생육과 생산성 증대 연구를 시도를 하였다. 그의 결과에 따르면 수광률이 높은 외줄기 재배를 하면서 외부 광량에 따라 이산화탄소와 EC를 감소시키고 초저녁 냉방 처리로 일평균 온도를 감소하면 생산량 증대가 가능하다. 그러나 식물체에 미치는 각 요인의 영향 정도와 관계성을 분석하지는 못하였다. 따라서 본 실험에서는 강원도 여름 작형 파프리카 생산 시 생산성에 영향을 미치는 다양한 환경 요인과 그 중 영향력이 높은 환경 인자(Influence factor)를 구명하고자 하였다.
재료 및 방법
실험 장소 및 재배 방법
본 실험은 강원도 소재 인제 군 1개소(FIn)와 철원 군 1개소(FCh)의 플라스틱 연동 온실에서 2021년에 수행하였다. 분석 농가는 적색계 파프리카를 재배하였으며 품종, 재식밀도, 정식일 등은 온실에 따라 달랐다(Table 1). FIn는 2021년 2월 1일 정식하여 11월 13일에 종료하였으며 암면(L × W × H : 120 × 12 × 8cm, Grodan, Netherlands) 수경재배로 3줄기 유인하였다. FCh는 2021년 3월 14일 정식하여 11월 11일에 종료하였으며, 코이어 배지(L × W × H : 100 × 15 × 14cm, Duck Yang, Sri Lanka)에 2줄기 유인하여 수경재배하였다. 관수 관리는 관행 방식의 누적일사량제어 및 관리를 따랐고, 재배환경 관리는 농가의 관리방식에 따라 수행되었다.
Table 1.
Information of surveyed sweet pepper export farms in Injae (FIn) and in Cheorwon (FCh) in Gangwon-do, South Korea in 2021
재배환경 및 생육 측정
재배환경 분석자료는 농가에 설치된 복합환경제어시스템(MAXIMIZER 4.2.0. build 4771 Version, Priva B. V., Netherlands)을 사용하여 온실 내·외부 온도와 상대습도, 일사량을 5분 단위로 계측한 자료를 사용하였다. 파프리카의 생육과 생산량 특성은 첫 개화 후 10월까지 2주 간격으로 초장과 마디 수, 엽장과 엽폭, 착과 수, 화방 높이, 수확 과실 무게 및 수량을 조사하였다. 표준화된 매뉴얼(GARES, 2019)에 준하여 초장은 지제부부터 생장점까지 길이를, 마디 수는 방아다리부터 생장점까지 마디를 조사하였다. 1cm 이상의 과실을 착과로, 화방 높이는 생장점부터 첫 번째 개화 화방까지 길이를 조사하였고, 생장점 아래 3번째 완전히 전개한 잎의 엽장과 엽폭을 측정하였다. 착색이 90% 이루어진 과실을 수확하여 수량과 무게를 측정하였다. 조사는 온실별 6개의 파프리카 개체를 표본조사 하였으며 표본은 임의 개체 선발하였다.
생장 억제 정도를 나타내는 지표로 ‘상대적 절간 비율(Eq. 1)’과 착과 효율성을 나타내는 지표로 ‘착과율(Eq. 2)’, 생산성을 나타내는 지표로 ‘착과 대비 수확률(Eq. 3)’을 산정하였다. 각 산출 값은 식물체의 8마디씩을 1그룹으로 하여 총 5그룹(40마디)으로 구분하여 Jang et al.(2016)의 방법에 따라 분석하였다. 각 그룹 발생 시기는 1그룹은 3–5월 초, 2그룹은 5월 중순–6월 초, 3그룹은 6월 중순–7월, 4그룹은 8–9월, 5그룹은 10월이었다. 단위 면적당 주간 생산량은 조사일에 표본 개체에서 과실을 수확하여 해당 주차(week)의 생산량을 산출하였다(Eq. 4).
다중 선형회귀분석 및 통계분석
다중 선형회귀 분석과 통계분석에 사용한 데이터는 두 온실의 개화일부터 마지막 조사일로 제한하여 FIn는 2021년 3월 4일–11월 13일, FCh는 2021년 3월 31일–11월 11일의 데이터를 사용하였다. 데이터 유의성 검정은 SPSS(IBM, New York, NY, USA)를 이용하여 표본의 기술 평균과 분산을 신뢰수준 95%에서 비교 분석하는 독립표본 T 검정 방식으로 수행하였다.
영향력 환경인자 분석은 다중 선형회귀모델을 사용하였다. 다중 선형회귀모델은 종속변수에 1개 이상의 영향인자가 있을 때 분석하기 용이한 모델로(Hwang et al., 2021), 1개 이상의 환경인자가 생산량에 갖는 영향력을 분석하기에 적합하다. 생산량은 온실별 6개 표본 식물체에서 한 주에 수확한 과중을 합산하여 y = [yp1, yp2, …, ypi]의 값을 갖는다. P1은 첫 수확 주차이고, Pi는 마지막 주차이다. 환경요인 e는 생산량과 같이 생산 첫 주(T1)부터 마지막 주차(Ti)까지 e = [eT1, eT2, …, eTi]의 값을 갖는다. 주차마다 구간(interval)의 y와 e가 주어지는데, 구간은 착과일부터 수확일이고, 해당 기간의 환경요인인 주간 온도, 야간 온도, 생육도일(Growing Degree Days, GDD), 하루 평균 일사량을 누적하여 활용하였다. 식물이 자라는데 필요한 최소한의 온도를 나타내는 생육도일은 일평균 온도에 파프리카의 기본 온도 10°C를 뺀 값(Marcelis et al., 2006)을 사용하였다. FIn와 FCh를 합친 전체 생산 주수인 i는 20이었다.
실험에서 사용한 다중 선형회귀모델은 아래와 같다(Eq. 5). 다중 선형회귀 분석은 종속변수(생산량) y를 설명하기 위하여 독립변수 x1, x2,, … , xi를 사용하며, 본 실험에서는 x주간온도, x야간온도, xGDD. x일사량이다. 다중 선형회귀분석 시 변수 선택법은 모든 요소를 포함하여 수확된 과중을 설명하기 위해서 ‘입력’을 사용하였다.
y = production (g), β0 = Constant
x1 = day temperature (°C), x2 = night temperature (°C)
x3 = GDD (°Cd), x4 = solar radiation (MJ·m-2)
ε = 0
결과 및 고찰
온실 내·외부 환경 특성
조사 기간 FIn와 FCh의 온실 외부 온도는 FIn가 15.1°C, FCh는 17.1°C로 FIn의 평균 기온이 더 낮았으나, FIn와 FCh의 실내 일평균(24시간 평균) 온도, 주/야간 평균 온도, 하루 평균 적산일사량은 각 21°C, 21–23°C, 18–19°C, 15MJ·m-2로 비슷한 수준으로 파프리카 생육 적정 범위였고, 습도는 FIn가 81.5%, FCh가 94.1%로 나타났다(Table 2).
Table 2.
Average temperature, average relative humidity (RH) and average integrated solar radiation (ISR) of the surveyed farms in Gangwon-do, South Korea, for the entire cultivation period in 2021
| Farm Code | Average temperature (°C) | average RH |
Average ISR (MJ·m-2·day-1) | |||
| Outside | 24h-mean | Day | Night | (%) | ||
| FIn | 15.1 ± 6.7z | 21.4 ± 2.0 | 23.5 ± 2.4 | 18.8 ± 1.5 | 81.5 ± 9.8 | 15.1 ± 7.4 |
| FCh | 17.1 ± 6.3 | 21.9 ± 2.0 | 21.9 ± 2.0 | 19.3 ± 2.0 | 94.1 ± 7.6 | 15.1 ± 7.2 |
| Significancey | ** | * | *** | * | *** | n.s. |
하루 평균 적산일사량은 재배 기간에 걸쳐 주간으로 큰 변화폭을 보였다. 첫 개화 화방이 과실로 발달하는 시기의 초기 광량(개화 후 1주)은 FIn와 FCh가 각 13.6, 17.1MJ·m-2로 시작하여 지속해서 높아져 FIn는 19주(7/9–7/15)에 24.3MJ·m-2·d-1, FCh는 16주(7/14–7/20)에 23.3MJ·m-2·d-1까지 높아졌다(Fig. 1A). 적산일사량 정점 이후 일사량이 급격하게 떨어지는 경향을 보였는데 FIn는 21주에 64.6% 수준으로, FCh는 17주에 61.8% 수준이었다. 그 뒤 적산일사량은 꾸준히 낮아져 FIn는 24주(8/13–8/19)에 7.6MJ·m-2, FCh는 20주(8/11–8/17)에 6.2MJ·m-2까지 낮아졌다. 이와 같은 일조량의 변화는 3–5월 정식하여 11월에 종료하는 여름 작형의 특성으로 재배 기간 중간인 7월 일사량이 가장 높지만 장마와 잦은 강우로 흐린 날이 많아 8월은 일사량이 급격하게 떨어지는 경향을 보인다. 이후 재배 후반기로 접어드는 9월부터는 계절이 변하면서 서서히 일사량이 낮아진다. 2021년 인제와 철원 지역의 장마는 7월 3일부터 19일까지(KMA, 2022)로 평년보다 장마 일수가 짧아 장마 기간에도 일사량이 확보되었으나 8월에 이틀에 한 번 강우로 흐린 날이 많았다(KMA, 2021).

이에 따라 재배 상·하반기 적산일사량에서 차이를 보였다. FIn의 상·하반기 총적산일사량은 각 2090.7MJ·m-2, 1582.5MJ·m-2, FCh는 각 1915.5MJ·m-2, 1315.9MJ·m-2로 하반기 총적산일사량이 상반기 대비 75.7%, 68.7% 수준에 그쳤다(Fig. 1B). 이와 같은 경향은 상·하반기 평균 적산일사량에서도 나타나 FIn가 상·하반기에 각 17.1MJ·m-2, 13.6MJ·m-2, FCh는 각 17.8MJ·m-2, 13.0MJ·m-2이었다(Fig. 1B). 첫 착과를 시작하는 파프리카의 최소 광 요구량은 2.3MJ·m-2·d-1이고, 성숙한 파프리카는 최소 22MJ·m-2·d-1(Marcelis et al., 2004; Myung, 2016; Jeeatid et al., 2017)로 알려져 있다. FIn와 FCh의 일사량은 첫 착과를 시작하는 시기에 최소 광 요구량보다 월등히 높았으나 파프리카가 충분히 성숙한 재배 하반기는 최소 광 요구량 대비 평균 약 60% 수준으로 분석되었다.
FIn와 FCh의 전 재배 기간 일평균(24시간 평균) 온도와 주/야간 온도는 파프리카 생육적온인 주간 25–27°C, 야간 18–20°C, 일평균 21°C(Bakker and Uffelen, 1988)에 적절하게 관리된 것으로 나타났다(Table 2). 그러나 개화 후 주간 일평균 온도변화에서는 FIn와 FCh 모두 특정 3–4주간 고온으로 유지되어 최대 25–26°C까지 상승하였다(Fig. 2). 개화 후 발생한 고온기는 FIn와 FCh의 정식 시기가 달라 개화 후 주차에 차이가 있었으나 모두 7월이었다. Jeong et al.(2009)이 한국과 네덜란드의 시설 내·외부 환경을 비교 분석한 결과에서도 네덜란드는 전 재배 기간에 큰 변화 없이 18–19°C로 균일하게 관리되지만 우리나라는 22–25°C로 변화폭이 크다고 하였다.

매일 시간별 주/야간 온도변화 분석 결과에서도 하루 중 매시간 적정온도 범위를 벗어나는 현상을 관찰할 수 있었다. FIn와 FCh는 대체로 주간 온도 적정 범위를 잘 유지하였으나 주간 28°C 이상, 야간 21°C 이상으로 적정 범위를 웃도는 날 수가 적지 않았다. FIn의 주간 온도는 상반기 72일, 하반기 80일 동안 28°C 이상이었고 7월 7일부터 8월 12일까지 매일 30°C를 넘었다(Fig. 3A). 야간 온도는 상반기 4일, 하반기 52일 동안 21°C 이상이었고, 주간 온도와 마찬가지로 7월 7일부터 8월 8일까지 매일 발생하였으며 최고 26°C까지 상승하였다(Fig. 3B). FCh의 주간 온도는 상반기 39일, 하반기 28일 동안 적정온도를 웃돌았고, 야간 온도는 상반기 30일, 하반기 43일 동안 적정온도를 웃돌았다(Fig. 3C and 3D). FIn와 마찬가지로 7월 12일부터 8월 2일까지 주간 온도가 매일 30°C를 넘었고, 7월 4일부터 8월 13일까지는 야간 온도가 매일 높았으며 최고 26°C까지 상승하였다. 하루 중 최소 1시간에서 최대 10시간 고온이 지속되었으며, 야간 온도가 24°C 이상으로 상승한 날은 FIn가 11일, FCh가 21일이었다(Data not shown).

생육 특성과 단위 면적당 생산량 변화
파프리카의 생육 특성을 관찰하기 위하여 1월 1일 기준으로 38주 차(9월 하순)에 생육 조사를 수행한 결과, FIn의 초장이 314.8cm로 FCh의 286.1cm보다 길었다 (Table 3). 전 재배 기간 조사된 생육지표의 평균으로 나타낸 평균 주간 생장길이는 FIn와 FCh가 각 8.4cm, 8.9cm, 화방 높이는 FIn 5.0cm, FCh 6.0cm, 줄기직경은 FIn 4.9mm, FCh 5.6mm로 FIn가 FCh 보다 짧고 가늘었다. 엽폭 차이는 없었으나 FIn의 엽장이 17.9cm로 FCh의 18.7cm 보다 짧았다. 초장만 FIn가 FCh보다 길게 나왔는데, FIn의 생육기간이 길기도 하지만 하루평균 온도가 1.6°C 높아지면 온도 차이로 인한 동화산물 생성과 전류의 차이로 초장과 절간 수가 각 약 20%, 15% 증가한다고 알려져(Park et al., 2012), FIn와 FCh의 온실 내부온도 차(약 0.8°C)로 초장 차이가 발생한 것으로 생각된다. 그러나 FIn와 FCh의 생육지표를 보면 초장 외 지표에서 FCh가 유의하게 높게 나온 것으로 보아 FCh는 FIn와 비교했을 때 조금 더 영양생장을 한 것으로 보인다. 파프리카의 영양생장을 유도하는 요인으로 고온, 고광 또는 약광으로 인한 낙화 또는 착과 불량이 알려져 있으나(Bakker, 1989; Marcelis et al., 2004; Choi et al., 2013) 본 실험에서는 온도와 광량에서 FIn와 FCh 간 뚜렷한 차이가 없었으므로 품종의 특성, 수경재배 급액 관리, 작물 초세 관리 뿐 아니라 온실 내 환경조절장치 등이 FIn과 FCh의 생육 특성에 영향을 주었을 것으로 생각된다(Kim et al., 2012). FIn은 온실 내 유동 팬이 설치되어(Table 1) 고온기 초저녁 온도 하강 효과가 FCh보다 컸을 것으로 보이며, 실제 FIn는 주간 온도가 FCh보다 0.8°C 높지만 야간 온도는 FCh보다 0.5°C 낮았다(Table 2). FIn의 유동 팬이 FCh 보다 큰 DIF를 발생시켜 초장이 길어진 반면, 지속적 온실 내부 공기 순환으로 생식 생장을 유도한 것으로 보인다. 식물체당 일일 평균 급·배액량은 FIn이 각 1060.1mL, 231.7mL, FCh가 각 918.0mL, 395.9mL로 FCh의 배액률이 FIn 보다 높았다(data not shown). 배액률은 수경재배에서 사용한 코이어 배지의 구성 비율(Chip과 Dust의 비율)에 따라 달라질 수 있어 식물의 생육과 배지 함수량 편차 분석이 필요하나 본 실험에서는 환경요인을 중점적으로 분석하였다.
Table 3.
Shoot growth characteristics on average of sweet pepper grown in FIn and FCh
| Farm Code | Plant heightz | Average growth length | Length of flowering | Stem diameter | Leaf size (cm) | ||
| (cm) | (cm/week) | (cm) | (mm) | Length | Width | ||
| FIn | 314.8 ± 12.6y | 8.4 ± 0.8 | 5.0 ± 0.8 | 4.9 ± 0.3 | 17.9 ± 0.5 | 10.5 ± 0.4 | |
| FCh | 286.1 ± 14.9 | 8.9 ± 0.7 | 6.0 ± 0.9 | 5.6 ± 0.4 | 18.7 ± 0.6 | 10.8 ± 0.5 | |
| Significancew | *** | * | ** | *** | *** | NS | |
생장 억제 정도를 나타내는 지표인 상대적 절간 비율 분석 결과, FIn은 1그룹에서 138%로 생장을 하다가 2그룹부터 95%, 93%, 80%, 83%로 생장이 억제되었다(Fig. 4). FCh는 1그룹에서 112%로 생장을 하였지만 FIn 보다 유의하게 낮았고, 2그룹부터 97%, 97%, 94%, 100%로 약한 생장억제를 보였다. 4그룹에서는 FIn과 FCh 모두 생장이 억제되었지만 FIn 이 유의하게 더 억제되었다. FIn는 FCh 보다 초반에 상대적으로 강한 영양생장을, 후반에 생식생장을 한 것으로 분석된다. 상대적 절간 비율은 100% 이하면 생장이 억제되는 생식생장을 나타내고 100% 이상이면 생장이 왕성한 영양생장으로 판단한다(Jang et al., 2016).
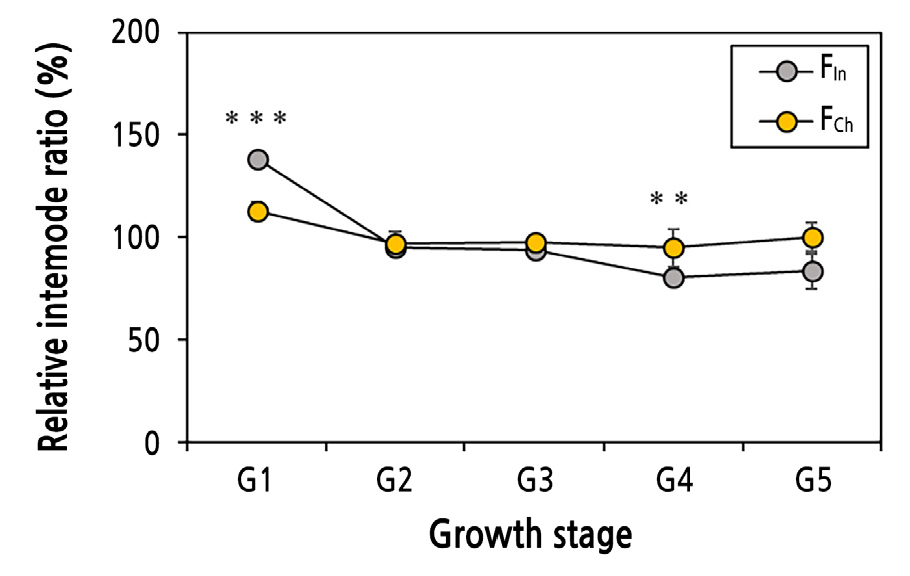
Fig. 4.
Changes in the relative internode ratio (average internode length/total average internode length x100) of FIn and FCh according to the growth stage. G1 emerged from March to early May, G2 from mid-May to early June, G3 from mid-June to July, G4 from August to September, and G5 from October. Bars indicate the standard errors of the means. ** and *** denote significance at p < 0.01 and 0.001, respectively (n = 6).
전 재배 기간 FIn와 FCh의 단위 면적당 생산량은 각 23.8kg·m-2, 15.3kg·m-2였다(Fig. 5). FIn와 FCh의 총생산량 차이는 품종, 재배 기간과 기술의 영향을 배제할 수 없지만, 개화 후 1–5주에 FIn의 일평균온도가 FCh보다 높아 상대적으로 영양생장이 유도되어 초기에 축적한 광합성 산물이 이후 생산량에 긍정적인 영향을 미친 것으로 추측된다. FIn와 FCh의 하반기 생산량은 각 7.6kg·m-2, 6.1kg ·m-2로 전체의 각 36.1%, 39.9%이었다(Fig. 5). 여름 작형에서 하반기(8–9월) 수량이 급격히 감소하는 경향은 Um et al.(2013)의 보고에서 여름 작형 파프리카 생산량이 8월 460kg·ha-1·day-1 로 최저였다는 결과와 유사하다. 이는 상·하반기의 광과 온도 환경이 달랐기 때문으로 상반기의 광량을 100%로 했을 때 하반기는 60–70% 수준으로 상대적으로 약광인데 반해 7월 한 달간 매일 고온으로 유지된 온실 내부로 인하여 낙화와 낙과를 유발하였을 것으로 생각된다. 광량과 생산량의 정의 관계는 이미 보고되어(Myung et al., 2012) 광량이 감소할수록 개화 개수와 과실 개수가 적어진다고 하였다(Jaafar, 1995; Jeeatid et al., 2017).

Fig. 5.
Total production in kilograms per square meter for FIn and FCh divided into two halves: the first half from May to July and the second half from August to October. The cultivation durations of FIn and FCh are 35 and 31 weeks, respectively. The production volume was measured every two weeks from the sample plants.
Fig. 6은 그룹별 착과율 결과이다. FIn의 그룹별 착과율은 1그룹부터 75.7, 61.8, 70.8, 68.8, 26.4%로 평균 60.7%의 착과율을 보였다. 그룹별 착과율의 차이는 착과기와 과실비대기(영양생장기)를 반복하는 파프리카 생육 특성에 의한 것으로 생각된다. 그러나 5그룹은 예외적으로 낮은 착과율을 보였다. FCh의 그룹별 착과율은 1그룹부터 25.7, 26.4, 17.4, 22.9, 5.6%로 FIn의 절반 수준의 착과율을 보였으나 FIn와 동일하게 5그룹의 착과율이 급격히 낮아졌다. 이는 재배 마무리 시기로 식물체의 수세가 약해진 영향과 하반기 저일조와 높은 주간/야간 온도 때문으로 추측된다. Bakker(1989)는 24시간 평균 온도가 높아질수록 착과 수가 적어진다고 보고하였고, Marcelis et al.(2004)는 개화 후 2주간 4일의 33°C의 고온처리가 개화 후 1주에 민감하게 반응하여 최고 75%의 낙화를 유도한다고 보고하였다. Rylski와 Spigelman(1982)는 파프리카 재배 시 야간 온도 18°C에서는 착화를 유도하지만 24°C에서 낙화 발생을 보고하였다. 본 실험에서는 낙화율을 조사하지 않아 상관관계를 분석하지는 못했지만 FCh의 야간 온도가 매일 24°C를 웃돌았던 7월 중 발생했던 3그룹의 착과율이 17.4%로 그 전후 26.4%(2그룹)와 22.9%(4그룹) 대비 약 65–75% 수준으로 낮은 것으로 미루어보아 야간 온도의 영향을 받은 것으로 추측된다. FCh의 착과율이 FIn의 절반 수준으로 낮았던 이유에 환경요인 외 다양한 요인이 있겠으나 높은 야간 온도(>21°C)는 파프리카 꽃눈분화 후 수정 불량을 일으키므로(RDA, 2020) FCh의 야간 온도가 FIn보다 높게 관리된 날수가 더 많고 평균적으로 높게 관리된 것도 영향을 미쳤을 것으로 생각된다. Bakker(1989)는 주야간 온도가 동시에 높아지면 낙과율이 증가하는데, 주간 온도보다는 야간 온도에 더욱 명확하게 반응하였다고 하였다.

착과 대비 수확률은 FIn와 FCh가 3그룹까지 81–91%의 높은 수확률을 보였다. 그러나 3그룹에는 FIn와 FCh가 각 61.4, 70.1%로 떨어졌고 5그룹에서는 6.1, 0%까지 떨어졌다. 이는 FIn의 경우 8–9월에 착과한 과실 중 약 40%가, FCh의 경우 약 30%가 낙과하거나 비상품과였다는 것을 의미한다. 따라서 하반기 저조한 생산량은 4–5그룹의 낮은 착과율과 수확률 때문으로 분석된다. 낙과는 일조량 감소(source strength) 또는 식물체 기관 간 영양분 경합(sink strength) 때문에 발생하며(Marcelis et al., 2004) 특히 개화 후 14일 이내의 어린 과실이 온도에 민감하다고 알려졌다(Wubs et al., 2009).
환경요인과 생산량 간 상관관계 분석
독립변수인 주간 온도와 야간 온도, 생육도일(Growing degree days, GDD)과 일사량이 종속변수인 생산량과 갖는 관계성 분석을 다중회귀모형으로 확인한 결과 다음과 같은 다중 선형회귀모델이 도출되었다.
y생산량 = 10792.279 +8.490x1주간온도 – 20.513x야간온도 – 19.962xGDD + 2.578x일사량
주간 온도와 일사량은 생산량과 양의 관계를 보였고, 야간 온도와 생육도일(일평균 온도 ‒10)은 음의 관계를 보였다. 이를 통하여 주간 온도와 일사량이 증가하면 생산량도 증가하고, 야간 온도와 생육도일이 증가하면 생산량은 감소하는 것을 알 수 있다. 각 환경요인의 영향력을 나타내는 표준화된 계수(Standardized coefficients)는 주간 온도와 야간 온도, 생육도일, 일사량이 각 0.405, ‒0.837, ‒0.505, 0.657로 나타나, 영향력은 야간 온도 > 일사량 > 생육도일(일평균 온도 ‒10) > 주간 온도 순서로 높았다(Table 4). 즉, 일평균 일사량이 1MJ·m-2 증가하면 생산량은 약 0.7g 많아지고, 주간 온도가 1°C 상승하면 생산량은 0.4g 많아지는 반면, 야간 온도가 1°C 상승하면 생산량은 0.8g 감소하고, 일평균 온도가 1°C 상승하면 생산량은 0.5g 감소함을 의미한다. 이러한 결과는 다른 연구 결과에서도 보고되었다. Um et al.(2013)은 여름 작형의 8월 수량 증대를 위하여 야간 온도를 낮게 관리하는 것을 제안하고 있다. 야간 온도는 착과와 수확량에 영향을 미치는 요인으로 야간 온도 24°C에서 식물체 당 6.6개의 과실을 수확했지만, 야간 온도 18°C에서는 12.6개를 수확했다고 보고되었고(Rylski and Spigelman, 1982), 상대적으로 높은 야간 온도에서는 품종과 관계없이 착과량이 적다고 보고되었다(Saha et al., 2010). 도출된 모델은 수정된 결정계수(adjusted R2) 0.679의 설명력을 가졌으며 유의성이 0.001보다 작아 통계적으로 유의미한 모형임을 나타냈다(Table 5).
Table 4.
Results of the impact of environmental factors on production of FIn and FCh
Table 5.
Summary on the fitness of the multiple regression model
| R | R2 | Adjusted R2 |
Std. error of the estimate | ANOVAy | ||
| df | F | Significance | ||||
| 0.864z | 0.746 | 0.679 | 1214.194 | 4 | 11.028 | 0.000 |
따라서 온도가 상승하는 고온기에는 광량을 최대로 확보하면서 야간 온도를 낮게 관리하여 일평균 온도가 21°C로 균일할 수 있도록 유지하는 것이 중요할 것으로 생각된다. 변화폭이 큰 온도관리는 동화산물 분배와 source-sink 비율을 변화시켜 생육의 불균일과 과실 수확에 영향을 미치기 때문에 균일한 생육과 생산량을 유지하기 위해서 온실 내부 주/야간 온도를 균일하게 유지할 필요가 있다고 하였다(Marcelis et al., 2004; Jeong et al., 2009). 네덜란드도 생산량 저조의 원인인 낙화와 낙과, 착과 불량과 비상품과율 예방을 위하여 시설 내부 하루평균 온도를 21°C, 야간 18.8°C로 주간 변동 없이 비교적 균일하게 관리하여 안정된 생산을 하고 있고(Jeong et al., 2009), 추가로 (i) 작형 변경, (ii) 과실 가지치기, (iii) 단위결실 과실 생장을 유도하는 방식(Heuvelink et al., 2002)을 사용하여 주간 안정생산을 도모하고 있다. 현재 강원도 파프리카 여름 재배의 관행으로 스크린을 통한 주간 온도 제어가 이루어지고 있다. 관행의 스크린 방식은 광량을 제한하는 한계점이 있으나(Woo et al., 2014) 보온이 어려운 온실 내부 냉방비를 고려하였을 때 경제적이고 반영구적인 스크린 방식이 여름철 온실 내부 온도관리에 효과적이다(Kim et al., 2021). 상업적 온실에서 고온기 파프리카 생산량 증대를 위하여 스크린을 통한 온도 제어 시 광량을 최소한으로 감소시키면서 주간 온도를 제어하고 일몰 시 빠른 온도하강을 유도할 수 있는 스크린 사용과 냉각팬 등 적극적인 장치 사용이 요구된다. 파프리카 재배의 총 재배 기간은 31–37주인 것을 고려하였을 때 15–24주는 파프리카 재배 하반기에 들어서는 시기이자 약광과 고온 환경이 조성되는 시기이므로 해당 시기에 실용적이고 효과적인 환경관리개선이 추후 연구되어야 할 것으로 생각된다.
