

Introduction
Materials & Methods
Plant materials and growing conditions
Temperature and light treatments
Data collection and analysis
Results and Discussion
Plant growth and flowering
Relationships of crop characteristics with ADT and DLI
Introduction
Eustoma grandiflorum (Raf.) Shinn. (eustoma or lisianthus) originates in the southern part of the United States (Dole and Wilkins, 2005; Halevy and Kofranek, 1984; Kawakatsu and Fukuta, 2012). This plant initially forms a rosette after seed germination, grows very slowly during winter, elongates its stems in spring, and then flowers in summer (Dole and Wilkins, 2005; Roh et al., 1989). Eustoma is a seed- propagated ornamental plant grown as cut flowers and potted plants worldwide. Selected cultivars and environ-mentally controlled greenhouses have enabled year-round production (Islam et al., 2005).
Eustoma is generally classified as a quantitative long-day plant (LDP) (Dole and Wilkins, 2005; Grueber et al., 1984; Roh et al., 1989; Thomas and Vince-Prue, 1997; Tsukada et al., 1982; Zaccai and Edri, 2002), but some researchers (Azrak, 1984; Halevy and Kofranek, 1984; Harbaugh, 1995; Islam et al., 2005) consider a number of cultivars to be day- neutral or obligate LD plants due to their complex responses to photoperiod. Harbaugh et al. (1997) reported that photoperiodic response of Eustoma was cultivar-dependent as most cultivars are facultative LDPs, but a few are almost day-neutral or obligate LD plants. Time to flowering of eustoma has also been shown to be reduced by high light intensity (Corr and Katz, 1997; Islam et al., 2005) and high temperature (Islam et al., 2005; Zaccai and Edri, 2002).
However, these previous studies have not sufficiently explained the independent or interactive effects of tem-perature, photoperiod, and light intensity because they used two parameters with two levels. In addition, the effects of these environmental conditions on growth and flowering of eustoma are affected by developmental stage (Zaccai and Edri, 2002). At the seedling stage (up to 4-5 true leaf pairs), higher temperatures induce rosette formation and delay bolting (Ohkawa et al., 1994) depending on cultivars (Harbaugh and Scott, 1999). At the later stage, stem elongation and flower bud initiation and development can be hastened by higher temperature, higher light intensity, and long days (LD) (Halevy and Kofranek, 1984; Harbaugh, 1995; Roh and Lawson, 1984).
For year round production of good-quality eustoma, it is necessary to understand the growth and flowering responses to combined conditions of three environmental factors including temperature, photoperiod, and light intensity. Therefore, this study was conducted to examine the effects of temperature, photoperiod, and light intensity during the post-seedling stage on growth and flowering in E. grandiflorum.
Materials & Methods
Plant materials and growing conditions
E. grandiflorum ‘El Paso Deep Blue’ seedlings were received from a commercial propagator (ACC KA Seed & Seedling Co., Ltd., Jincheon, Korea), transplanted to plastic pots (7 cm diameter) filled with Sunshine Mix #1 (Sun-Gro Horti-culture, Bellevue, WA, USA) on April 10, and grown in a greenhouse at 22 ± 2°C until the beginning of treatments. The potted plants were watered using a stagnant wick subirrigation system with nutrient solutions of 150 mg・L-1 N from 20N-3.7P-16.6K (Technigro 20-9-20 Plus, Sun-Gro Horticulture) and 15N-0P-12.5K fertilizer (Technigro 15-0-15 Plus, Sun-Gro Horticulture) in alternation.
Temperature and light treatments
Plants with six expanded leaves were moved to growth modules in walk-in chambers. Three environmentally controlled chambers were maintained at 14 ± 2°C (LT), 20 ± 2°C (MT), and 26 ± 2°C (HT), respectively. Each chamber was divided into two sections that generated 8 h (short day, SD, 08:00-16:00) and 16 h (long day, LD, 08:00-24:00) photoperiods using fluorescent lamps (FL; FL20SEX-D, Kumho Electric Inc., Seoul, Korea) and incandescent lamps (INC; IL220V30W-L, Kumho Electric Inc., Seoul, Korea). Three modules in each section provided three photosynthetic photon fluxes [PPF, 100 ± 5 (LL), 200 ± 10 (ML), and 400 ± 20 (HL) μmol・m-2・s-1] at canopy height. The FL:INC ratio in PPFD at the plant canopy was about 8:2; therefore, plants were exposed to four levels of daily light integral [DLI, DLI (mol・m-2・d-1) = PPF (μmol・m-2・s-1) × photoperiod (h・d-1) × 60 (min・h-1) × 60 (s・min-1) × 10-6], about 3 (LL × SD), 6 (LL × LD and ML × SD), 12 (ML × LD and HL × SD), and 24 (HL × LD) mol・m-2・d-1. Twenty plants were grown in each module under different light and temperature conditions (total 18 treatments) for 3 weeks.
Data collection and analysis
Morphological characteristics such as plant height, numbers of nodes and branches, visible flower buds, open flowers and shoot dry weight (DW) were evaluated at six weeks after the start of treatments based on ten plants per treat-ment. Leaf canopy height was measured from the surface of the growth medium to the top of the shoot and DW was measured after drying shoots for three days at 80°C. The remaining ten plants were grown continuously until the 10th week after treatment and the date of VB (when the first flower bud was 2 mm in size) and flowering, numbers of nodes below the first flower bud and cumulative flower buds were recorded.
Data were analyzed by SAS (SAS Institute Inc., Cary, NC, USA) using analysis of variance (ANOVA) and the general linear model (GLM) procedures. Regression analysis was conducted using SigmaPlot (SigmaPlot v10, Systat Software Inc., Chicago, IL, USA). A p < 0.05 was considered to indicate significance.
Results and Discussion
Plant growth and flowering
Temperature (ADT), photoperiod, and light intensity solely and interactively influenced the growth characteristics of E. grandiflorum ‘El Paso Deep Blue’ (Table 1). After tem-perature and light treatments for three weeks and subsequent growth for three weeks in a greenhouse, plant height increased as ADT increased from 14°C to 26°C. Light conditions had a smaller impact on plant height than temperature in this study. However, LD promoted stem elongation relative to SD under MT+LL, and HT+HL inhibited stem elongation regardless of photoperiod. High temperature, long photoperiod, and low PPFD promote the biosynthesis of active gibberellic acids such as GA1 and GA4 in cyclamen (Oh, 2007). Additionally, longer photoperiod induced transiently elevated levels of GAs (Olsen et al., 1997). Silene armeria under LD had higher levels of GA1, whereas the content was lower under SD (Talon and Zeevaart, 1990). Stem elongation of spinach, a long day plant (LDP), under LD (16 h) was inhibited by theobroxide owing to decreased endogenous GA1 (Kong et al., 2006). Thus, elongated stems of eustoma plants grown under high temperature, long photoperiod and low PPFD in this study were assumed to result from an increase in active GAs for stem elongation.
Leaf number and shoot dry weight increased with increasing temperature, photoperiod, and light intensity (Table 1). Moreover, the promoting effects of PPFD on dry mass accumulation became greater as temperature increased from 14°C to 26°C because the difference in dry weight among LL, ML and HL grew with increasing temperature. Conversely, temperature had little effect on the increase in dry weight in response to LD because there was little difference in dry weight between SD and LD, regardless of temperature change. Dramatic increases in leaf number under HL resulted from branching and unfolding of small leaves on lateral shoots. Temperature and light conditions affect photosynthetic and leaf unfolding rates in all plants (Dole and Wilkins. 2005). For example, higher irradiance promoted growth characteristics by increasing photosynthetic rate in wax begonia (Nemali and van Iersel, 2004). Additionally, Cyclamen persicum showed increased photosynthetic rate, shoot dry weight and leaf number in response to higher daily light integral (Oh et al., 2009), while higher temperature increased plant volume, leaf number and plant dry weight at flowering (Karlsson and Werner, 2001a, 2001b). In the pansy (Viola × wittrockiana Gams.) ‘Universal Violet’, increasing light integral and temperature up to an optimum of 25 ± 3°C increased dry weight, and the rate of leaf production on the main stem increased linearly with both light integral and temperature (Adams et al., 1997a). High light intensity also increased shoot dry weight and stem diameter relative to low light in E. grandiflorum ‘Echo Blue’ and ‘Fuji Deep Blue’ (Islam et al., 2005). In addition, supplemental lighting during the seedling stage increased the number of leaves in petunia and pansy (Oh et al., 2010).
The number and length of branches showed similar trends to leaf number and shoot dry weight in response to tem-perature and light conditions (Table 1). LT at 14°C under all light treatments did not induce branching, while MT under LL and ML inhibited lateral shoot growth (Table 1). All light conditions under HT induced the development of axillary buds except for SD + LL. At HT, the number and length of branches increased with photoperiod and PPFD. Low temperatures decreased the growth and developmental rates of processes such as branching and leaf unfolding in cyclamen (Karlsson and Werner, 2001b), and low irradiance intensified apical dominance as a phenomenon of shade avoidance syndrome induced by low irradiance under light with low red to far-red ratio (Franklin and Whitelam, 2005; Huber and Stuefer, 1997; Moe and Heins, 1990).
All flowering characteristics of E. grandiflorum were influenced by temperature and light conditions in this study (Table 1 and 2). After treatment for three weeks followed by growth in the greenhouse for three weeks, the number of flowers increased with increasing temperature, photoperiod, and PPFD; however, the eustoma plant had no flower under LT (Table 1). Days to visible bud and first flowering decreased with increasing temperature, photoperiod, and light intensity (Table 2). The number of nodes under the first flower was slightly reduced by higher temperature, longer photoperiod, and higher PPFD. The number of open flowers and visible buds counted until 10 weeks after treatment increased somewhat with increasing temperature, photoperiod and PPFD. Plants grown in the MT and HT chambers had more flower buds than those grown in the LT chamber, and the number of flower buds increased proportionally to PPFD. Islam et al. (2005) reported that providing a DLI of 8.8 mol・m-2・d-1 for 16 h (LD) decreased days to visible budding and flowering compared with that of 4.4 mol・m-2・d-1 DLI for 16 h in E. grandiflorum ‘Echo Blue’ and ‘Fuji Deep Blue’. In locations with lower light intensities, LD will enhance bolting, and high intensity discharge (HID) lighting such as high pressure sodium lamps will improve plant quality (Dole and Wilkins, 2005).
Temperature, photoperiod, and light intensity generally affect flowering of plants. Flowering is promoted by higher temperatures in cyclamen (Karlsson and Werner, 2001a, 2001b; Oh et al., 2013) and pansy (Adams et al., 1997b). High PPFD also hastens flowering in some floriculture crops, with facultative irradiance response (Oh et al., 2009). Photoperiod affects flowering of photoperiod-sensitive plants such as LDPs and SDPs. Flowering of E. grandiflorum generally occurred quicker or days to flowering decreased under LDs (Islam et al., 2005; Zaccai and Edri, 2002). However, the influence of photoperiod on days to flowering was relatively small compared with temperature and light intensity in this study. This was likely because the effect of the range of photoperiods from 8 to 16 h used in this study was smaller than that of temperatures from 14°C to 26°C and PPFs from 100 to 400 μmol・m-2・s-1 on flowering of this cultivar. In lisianthus, temperatures below 16°C reduced growth rate and increased production time (Harbaugh et al., 1997); however, temperature for bolting and flowering must be maintained at above 18/16°C (day/night) in a greenhouse (Harbaugh, 1995; Roh and Lawson, 1984; Roh et al., 1989). Therefore, the ADT range of this study included the limiting temperature of 14°C. Conversely, the cultivar of ‘El Paso Deep Blue’ could be considered an almost day- neutral plant because photoperiod had very little effect on flowering.
Relationships of crop characteristics with ADT and DLI
There were significant interactions among the effects of some environmental conditions on growth and flowering characteristics (Table 1 and 2). However, we carried out regression analysis using ADT and DLI derived from PPF and photoperiod because the effects of light conditions, especially photoperiod, were smaller than those of tempera-ture. After PPF and photoperiod of Table 1 were transferred into daily light integral (DLI), several characteristics had significant correlations with ADT and DLI based on regression analysis (Table 2). For example, as ADT and DLI became higher, the number of leaves (y = -29.30 + 2.10ADT + 1.46DLI, R2=0.83), number of flowers (y = -8.05 + 0.42ADT + 0.17DLI, R2 = 0.71), lateral shoot length (y = -7.87 + 0.40ADT + 0.19DLI, R2 = 0.69), and shoot dry weight (Y = -0.88 + 0.10ADT + 0.06DLI, R2 = 0.86) increased (Fig. 1A, B, C, D).
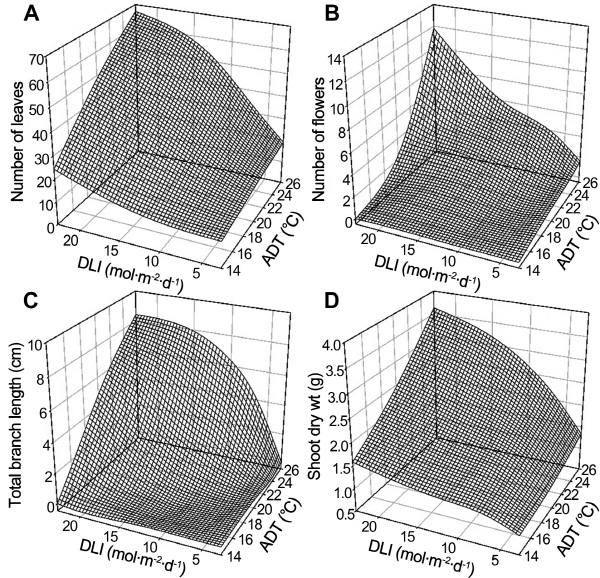
The number of leaves increased in proportion to DLI and ADT (Fig. 1A). In addition, the impact of temperature became larger with increasing DLI, and the effect of DLI was larger as temperature increased. Low ADT and low DLI inhibited lateral shoot elongation, whereas lateral shoot length increased proportionally to ADT under high DLI (Fig. 1C). Lateral shoot length increased sigmoidally under high ADT, with the maximum value occurring at 15 mol・m-2・d-1 DLI. Shoot dry weight showed a greater increase under higher ADT and DLI (Fig. 1D). DLI had little effect on the number of flowers under LT but a large impact under HT (Fig. 1B).
Overall, an increase of about 1 mol・m-2・d-1 DLI could constitute an increase of 0.40°C to 0.76°C ADT depending on crop characteristics when the ADT and DLI are above 20°C and 12 mol・m-2・d-1, respectively, within the environ-mental range investigated in this study. Therefore, growers can select one regimen of heating or supplemental lighting based on cost without delaying harvest time or reducing crop quality.
