

Introduction
Materials and Methods
Plant Materials
Chromosomal Study
Data Analysis
Results
Chromosome Counting
Color Differences in Cotyledons
Frequencies of Different Cotyledon Types
Morphological Characteristics
Seedling Characters
Plant Characters
Discussion
Conclusion
Introduction
The stock flower, Matthiola incana, belongs to Brassicaceae, has a large genetic variation, and can be grown in different regions. This Mediterranean flower is the subject of many studies around the world (Celikel and Reid, 2002; Heuer and Ravina, 2004; Tatsuzawa et al., 2012). Also, this commercial cut flower is one of the best options for greenhouse owners during the winter due to its high resistance against cold (Eid et al., 2009). Flower doubleness seems to be the main feature for breeding research in the ornamental species such as stocks, roses (Dubois et al., 2010), Catharanthus (Chen et al., 2012), and carnations (Yagi et al., 2014). Two different forms of stock flower have been characterized: single and double-flowered. The second form is completely sterile, lacking reproductive organs (Philp and Huskins, 1931; Roeder and Yanofsky, 2001) and thus, double-flowered stock has to be produced from seeds of single-flowered plants (Lesley and Frost, 1927). On the other hand, double flowered plants are more valuable for their beauty and marketability, as it is used as a cut flower (Winge, 1931). Since the proportion of double to single-flower progenies is of financial importance for this plant (Johnson, 1953), M. incana breeders have attempted to raise this ratio by different methods, including finding high doubleflower cultivars (Frost and Lesley, 1954) and the identification of genetic markers.
Kappert (1937) discovered a linkage between the color of cotyledons with the gene that controls the single and doubleflowered state. Based on this research, the homozygous allele of this gene induces pale green cotyledons. If all seedlings with dark green cotyledon are removed, it may lead to 100% double-flowered plants. Although the difference between pale green cotyledons and dark ones is sometimes not clear and cannot be used for all cultivars, this method is the principle selective process for stock flowers and is used in industry at the present time. Also, Ecker et al. (1993) indicated a strong linkage between the wavy leaf gene and the double flowering state; however, the wavy form leaves cannot be identified at the seedling stage. There are some investigations of the relationship between the cytogenetic features and proportion of double flowers (Frost, 1927; Frost and Lesley, 1954; Prakken, 1942). A high percentage of double flowers (90%) were discovered in aneuploid stocks and determined to be “high double” by Frost et al. (1959). Based on this study, this large percentage of double flowers is due to the existence of chromosome fragments that emerged from chromosome breakage.
Most cytogenetic studies on the stock plant date back to over half a century. There is a wide variety of stock cultivars in terms of morphological features and the ratio of double-to-single flowers (Tatsuzawa et al., 2012). According to the study of Emsweller et al. (1937): “There is need for the discovery of some simple seedling characteristic to enable florists and gardeners to select with certainty double-flowered plants in the seedling stage”. Therefore, finding seedling characteristics to distinguish double flower plants at early growth stages was the aim of our research. In stock cultivars, we observed different morphological types in cotyledon of seedlings, which varied in shape, size, and number. The main aim of the present study was the identification and investigation of these phenotypes in cotyledons. For this purpose, in this study we endeavored to clarify the relationship between cotyledon shape and the double flowering state in three types of stock cultivars (Emsweller et al., 1937): type-1 never creates any double flowers (Singles), type-2 creates 40-50% double flowers (Eversportings), and type-3 creates more than 80% double flowers (High doubles). In recent years, “Eversporting” cultivars are very common and cost effective for farmers, and because of their low rate of double- flowering trait the farmers tend to purchase high double cultivars. In addition, we tried to compare the different forms of cotyledons and their morphological and cytogenetic characteristics among different cultivars of stock flowers.
Materials and Methods
Plant Materials
In this study, eight stock cultivars were tested for cotyledon shape (Table 1). The seeds were obtained from seed companies and public breeding institutions, and planted in seedling trays in a research greenhouse. At the four-leaf stage, seedlings were transferred to 6-cm-diameter pots containing a mixture of coco peat and perlite. The seedlings were counted and labeled based on the shape of the cotyledons (dumbbell, heart and cup-shaped) at the two-leaf stage. Also, the seedlings with dumbbell-shaped cotyledons (DC) were exposed to cold temperatures below 8°C with light and divided based on the color of the cotyledons (pale and dark green). Plants were transferred to larger containers (mixture of soil, peat moss, perlite, and coarse sand), in greenhouse conditions, and maintained at 8-18ºC for better growth. In order to study morphological characteristics at two growth stages (seedlings and adult plants), a completely randomized design with two factors (cultivar and cotyledon type) was performed. Nine morphological characteristics were evaluated at the seedling and mature plant stages. All measurements relating to seedlings were conducted at the two-leaf stage, and in the mature plants measurement occurred after the full emergence of inflorescent.
Chromosomal Study
In order to study chromosome number, anthers of young floral buds were taken from each cultivar and fixed in a Carnoy's solution for 24 h (Ranjbar and Karami, 2014). The materials were washed two times in sterile water and stored in 70% ethanol. In this study, we used 2% aceto-orcein for staining. Finally, 30 diakinesis cells from each cultivar were imaged using an Olympus CX21 microscope. The chromosome number at mitotic metaphase was evaluated with the application of squash preparation and aceto-orcein stain. The apical meristem (1-1.2 cm long root tip) was used for cytogenetic analysis of somatic chromosomes. On average, five well-spread mitotic metaphase chromosomes per plant for each cultivar were analyzed cytologically.
Data Analysis
Analysis of variance (ANOVA) for the comparison of means (±SE) of morphological characteristics was used. Chi square analysis was used to test whether the difference between observed ratios of double-to-single flowers and hypothesized if a 1:1 ratio would be statistically significant or not. Pearson’s correlation coefficients were computed to check the relationship between chromosome number and cotyledon traits. Finally, data analysis was conducted using SPSS ver.16.0 software.
Results
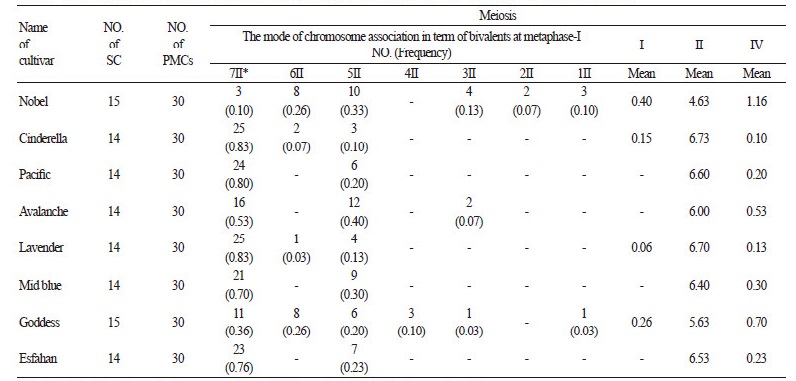
Chromosome Counting
For the cytogenetic study, 30 pollen mother cells at the diakinesis stage from each cultivar were studied. The number of chromosomes in gametic cells of the stock flower was calculated as (n = x= 7). It was expected that bivalent chromosomes were present at the diakinesis stage of meiosis I, but quadrivalent chromosomes were also observed in the studied cultivars. The results indicated that ‘Nobel’ and ‘Goddess’ cultivars have abnormal chromosome behavior and additional chromosomes are present in some of their diakinesis cells. The observed aneuploidy cells in ‘Nobel’ and ‘Goddess’ cultivars were confirmed by investigating somatic cells (2n+1=15). Diakinesis characteristics of eight cultivars are presented in Table 1. The minimum number of bivalents and the maximum number of quadrivalents were observed in the ‘Nobel’ cultivar. The highest number of bivalents and the least number of quadrivalents were recorded in the ‘Cinderella’ cultivar. Unpaired chromosomes (univalent) were detected in ‘Nobel’, ‘Goddess’, ‘Cinderella’, and ‘Lavender’ cultivars. High Frequency of aneuploidy in the pollen mother cells of ‘Nobel’ and‘ Goddess’ cultivars may be related to the abnormal chromosome behaviors during meiosis.
Color Differences in Cotyledons
Seedlings with dumbbell-shaped cotyledons (DC) were found in in six cultivars, which could be divided based on cotyledon color (light and dark green), as the difference between these two colors was visible in the two-leaf stage (Fig. 1A and 1B). The results indicated that the number of double flowers was equal to that of the individuals with pale green dumbbell-shaped cotyledons; furthermore, the number of single flowers is equal to that of the dark green dumbbell-shaped cotyledons (data not shown). However, in ‘Esfahan’ and ‘Nobel’ cultivars, no difference was found to have different colors in the DC seedling population. Therefore, the feature of color in DC seedlings was not the case for all stock flower cultivars. In the populations with both cotyledon forms (heart-shaped and cup-shaped), there was no difference in cotyledon color (lacks divisibility). Hence, after the emergence of inflorescence, the distribution of double-flowered and single-flowered individuals was determined.
Frequencies of Different Cotyledon Types
The frequencies of different cotyledon types in studied cultivars are shown in Fig. 2. The maximum and minimum numbers of DC (dumbbell-shaped cotyledons) were observed in ‘Esfahan’ (100%) and ‘Goddess’ (39.04%) cultivars, while the highest number of HC (heart-shaped cotyledons; 42.46%) was detected in ‘Goddess’ cultivar seedlings; the highest distribution of CC (cup-shaped cotyledons) was obtained from ‘Nobel’ cultivar, which also had the highest percentage of double-flowered plants. The least CC and HC numbers were observed in ‘Cinderella’, which had the least morphological characteristic values. Each shape of cotyledon showed a different percent of double and single-flowered plants. The results of a Chi-square test indicated that in both heart-shaped and cup-shaped cotyledon seedling populations, the ratio of double-to-single flowers was inconsistent with the hypothesis of a 1:1 ratio at the p < 0.05 level (Table 2). More importantly, the number of double flowers was considerably higher than single flowers in heart-shaped and cup-shaped cotyledon seedlings in the studied cultivars. In five cultivars, the CC seedlings formed only double flowers. A high percentage CC cotyledon had an increased overall percentage of double flowers in these cultivars.
Morphological Characteristics
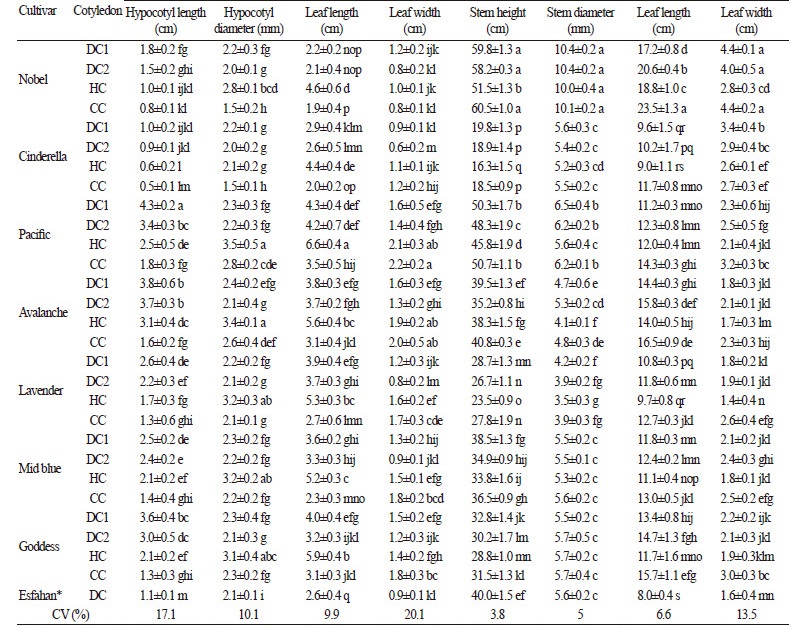
After the emergence of inflorescence, it was observed that there was a significant difference among the individuals in three different populations of HC, DC and CC plants, and a relationship between the characteristics of the seedlings and mature plants of each cotyledon phenotype. Moreover, a significant difference (p < 0.01) was also observed among the studied cultivars. The results obtained from morphological characteristic investigation in two growth stages were based on the shape of cotyledons in eight stock cultivars (Table 3). Considering the coefficient of variation (% CV) parameter, we observed high variation for both primary leaf widths in seedlings and the stem height of matured plants. Variance analysis indicated that the effects of cultivar and cotyledon shape had a significant difference in the 1% probability level in all studied characteristics.
Seedling Characters
In order to specifically comparing the cotyledon phenotypes between different cultivars, we performed slicing (the division of interactions) of morphological characteristics (data not reported). The color of cotyledons and the size of the stock seedling are two important traits that play a significant role in distinguishing double-flowered seedlings from the single-flowered ones (Emsweller et al., 1937). In seedlings with dumbbell-shaped cotyledons and dark or pale green colors, the two cultivars ‘Pacific’ and ‘Goddess’ showed significant differences in height (p < 0.0001). The shortest seedlings were detected in ‘Cinderella’, which has cup-shaped cotyledons (0.53 cm), while the tallest seedlings (4.33 cm) were observed in DC seedlings in ‘Pacific’ (Fig. 1). In general, the maximum and the minimum values for seedling height were observed in DC and CC cotyledons, respectively. Moreover, the results revealed that there was no significant difference between ‘Goddess’, ‘Nobel’, ‘Cinderella’, and ‘Lavender’cultivars with respect to the height of seedlings with cup-shaped cotyledons. In this test, the range of trunk diameter in seedlings varied from the least value in CC of ‘Cinderella’ cultivar (0.50 cm), to the greatest value in DC of ‘Pacific’(4.33 cm). The biggest dimensions of primary leaf (6.58 mm × 2.11 mm) were measured in HC seedlings from the ‘Pacific’ cultivar, and the smallest dimensions (2 mm × 0.80 mm) were measured in DC seedlings of the ‘Nobel’ cultivar. Lower values for morphological traits at the seedling stage were observed in heart-shaped cotyledons (HC).
Plant Characters
Based on the characteristics of mature plants, the difference in stem height between double-flowered and single-flowered plants was observed in ‘Pacific’, ‘Avalanche’, ‘Midblue’, and ‘Goddess’ cultivars. In these cultivars, the double-flowered plants had longer stems than the single-flowered ones, suggesting that this may be related to the delay in flower initiation in doubleflowered plants compared to the single-flowered ones in these cultivars (Mousavi et al., 2008). The highest values of stem height, width, and length in matured plant leaves were related to CC cotyledons, although this cotyledon form showed the lowest values for seedling height and primary leaf length in each cultivar. The important point here is that the highest inflorescence measurement was observed in CC seedlings, and that this value was the greatest in ‘Goddess’ compared to other cultivars (Fig. 3).
Fig. 3
(Right) A comparison of inflorescence size among four cotyledon phenotypes in eight Matthiola incana L. cultivars. (Left) The difference between the length of the inflorescence with double flowers in ‘Pacific’ cultivar of Matthiola incana. The dark green dumbbell-shaped cotyledon did not produce doubles, so (B) is not in left figure.
Discussion
The aim of the present study was to focus on the shapes of cotyledons in Matthiola incana L. cultivars. In addition, cytogenetic analysis was performed during diakinesis to investigate the relationship between the number of chromosomes and the percentage of cotyledon forms. All diakinesis cells in ‘Cinderella’, ‘Pacific’, ‘Avalanche’, ‘Lavender’, ‘Midblue’ and ‘Esfahan’ cultivars possessed n=x=7 chromosomes (Allen, 1924; 2n=2x=14), but ‘Nobel’ and ‘Goddess’ cultivars had extra chromosomes in several gametic and somatic cells (2n+1). Frost et al. (1959) stated that the origin of these extra chromosomes was the breakage of chromosomes during anaphase I in aneuploid stock plants. Frost and Lesley (1954) described aneuploidy in stocks with high instance of double flower progenies. They found that the extra chromosomes resulted from the increase in the percentage of double plants. ‘Nobel’ and ‘Goddess’ cultivars showed extra chromosomes in gametic cells. The percentage of double-flowered plants in ‘Nobel’ and ‘Goddess’ cultivars was 86.21% and 80.14%, respectively. Since the presence of quadrivalent chromosomes may produce some breakage and closures in the first anaphase of meiosis, which eventually lead to an imbalance in chromosomal segregation and production of pollen that transmit altered chromosomes to the next generation (Veilleux, 1985). It is obvious that disorders such as univalent and quadrivalent chromosome formation may cause the elimination or lack of some characteristics in future generations (Morrison and Rajhathy, 1960; Ramsey and Schemske, 2002). The lowest and highest numbers of bivalent and quadrivalent chromosomes were both observed in the ‘Nobel’ cultivar. This finding is consistent with the results obtained by previous work with aneuploid stock flower plants (Frost, 1931).
Different cotyledon phenotypes are observed in the Brassicaceae family and have been investigated genetically in Arabidopsis (Chandler, 2008). Three genes, namely CUC1, CUC2, and CUC3 from the cup-shaped cotyledon (CUC) gene group that are under cotyledon boundary control were identified as the responsible genetic factors for distinguishing two cotyledons in Arabidopsis (Aida et al., 1997). Based on the study conducted by Aida et al., 1997 mutations in these genes led to the formation of cup- or heart-shaped cotyledon during seedling development. Additionally, according to this study, some changes in the structure of the flower were observed that were present in seedlings with cup-shaped cotyledons. While in the present study no structural change was observed in the stock flowers of CC and HC seedlings, a number of changes were noted in some morphological characteristics in plants with cup-shaped cotyledons compared to those with dumbbell-shaped cotyledons. The results showed that the small seedlings with dark green dumbbell-shaped cotyledons (DC2) and the seedlings with heart-shaped cotyledons (HC) produced a higher number of single flowers; however, the larger seedlings with pale green dumbbell-shaped cotyledons (DC1) and the seedlings with cup-shaped cotyledons (CC) produced more double flowers. This finding is consistent with the study conducted by Saunders (1923) on the size of the stock seedlings.
Based on a previous study (Kappert, 1937), it was assumed that approximately equal numbers of double and single flowers were observed in seedlings with dumbbell-shaped cotyledons, while this hypothesis was rejected in ‘Nobel’, ‘Goddess’, and ‘Pacific’ cultivars. In seedlings having dumbbell-shaped cotyledons that were dark and pale green in color in the two cultivars ‘Pacific’ and ‘Goddess’, significant differences were observed in the plant height (p < 0.0001), consistent with Emsweller’s findings (1937). He found that double-flowered seedlings with pale green dumbbell-shaped cotyledons are more robust in hypocotyl length and diameter than the single-flowered plants. Thus, distinguishing the double-flowered plants in these cultivars, compared to other cultivars, is easier at the seedling stage, supporting the hypothesis that there is a relationship between the aneuploidy of the cultivar and the high number of CC seedling population requires genetic studies. Investigation of the relationship between these characteristics demonstrated that there was a significant negative correlation between the percentage of double flowering and the number of seedlings with dumbbell-shaped cotyledons (p <0.05). This may be due to the high percentage of single-flowered plants in the population of this cotyledon shape, which may lead to the reduction in the total percentage of the double-flowered trait. On the other hand, a significant negative correlation (p < 0.01) was noticed between HCand DC populations. Moreover, the results demonstrated that the strongest positive correlation existed between the mean number of quadrivalent chromosomes and the frequency of CC seedlings; however, a highly negative correlation was found between the number of bivalent chromosomes and the frequency of CC seedlings (Table 4).
Table 4. Pearson correlation analysis of six studied features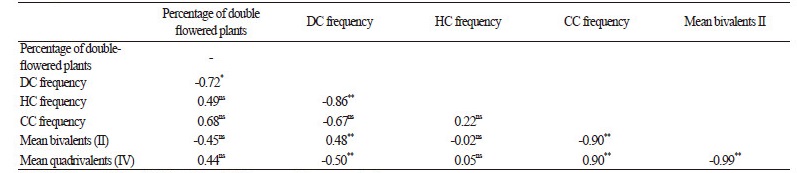 |
*, **Significant at 5 % and 1 % level of probability, respectively. ns: not significant |
However, in the present study, besides aneuploidy as a significant factor in high flower doubling (Frost and Lesley, 1954), we suggest that one of the reasons for the high flower doubling in these cultivars could be predicted by the type of cotyledon. This positive correlation between flower number and cotyledon physiology could be attributed to the ‘Nobel’ and ‘Goddess’ cultivars with the highest number of quadrivalent chromosomes that had the highest number of cup cotyledons.
Conclusion
The population of cup cotyledons is much smaller than that of other cotyledon forms. Furthermore, we cannot limit our research to this form of cotyledon and overlook other forms; however, in the two stock plant populations of seedlings with heartshaped and cup-shaped cotyledons, a large number of plants are double-flowered. It can also be shown that almost all seedlings with cup-shaped cotyledons produced double-flower plants in these cultivars. Therefore, it is required to investigate the frequencies of various cotyledon shapes in a large number of stock cultivars. The aneuploidy is an important phenomenon because a high number of cup-shaped cotyledons were observed in the ‘Nobel’ cultivar, which is highly double-flowered. The high rate of double-flowers in this cultivar is important with regard to the presence of additional chromosomes and the high number of cup-shaped cotyledons. Any morphological markers linked with the double-flowering trait are useful tools for early selection of stock breeding programs. This selection can be helpful in reducing the high costs that result from the long period of storage of plants, in addition to the predetermined markers.
