서 언
재료 및 방법
브로콜리의 생육 및 시험구 처리
식물 전기적 신호(PIES) 모니터링
잎의 엽록소, 프롤린 함량 분석
줄기 추출액의 이온 함량 분석
브로콜리 토양의 수용성 및 교환성 원소 함량 분석
데이터 통계처리
결과 및 고찰
토양 요소 처리와 브로콜리 전기적 신호의 관계
PIES와 식물 줄기 추출액의 EC, 원소 함량의 상관관계
질소 처리가 브로콜리 잎의 엽록소와 프롤린 함량에 미치는 영향
서 언
농업에서 토양의 정밀 양분 관리는 작물, 기후 등을 고려해 적절한 양의 비료를 공급함으로써 작물의 생산성을 유지하고 양분의 환경부하를 경감시킬 수 있다(Kim et al., 2015). 비료 부족 혹은 과잉 공급에 의한 토양 양분의 불균형은 작물의 영양소 결핍과 생육의 불균형을 초래하고, 환경에 부정적인 영향을 줄 수 있어 결과적으로 작물의 스트레스와 생산성의 악화를 초래하고 농업의 지속가능성을 유지하기 어렵게 한다(Biswas and Naher, 2019; Lee et al., 2017). 또한 양분이 결핍되면 식물의 생육에 문제가 생기거나 고사에 이르기도 하며, 양분이 과잉으로 공급되면 작물에 양분이 과잉으로 축적되고, 토양의 물리성이 악화될 수 있어 작물의 생산성과 품질을 저하시킨다(Lee et al., 1989; Seo and Jung, 2008).
토양에서 양분 순환의 불균형은 환경에도 영향을 미칠 수 있다. 토양에 투입되는 질소와 인은 작물의 생육을 증대시키지만 농업에서 양분 과잉에 의한 환경 오염은 수계를 오염시켜 부영양화를 일으키고 온실가스 발생원이 되기도 한다(Vitousek et al., 2009). 따라서 토양 양분의 공급 및 조절은 농경지의 지속 가능성 유지 및 최적의 수확 시기와 최대 수확량을 얻기 위해 중요한 요소이기 때문에 작물을 재배할 때 작물의 생산량과 품질의 향상을 위해 토양 양분의 적절한 관리가 중요하다(Day and Intalap, 1970; Lee et al., 2010; Kim et al., 2019).
일반적으로 토양에 처리되는 비료량은 표준 시비 처리량보다 많은데 이는 작물의 생육 상태를 즉각적으로 판단할 수 없기 때문에 생육 증대를 위해 과잉의 비료를 처리하는 경향이 있기 때문이다. 작물에 양분이 부족한 경우 작물 잎의 색과 상태 등을 통해 육안으로 스트레스를 감지할 수 있지만 이미 생육이 지연되거나 이상 증상이 외부로 나타난 후에 양분을 추가로 처리하는 것은 효과적이지 않을 수 있기 때문에 작물의 상태를 빠르게 평가하는 것이 필요하다. 작물의 상태를 육안으로 평가하는 것 외에 스펙트럼을 이용해 스트레스를 측정하는 새로운 센서와 기기의 출현으로 빠르고 정확하게 스트레스를 평가할 수 있는 수준이 향상되고 있다(Barton, 2012). 그러나 이미지를 이용해 식물 상태를 평가하는 방식은 육안으로 평가하는 것을 수치화시키는 것으로 외부에 이상 증상이 나타난 후 평가한다는 점에서는 육안으로 평가하는 것과 차이가 없다.
식물의 전기적 신호(PIES, plant induced electrical signal)는 식물 줄기 양쪽에 전극을 삽입해 식물 줄기 내부의 저항을 측정하고 전기전도도로 환산하여 나타내며 물관의 양수분 흡수를 반영하므로 이를 이용하면 식물의 생리적인 반응을 비파괴적으로 실시간 모니터링할 수 있다(Park et al., 2018). 식물 내부의 양수분 흡수를 반영하는 PIES는 온도, 습도, CO2 등 식물의 생육 환경과 밀접한 관련이 있으며 이전 연구에서 브로콜리(Brassica oleracea var. italica)의 온도 변화에 따른 스트레스 평가에 사용할 수 있음을 확인하였다(Cha et al., 2020). 그러나 토양 양분 함량이 변화하는 조건이 식물의 생리적 반응에 어떤 영향을 미치며 PIES가 어떻게 반응하는지 알려져 있지 않다. PIES를 이용해 적절한 양분 공급 수준을 설정하기 위해서는 양분 공급과 식물의 생육 및 PIES와의 관계를 평가하는 것이 필요하다. 따라서 본 연구의 목적은 토양 양분 함량의 부족, 과잉 등의 환경 조건이 식물의 생육과 전기적 신호에 어떻게 영향을 미치는지 평가하고 실제 줄기의 이온 함량과 PIES의 관계를 평가하는 것이다.
재료 및 방법
브로콜리의 생육 및 시험구 처리
브로콜리 모종(나이스 그린)은 국립원예특작과학원(전라북도 완주군)의 노지 포장의 토양에 정식하여 6주간 생육시켰다. 정식한 토양은 양토이고 pH는 5.63으로 실제 우리나라 밭토양 재배 환경에서의 토양과 유사한 특성을 나타냈다(Sohn et al., 1999). 브로콜리는 생육 기간 동안 포장용수량 상태로 점적 관수하였다.
브로콜리가 동일한 생육 환경 조건에서 다른 토양 양분 조건에 노출될 때 PIES가 어떻게 반응하는지 평가하기 위해 노지 재배 환경에서 비료 처리를 다르게 하여 실험을 수행하였다. 시료 별 토양 비료 처리는 브로콜리 표준 시비량(Korean Soil Information System, 2021)에 따랐으며, 기비는 처리구별 차이가 없었고 처리구의 경우 부족(N 0)은 질소비료를 추비로 처리하지 않았으며 적정(N 1)은 표준 시비량에 따른 질소비료 적정량인 2.2kg/10a로 요소비료를 처리하였고 과잉(N 1.5)은 1.5배인 3.3kg/10a로 처리하였다. 비료를 처리하고 2주 경과 후 브로콜리를 수확하였다. 브로콜리는 지제부를 수확하여 초장, 직경, 마디수, 무게를 측정하였으며 줄기는 수액 추출에 사용하였고 잎은 증류수로 세척 후 10g은 냉동시켜 엽록소와 프롤린 분석에 사용하였으며 나머지는 60°C 오븐에서 건조하여 원소 함량 분석에 사용하였다.
식물 전기적 신호(PIES) 모니터링
브로콜리를 질소 비료 부족, 적정, 과잉 조건에 노출시키면서 브로콜리의 반응을 PIES로 모니터링하였다. PIES는 브로콜리 지면으로부터 약 5cm 높이의 줄기 양쪽에 3개의 스테인리스 침이 달린 전극을 각각 5mm씩 삽입하여 모니터링하였으며 Junsmeter II (Prumbio, Suwon, Korea)를 사용하여 저항을 측정하고 전기전도도로 식(1)을 이용해 변환하였다(Park et al., 2018).
식(1)에서 PIES는 식물 줄기의 전기전도도(mS·m-1)이고, R은 줄기의 저항, D는 줄기의 직경(mm), L은 줄기에 삽입된 바늘의 길이(mm)이다. d는 바늘의 직경(mm)이고, k는 상수이다.
잎의 엽록소, 프롤린 함량 분석
브로콜리의 잎을 증류수로 세척한 후 작게 분쇄하여 엽록소, 프롤린 함량을 분석하였다. 엽록소 분석을 위해 50mL 코니컬 튜브에 잎 0.5g과 25mL의 80% 아세톤을 첨가하여 혼합하고 24시간 동안 암조건에 두었다. 시간이 경과한 후 엽록소가 추출된 용액을 0.45µm 필터로 여과하고 UV/Vis 분광 광도계(Orion AquaMate 7000, Thermo-Fisher Scientific, Waltham, MA, USA)로 각각 645, 665nm에서의 흡광도를 측정하여 식물 잎의 엽록소 a와 b 함량을 계산하였다(Maclachlan and Zalik, 1963).
프롤린을 잎에서 추출하기 위해 50mL 코니컬 튜브에 0.5g의 브로콜리 잎과 10mL 3% sulfosalicylic acid를 첨가하고 초음파분산기에서 30분간 반응시켰다. 추출된 용액을 25°C 4000rpm 에서 10분간 원심 분리하고 0.45µm 필터로 여과하였다. 여과액 2mL에 acid-ninhydrin reagent (2.5g ninhydrin, 60mL glacial acetic acid, 40mL 6M phosphoric acid) 2mL와 glacial acetic acid 2mL를 첨가하여 혼합하고 100°C에서 1시간 동안 가열하였다(Bates et al., 1973). 가열한 시료를 냉각시킨 후, 4mL의 톨루엔을 첨가하여 발색단을 분리하고 톨루엔을 blank로 하여 520nm에서 흡광도를 측정하였다.
줄기 추출액의 이온 함량 분석
브로콜리를 수확한 후, PIES 측정용 전극을 삽입했던 부분을 포함하여 줄기를 5cm 잘라 무게를 측정하고 1L 비커에 브로콜리 줄기와 400mL의 초순수를 첨가하여 180rpm에서 24시간 교반하였다(White et al., 1985). 추출한 용액의 pH와 EC를 측정하고, 용액을 주사기필터로 여과하여 유도결합플라즈마 분광광도계(ICP-OES, Perkin Elmer)를 사용하여 원소 함량을 분석하였다. 질산태 질소는 Griess 반응을 이용하여 UV-VIS spectrophotometer로 분석하였다(García-Robledo et al., 2014).
브로콜리 토양의 수용성 및 교환성 원소 함량 분석
브로콜리를 수확하면서 뿌리 근처의 토양 시료를 채취하였으며 풍건하여 2mm체를 통과한 시료를 분석에 사용하였다. 토양에 존재하는 유효태 원소 함량을 분석하기 위해 토양 시료 5g을 50mL 코니컬 튜브에 취하여 초순수 25mL와 1N C2H7NO2 50mL를 각각 첨가하여 240rpm에서 30분간 교반하였다. 초순수로 추출한 용액의 pH와 EC를 측정하고 10mL를 취해 주사기 필터로 여과한 후 ICP-OES를 사용하여 원소 함량을 분석하였다. 1N C2H7NO2로 추출한 용액도 같은 방법으로 전처리 하여 ICP-OES를 사용하여 교환성의 양이온 함량을 분석하였다(Schollenberger and Simon, 1945).
데이터 통계처리
데이터의 통계 분석은 SPSS 소프트웨어(IBM, Armonk, NY, USA)를 사용하여 수행하였다. PIES 데이터는 3개의 데이터를 평균하여 평균값으로 나타내었다. PIES와 줄기 추출액의 원소 함량과 상관관계를 분석하였다. 생육, 원소함량, 엽록소, 프롤린 함량은 평균과 표준 편차로 표시하였으며 처리구별 차이를 일원배치 분산분석(One-way ANOVA Analysis)을 이용하여 비교하였고 사후 분석은 Duncan’s multiple range test로 시행하였으며 유의 수준은 0.05로 하였다.
결과 및 고찰
토양 요소 처리와 브로콜리 전기적 신호의 관계
PIES는 식물이 양수분을 흡수할 때 식물 줄기에서 저항의 변화를 측정하여 전극 사이의 거리와 접촉면을 이용해 전기전도도로 계산하는 식물의 전기적 신호이며 식물의 생리적인 반응과 생육의 정도를 평가할 수 있는 지표이다(Park et al., 2018). 따라서 PIES는 토양의 양분, 수분 등의 환경과 밀접한 관계를 가지고 있는데, 본 연구에서 비가 오지 않는 환경에서는 PIES에 큰 차이가 없었으나 비가 올 때 PIES가 처리구에 따라 다르게 나타났으며 적정(N 1, control)의 PIES가 제일 높고 양분 과잉(N 1.5)과 양분 부족(N 0) 순으로 높게 났다(Fig. 1). 이는 비가 오면서 토양의 수분 함량이 증가하고 수분 흡수와 함께 양분 흡수도 증가하기 때문인 것으로 판단된다. 따라서 PIES의 값으로 판단했을 때 비가 올 때 적정 비료 처리구 브로콜리의 양수분 흡수가 제일 좋은 것으로 판단되지만 본 실험에서 처리한 비료량의 차이는 식물 생육과 수확량에 유의미한 차이를 나타낼 정도는 아니었다. 질소 비료의 처리가 항상 식물의 생육을 증가시키는 것은 아니다. 무기질 질소 비료와 유기질 비료를 처리하고 상추의 생육을 평가한 연구에서 유기질 비료는 상추의 생육을 증가시켰으나 무기질 비료를 사용하였을 때 질소를 처리하지 않은 것과 상추 생육이 비슷하게 나타났는데 이는 비료 처리로 인한 토양 EC 상승이 식물에 부정적인 영향을 미쳤기 때문이다(Liu et al., 2014). 양분은 토양의 물리화학적 특성을 향상시켜, 식물의 수확량과 생육을 증대시키지만 과잉시에 식물이 웃자라거나 양분 축적으로 인해 식물 생육이 저하되기 때문에 적정 비료 처리구 보다 PIES가 낮게 측정된 것으로 판단된다(Rudmin et al., 2019; Ingestad, 1977). 양분이 부족할 경우 영양소가 결핍되어 식물의 생육이 저하되기 때문에 양분 과잉 수준보다 식물의 생육이 저해 받아 본 연구에서 PIES가 더 낮게 측정된 것으로 시사된다(Masinde and Agong, 2011). PIES 값은 수확 시기가 지나면서 감소하는 경향을 보여 식물체의 활력이 시간이 지나면서 점점 떨어지는 것으로 판단된다. PIES를 식물의 생육과 연계시키기 위해 생육 지표로 평가한 마디수는 처리구별 차이가 나타났지만 생체중, 초장, 직경 등은 유의미한 차이가 나타나지 않았다(Table 1). 따라서 육안으로 보이는 생육 정도의 차이는 없는 것으로 판단된다.
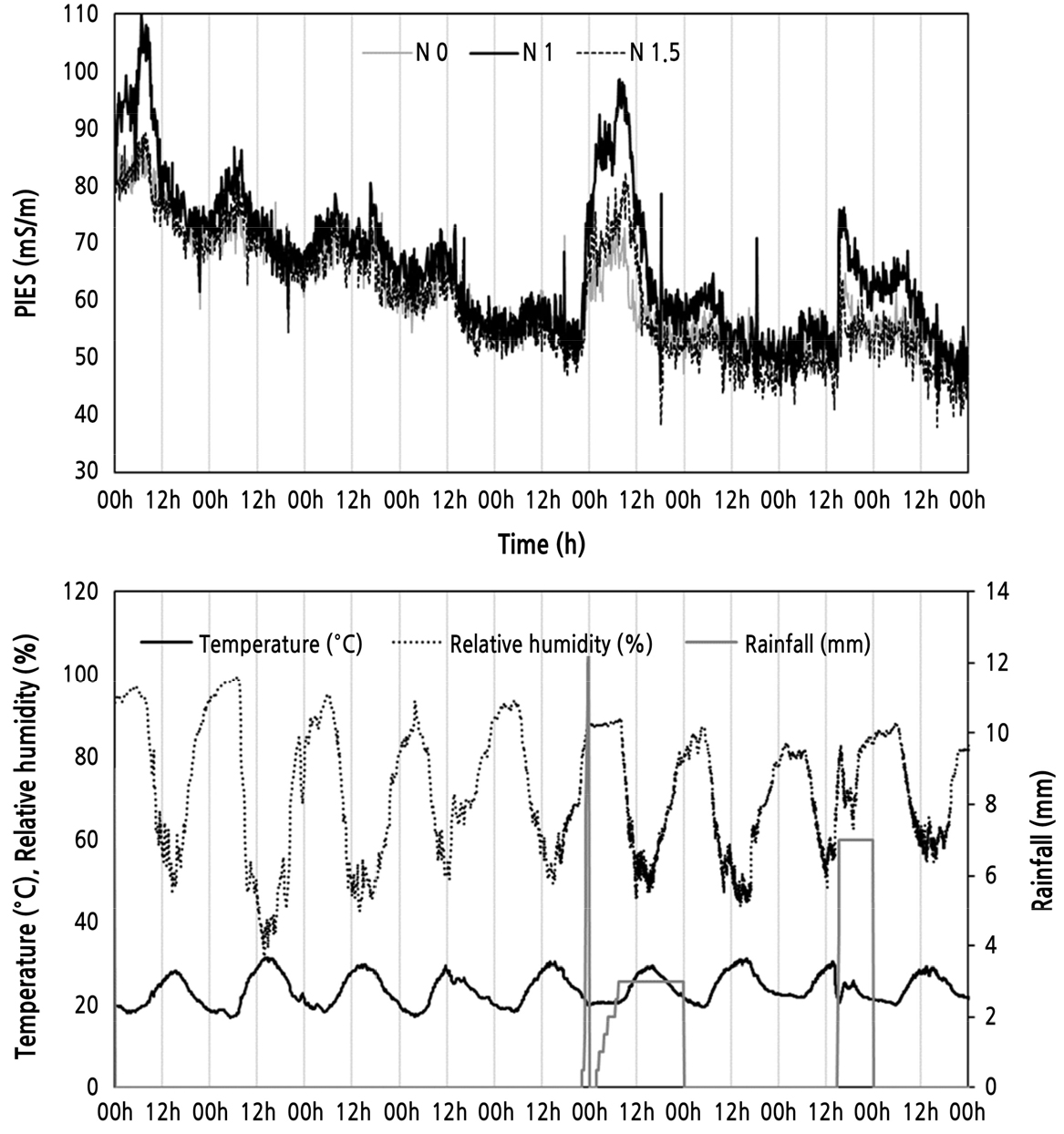
Fig. 1.
Plant-induced electrical signal (PIES) monitored under control (N 1), low urea (N 0), and high urea (N 1.5) conditions and temperature, relative humidity and rainfall during PIES monitoring.
Table 1.
Shoot fresh weight and length, stem diameter, leaf weight, and stem water content of broccoli and pH and EC of soils with different urea application
|
Plant
| |
Soil
|
Shoot fresh
weight (g)
|
Shoot length
(cm)
|
Stem diameter
(mm)
|
Number of leaves
|
Stem water
content (%)
| |
pH
|
EC (dS·m-1)
|
|
N 0
|
3250 ± 309 a
|
59.3 ± 3.1 a
|
44.7 ± 7.8 a
|
83.7 ± 4.0 a
|
93.03 ± 0.30 a
| |
5.53 ± 0.06 b
|
0.085 ± 0.015 a
|
|
N 1
|
3482 ± 1035 a
|
62.12 ± 0.9 a
|
44.9 ± 4.6 a
|
88.3 ± 7.7 a
|
92.74 ± 0.15 a
| |
5.67 ± 0.13 ab
|
0.066 ± 40.0049 a
|
|
N 1.5
|
3891 ± 320 a
|
61.0 ± 1.0 a
|
45.9 ± 4.3 a
|
65.0 ± 4.0 b
|
93.33 ± 0.15 a
| |
5.94 ± 0.13 a
|
0.055 ± 0.0086 a
|
PIES와 식물 줄기 추출액의 EC, 원소 함량의 상관관계
PIES는 식물 줄기에서 양수분이 물관을 통해 이동하는 정도를 전극을 사용하여 측정하기 때문에 PIES와 식물 줄기 추출액의 원소 함량과 상관성이 존재한다(Park et al., 2018; Milham et al., 1970). 따라서 브로콜리를 수확하고 줄기를 물로 추출하여 수용액에 존재하는 다량원소의 함량을 분석하고 브로콜리의 줄기 건조 질량 당 함량으로 계산하였다(Table 2). 토양의 질소 비료 처리에 따른 줄기 추출액의 원소 함량은 비료 과잉 처리구에서 높게 나타났으나 개체간 값의 차이가 커 요소 처리에 따른 값의 통계적인 유의성은 없는 것으로 나타났다. 이는 브로콜리의 생육에 큰 차이가 없었던 것과 일치하는 결과이며 비료의 처리가 식물의 생육에 영향을 미칠 만큼 차이가 없었기 때문인 것으로 판단된다. 토양에서 초순수와 ammonium acetate로 추출 가능한 원소 함량을 분석한 결과에서도 처리별 원소함량에 차이가 없는 것으로 나타났다(Table 3).
Table 2.
Electrical conductivity and macronutrient concentrations of broccoli stem extract (mmol·kg-1)
|
Samples
|
Electrical conductivity (dS·m-1)
|
NO3- |
Ca
|
K
|
Mg
|
P
|
S
|
|
N 0
|
0.12 ± 0.02 c
|
9.4 ± 4.9 a
|
5.6 ± 7.7 a
|
212.4 ± 138.5 a
|
6.0 ± 8.5 a
|
12.4 ± 16.0 a
|
48.6 ± 55.2 a
|
|
N 1
|
0.81 ± 0.19 b
|
6.2 ± 1.2 a
|
6.7 ± 0.5 a
|
191.9 ± 13.1 a
|
5.0 ± 0.8 a
|
15.1 ± 2.8 a
|
54.8 ± 22.5 a
|
|
N 1.5
|
1.37 ± 0.01 a
|
6.7 ± 1.1 a
|
14.2 ± 0.5 a
|
322.2 ± 31.4 a
|
15.2 ± 1.4 a
|
30.9 ± 6.2 a
|
168.0 ± 108.5 a
|
Table 3.
Nutrient concentrations in soil extracted with water and ammonium acetate (mg·kg-1)
|
Extractant
|
Samples
|
Ca
|
K
|
P
|
S
|
|
H2O
|
N 0
|
0.9 ± 0.3 a
|
1.5 ± 0.4 a
|
0.13 ± 0.02 a
|
3.9 ± 0.7 a
|
|
N 1
|
0.6 ± 0.2 a
|
1.0 ± 0.1 a
|
0.10 ± 0.02 a
|
3.1 ± 0.6 ab
|
|
N 1.5
|
0.4 ± 0.2 a
|
0.8 ± 0.1 a
|
0.08 ± 0.05 a
|
2.0 ± 0.4 b
|
|
NH4OAC
|
N 0
|
70.4 ± 7.7 a
|
10.5 ± 0.9 a
|
0.05 ± 0.01 a
|
5.5 ± 1.1 a
|
|
N 1
|
71.5 ± 7.5 a
|
8.5 ± 0.7 ab
|
0.04 ± 0.01 a
|
4.7 ± 0.9 a
|
|
N 1.5
|
66.3 ± 4.6 a
|
7.6 ± 0.2 b
|
0.04 ± 0.001 a
|
3.1 ± 0.7 a
|
PIES가 브로콜리 개체별 줄기 추출액의 원소 함량과 관계가 있는지 알아보기 위해 줄기추출액의 EC와 원소 함량과의 상관관계를 분석하였다. PIES 값이 높을 때 양수분의 이동이 활발하다는 것을 의미하기 때문에 PIES 값이 높은 시간대인 오전 시간의 PIES 값을 평균하여 줄기 추출액의 원소 함량과 상관 관계를 분석하였다(Table 4). PIES는 NO3-와 유의한 양의 상관관계가 있는 것으로 나타났으나 다른 원소와는 상관관계가 나타나지 않았다(Table 4). 본 연구에서는 질소비료의 처리를 달리하였기 때문에 줄기의 질소 이동을 PIES가 잘 반영하는 것으로 판단된다. 식물이 적정 생육 온도에서 질산염을 식물체 내에 저장하는 양과 질소를 고정하는 정도에 상관 관계가 존재하므로 줄기의 질산태 질소 함량은 질소 동화와 관계가 있다(Herridge, 1982). 암모늄태 질소는 독성이 있어 줄기에서 이온 상태로 존재하기보다 동화되어 줄기에 축적되어 있는 양은 많지 않으며 질산태 질소의 축적이 더 우세하여 PIES와 상관관계가 높은 것으로 시사된다(Salsac et al., 1987; Magalhaes and Huber, 1991; McNeill and Unkovich, 2007).
Table 4.
Correlation between PIES, EC and nutrients of broccoli stem
|
PIES
|
Stem EC
|
NO3- |
Ca
|
K
|
Mg
|
P
|
S
|
|
PIES
|
1
| | | | | | | |
|
Stem EC
|
–0.078
|
1
| | | | | | |
|
NO3- |
0.803* |
–0.389
|
1
| | | | | |
|
Ca
|
–0.280
|
0.739* |
–0.617
|
1
| | | | |
|
K
|
–0.536
|
0.527
|
–0.715* |
0.897** |
1
| | | |
|
Mg
|
–0.272
|
0.688
|
–0.548
|
0.987** |
0.920** |
1
| | |
|
P
|
–0.149
|
0.746* |
–0.533
|
0.980** |
0.847** |
0.971** |
1
| |
|
S
|
–0.280
|
0.704
|
–0.448
|
0.794* |
0.632
|
0.772* |
0.705
|
1
|
식물 줄기 수액의 원소들은 대부분 이온의 형태로 이동하며, 전기 전도도는 이온이 얼마나 존재하는지를 나타내는 것이므로 EC와 줄기 추출액의 이온 농도는 상관관계가 높을 것으로 추정 가능하다(Pate and Jeschke, 1995; Nardini et al., 2010). 그러나 질산태 질소 외의 다른 원소들과는 상관관계가 높지 않았는데 이는 PIES의 경우 물관부의 양수분 이동을 측정하며 줄기 추출액은 물관부의 양분만 추출하는 것이 아니라 줄기 유세포의 세포질에 존재하는 양분도 추출되었기 때문인 것으로 판단된다. 줄기의 물관과 목부유조직에서는 아포플라스틱 경로를 통해 양분이 평형 상태에 이를 때까지 자유롭게 교환되며 줄기 세포질의 다양한 부분에서 K, Ca, Mg 등이 발견된다(Metzner et al., 2010). 질소 처리량이 높을 때 통계적으로 원소 함량에 유의한 차이는 없었으나 적정 처리구와 과잉 처리구를 비교했을 때 대부분의 원소 함량이 과잉 처리구에서 높게 나타났다. 요소를 식물에 처리했을 때 확산성 양이온과 음이온이 모두 증가되었으며 유기산의 함량 또한 증가되는 선제 연구를 통해 질소 처리가 식물 체내에 존재하는 양이온과 음이온의 균형을 조절하고 증가시켜 영향을 주는 것으로 판단된다(Yang et al., 2007). 식물이 이온을 흡수할 때, 양이온과 음이온의 균형을 유지하는 것이 생리 활성의 조절이나 근권 pH를 조절할 때 중요한 요소로 작용되기 때문에 줄기에서 원소들 간의 상관관계가 높게 나타났다(Haynes, 1990).
브로콜리를 수확하고 토양을 채취해 토양 추출액의 EC를 분석하였으나, 토양 질소 처리량에 따른 유의미한 차이가 나타나지 않았으며 용액의 양분 함량을 ICP-OES로 분석한 결과, EC는 수용액의 Ca와 0.943, S와 0.900, 총 이온과 0.909 등으로 모든 이온들과 상관관계가 높게 나타났다(data not shown). 그러나 PIES의 경우 토양의 EC와 수용액 혹은 유효태의 원소 함량과 상관 관계가 낮게 나타났는데 이는 비료를 토양에 처리한 직후의 함량을 분석한 것이 아닌 수확 시 토양을 채취해 분석했기 때문에 식물의 양분 흡수로 인해 상관 관계가 낮게 나타난 것으로 판단된다(Baligar et al., 2001).
질소 처리가 브로콜리 잎의 엽록소와 프롤린 함량에 미치는 영향
토양 질소가 증가하는 조건에서 엽록소 a와 b 함량은 적정 처리구인 N 1에 비해 높았으며, 토양 질소가 감소하는 조건에서는 N 1보다 낮았다(Fig. 2). 이전 연구에서도 총질소의 함량이 높은 조건에서 엽록소 함량은 높게 나타났는데 이는 질소가 단백질을 구성하는 원소이기 때문에 함량이 높을수록 켈빈 회로에서 틸라코이드 형성을 도와 엽록소가 증가되기 때문인 것으로 시사된다(Evans, 1989; Hamilton and Mitchell, 1997). 프롤린은 다양한 스트레스 환경에서 식물의 반응을 조절하는 것으로 알려져 있으며 특히 염 스트레스 조건에서 함량이 증가하고 삼투 조절에 중요하다(Eum et al., 2020; Shin et al., 2020; Sun et al., 2020). 본 연구에서 프롤린 함량은 적정 질소처리구의 값이 비교적 높게 나타났으나 모든 처리구에서 식물이 스트레스를 받아 축적된 수준은 아니었다(Fig. 3). 프롤린 함량이 약 10% 정도는 일반적으로 스트레스를 받지 않은 식물에서도 나타나며 비료 부족이나 과잉 처리구에서도 프롤린 함량이 높지 않아 식물이 양분에 대한 스트레스를 받지 않은 것으로 판단된다(Lehmann et al., 2010; Sharma and Verslues, 2010). 암모늄의 공급량에 따라 식물의 프롤린의 함량이 영향을 받을 수 있지만 본 논문의 질소 처리에 따라 큰 차이가 나타나지 않았으며 식물의 생육량을 평가했을 때 토양의 양분이 생육에 부족한 상태가 아니었기 때문에 식물이 스트레스를 받지 않은 것으로 판단된다(Claussen, 2002; Houdusse et al., 2005).
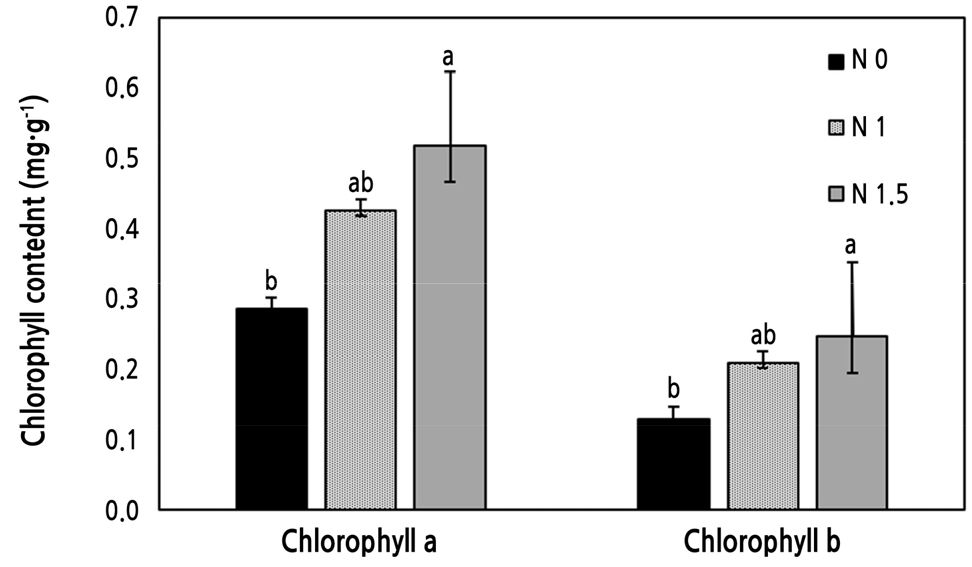
Fig. 2.
Chlorophyll a and b contents in broccoli leaves grown under control (N 1), low urea (N 0), high urea (N 1.5) conditions. Data are presented as mean ± standard deviation. Bars with different letters indicate significant differences (p < 0.05) among means using one-way ANOVA followed by a Duncan post-hoc test.
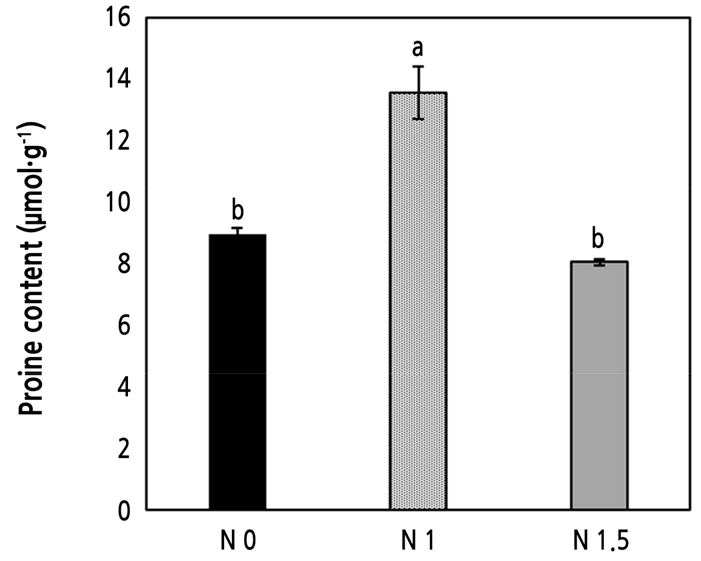
Fig. 3.
Proline content in broccoli leaves grown under control (N 1), low urea (N 0), high urea (N 1.5) conditions. Data are presented as mean ± standard deviation. Bars with different letters indicate significant differences (p < 0.05) among means using one-way ANOVA followed by a Duncan post-hoc test.
Acknowledgements
본 연구는 농촌진흥청 공동연구사업(세부과제번호: PJ015050012021)의 지원에 의해 수행되었음.
References
Baligar VC, Fageria NK, He ZL (2001) Nutrient use efficiency in plants. Commun Soil Sci Plant Anal 32:921-950 doi:10.1081/CSS-100104098
10.1081/CSS-100104098Barton CV (2012) Advances in remote sensing of plant stress. Plant Soil 354:41-44. doi:10.1007/s11104-011-1051-0
10.1007/s11104-011-1051-0Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205-207. doi:10.1007/BF00018060
10.1007/BF00018060Biswas JC, Naher UA (2019) Soil nutrient stress and rice production in Bangladesh. In Advances in Rice Research for Abiotic Stress Tolerance 431-445. doi:10.1016/B978-0-12-814332-2.00021-6
10.1016/B978-0-12-814332-2.00021-631859431Cha SJ, Park HJ, Lee JK, Kwon SJ, Jee HK, Baek, H, Park JH (2020) Multi-sensor monitoring for temperature stress evaluation of broccoli (Brassica oleracea var. italica). J Appl Biol Chem 63:347-355. doi:10.3839/jabc.2020.046
10.3839/jabc.2020.046Claussen W (2002) Growth, water use efficiency. and proline content of hydroponically grown tomato plants as affected by nitrogen source and nutrient concentration. Plant Soil 247:199-209. doi:10.1023/A:1021453432329
10.1023/A:1021453432329Day AD, Intalap S (1970) Some effects of soil moisture stress on the growth of wheat (
Triticum aestivum L. em Thell.). Agron J 62:27-29. doi:10.2134/agronj1970.00021962006200010009x
10.2134/agronj1970.00021962006200010009xEum HL, Yi TG, Hong SJ, Park NI (2020) Variations of bioactive compound contents and antioxidant capacity of asparagus seedlings in 23 varieties. Hortic Sci Technol 38:291-302
Evans JR (1989) Photosynthesis and nitrogen relationships in leaves of C 3 plants. Oecologia 78:9-19. doi:10.1007/BF00377192
10.1007/BF0037719228311896García-Robledo E, Corzo A, Papaspyrou S (2014) A fast and direct spectrophotometric method for the sequential determination of nitrate and nitrite at low concentrations in small volumes. Mar Chem 162:30-36. doi:10.1016/j.marchem.2014.03.002
10.1016/j.marchem.2014.03.002Hamilton D, Mitchell S (1997) Wave‐induced shear stresses, plant nutrients and chlorophyll in seven shallow lakes. Freshw Biol 159-168. doi:10.1046/j.1365-2427.1997.00202.x
10.1046/j.1365-2427.1997.00202.xHaynes RJ (1990) Active ion uptake and maintenance of cation-anion balance: A critical examination of their role in regulating rhizosphere pH. Plant Soil 126:247-264. doi:10.1007/BF00012828
10.1007/BF00012828Herridge DF (1982) Relative abundance of ureides and nitrate in plant tissues of soybean as a quantitative assay of nitrogen fixation. Plant Physiol 70:1-6. doi:10.1104/pp.70.1.1
10.1104/pp.70.1.116662425PMC1067075Houdusse F, Zamarreño AM, Garnica M, García-Mina J (2005) The importance of nitrate in ameliorating the effects of ammonium and urea nutrition on plant development: the relationships with free polyamines and plant proline contents. Funct Plant Biol 32:1057-1067. doi:10.1071/FP05042
10.1071/FP0504232689201Ingestad T (1977) Nitrogen and plant growth; maximum efficiency of nitrogen fertilizers. Ambio, Stockholm, Sweden, pp 146-151
Kim KC, Ahn BK, Ko DY, Lee CK, Cheong SS, Song YJ, Ko YS (2015) Chemical properties of paddy soils in Jeollabuk-do area. Korean J Soil Sci Fert pp 143-143
Kim YH, Kong MS, Lee EJ, Lee TG, Jung GB (2019) Status and changes in chemical properties of upland soil from 2001 to 2017 in Korea. Korean J Environ Agric 38:213-218. doi:10.5338/KJEA.2019.38.3.28
10.5338/KJEA.2019.38.3.28Korean Soil Information System (2021) Setting the standard fertilization amount for autumn cropped broccoli in an open field cultivation area. http://soil.rda.go.kr/soil/sibi/cropSibiPrescript.jsphttp://www.nongsaro.go.kr, Accessed 1 June 2021
Lee CR, Ok JH, An MS, Lee SB, Park KL, Hong SG, Park CB (2017) Soil chemical properties of long-term organic cultivation upland. Korean J Organic Agric 25:161-170. doi:10.11625/KJOA.2017.25.1.161
10.11625/KJOA.2017.25.1.161Lee IH, Yuk CS, Park H (1989) Yield and missing plant rate of Panax ginseng affected by the annual change in physico-chemial properties of ginseng cultivated soil. Korean J Soil Sci Fert 22:18-24
Lee YH, Lee ES, Kim CD, Song CW (2010) Soil management of facility horticultural crops and Soil Sciences and Fertilizer Conference, Suwon, Korea, soil from 2001 to 2017 in Korea, Korean J Environ Agric 38:213-218
Lehmann S, Funck D, Szabados L, Rentsch D (2010) Proline metabolism and transport in plant development. Amino Acids 39:949-962. doi:10.1007/s00726-010-0525-3
10.1007/s00726-010-0525-320204435Liu CW, Sung Y, Chen BC, Lai HY (2014) Effects of nitrogen fertilizers on the growth and nitrate content of lettuce (
Lactuca sativa L.). Int J Environ Res Public Health 11:4427-4440. doi:10.3390/ijerph110404427
10.3390/ijerph11040442724758896PMC4025000Maclachlan S, Zalik S (1963) Plastid structure, chlorophyll concentration, and free amino acid composition of a chlorophyll mutant of barley. Can J Bot 41:1053-1062. doi:10.1139/b63-088
10.1139/b63-088Magalhaes JR, Huber DM (1991) Response of ammonium assimilation enzymes to nitrogen form treatments in different plant species. J Plant Nutr Soil Sci 14:175-185. doi:10.1080/01904169109364193
10.1080/01904169109364193Masinde PW, Agong SG (2011) Plant Growth and Leaf N of Spiderplant (
Cleome gynandra L.) Genotypes under varying nitrogen supply (No. RESEARCH)
McNeill A, Unkovich M (2007) The nitrogen cycle in terrestrial ecosystems. In Nutrient cycling in terrestrial ecosystems Springer, Berlin, Heidelberg, pp 37-64. doi:10.1007/978-3-540-68027-7_2
10.1007/978-3-540-68027-7_2Metzner R, Schneider HU, Breuer U, Thorpe MR, Schurr U, Schroeder WH (2010) Tracing cationic nutrients from xylem into stem tissue of French bean by stable isotope tracers and cryo-secondary ion mass spectrometry. Plant Physiol 152:1030-1043. doi:10.1104/pp.109.143776
10.1104/pp.109.14377619965970PMC2815875Milham PJ, Awad AS, Paull RE, Bull JH (1970) Analysis of plants, soils and waters for nitrate by using an ion-selective electrode. Analyst 95:751-757. doi:10.1039/an9709500751
10.1039/an9709500751Nardini A, Grego F, Trifilò P, Salleo S (2010) Changes of xylem sap ionic content and stem hydraulics in response to irradiance in Laurus nobilis. Tree Physiology 30:628-635. doi:10.1093/treephys/tpq017
10.1093/treephys/tpq01720339142Park HJ, Park JH, Park KS, Ahn TI, Son JE (2018) Nondestructive measurement of paprika (
Capsicum annuum L.) internal electrical conductivity and its relation to environmental factors. Hortic Sci Technol 31:691-701. doi:10.12972/kjhst.20180069
10.12972/kjhst.20180069Pate JS, Jeschke WD (1995) Role of stems in transport, storage, and circulation of ions and metabolites by the whole plant. Plant Stems, Academic Press, pp 177-204. doi:10.1016/B978-012276460-8/50010-2
10.1016/B978-012276460-8/50010-2Rudmin M, Banerjee S, Makarov B, Mazurov A, Ruban A, Oskina Y, Shaldybin M (2019) An investigation of plant growth by the addition of glauconitic fertilizer. Appl Clay Sci 180:105178. doi:10.1016/j.clay.2019.105178
10.1016/j.clay.2019.105178Salsac L, Chaillou S, Morot-Gaudry JF, Lesaint CH, Jolivet E (1987) Nitrate and ammonium nutrition in plants. Plant Physiol, Biochem, Paris 25:805-812. doi:10.1007/978-94-009-4356-8_44
10.1007/978-94-009-4356-8_44Schollenberger CJ, Simon RH (1945) Determination of exchange capacity and exchangeable bases in soil-ammonium acetate method. Soil Science 59:13-24. doi:10.1097/00010694-194501000-00004
10.1097/00010694-194501000-00004Seo YH, Jung YS (2008) Role of Chemical Fertilizer in Terms of Nutrients Balace. KSSSF:50-62
Sharma S, Verslues PE (2010) Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant Cell Environ 33:1838-1851. doi:10.1111/j.1365-3040.2010.02188.x
10.1111/j.1365-3040.2010.02188.x20545884Shin YK, Bhandari SR, Cho MC, Lee JG (2020) Evaluation of chlorophyll fluorescence parameters and proline content in tomato seedlings grown under different salt stress conditions. Hortic Environ Biotechnol 61:433-443. doi:10.1007/s13580-020-00231-z
10.1007/s13580-020-00231-zSohn BK, Cho JS, Kang JG, Cho JY, Kim KY, Kim HW, Kim HL (1999) Physico-chemical properties of soils at red pepper, garlic and onion cultivation areas in Korea. Korean J Soil Sci Fert 32:123-131
Sun S, Qi X, Wang R, Lin M, Fang J (2020) Evaluation of freezing tolerance in Actinidia germplasm based on relative electrolyte leakage. Hortic Environ Biotechnol 61:755-765. doi:10.1007/s13580-020-00272-4
10.1007/s13580-020-00272-4Vitousek PM, Naylor R, Crews T, David MB, Drinkwater LE, Holland E, Johnes PJ, Katzenberger J, Martinelli LA, et al. (2009) Nutrient imbalances in agricultural development. Science 324:1519-1520. doi:10.1126/science.1170261
10.1126/science.117026119541981White JW, Cook ER, Lawrence JR (1985) The DH ratios of sap in trees: Implications for water sources and tree ring DH ratios. Geochim Cosmochim Acta 49:237-246. doi:10.1016/0016-7037(85)90207-8
10.1016/0016-7037(85)90207-8Yang C, Chong J, Li C, Kim C, Shi D, Wang D (2007) Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil 294:263-276. doi:10.1007/s11104-007-9251-3
10.1007/s11104-007-9251-3

