

서 언
최근 들어 멜론의 맛과 가격이 적정 수준으로 유지되면서 국내 소비량이 증가하고 있으며, 일부 지역에서는 해외로 수출까지 하고 있어 농업인의 큰 소득원으로 자리잡고 있다. 우리나라에서는 멜론이 대부분 시설 내에서 재배되기 때문에 연중 생산이 가능한데 특히 6-9월까지 생산되는 여름작형에 가장 많이 재배되고 있다. 그런데 멜론 시설재배지에서는 몇 년 전부터 잎이 황화되는 증상이 발생하기 시작하였는데, 그 발생 지역과 면적이 점차 확대되면서 멜론 안정생산에 문제가 되고 있다(Park et al., 2011). 황화엽 증상은 정상엽에 비해서 잎이 황화증상을 보이고, 처음에는 하위엽에서 시작하여 점점 상위엽으로 진행된다. 황화엽이 발생되면 생육이 더디고, 과실 비대가 느려지며 네트 형성이 되지 않고, 당도가 낮아지는 등 상품과 생산이 어렵다(Park et al., 2011). 이러한 멜론의 황화엽 증상은 재배상 환경조건에 의해 발생되는 생리적인 원인과 바이러스 이병에 의해 나타나는 것으로 추정되지만(Park et al., 2011; Takeshita, 2004) 정확한 원인은 아직 밝혀지지 않았다. 진딧물 매개에 의한 바이러스 감염으로 발생하는 황화증상은 생리적인 원인에 의해 발생할 수 있는 황화증상과 유사하지만 멜론에 있어서는 서로 다른 원인에 의한 것으로 추정되고 있으며, 생리적인 원인에 의한 것은 뿌리 양이 적거나 잎이 크고 넓은 품종으로 과실비대가 잘되는 품종에서 많이 나타난다고 보고되었다(Takeshita, 2004). 멜론은 과실 성숙기에 서서히 시들다가 갑작스럽게 황화가 진전되면서 품질 저하를 초래하여 경제적 손실을 입힌다고 하였으며(Lee et al., 2009), 오이에서는 근권온도가 15°C에서 황화증상이 일어나며 잎이 어리고 저온에 노출되는 시간이 길어질수록 황화증상이 더욱 급속하게 진전된다고 하였다(Moon, 2001).
전 세계적으로 박과작물에서 발생되는 바이러스는 Cucumber mosaic virus(CMV; Enzie, 1943), Watermelon mosaic virus (WMV; Diaz-Pendon et al., 2005), Cucumber green mottle mosaic virus(CGMMV; Rajamony et al., 1987), Melon necrotic spot virus(MNSV; Choi et al., 2003; Kim et al., 2008; Kim et al., 2012), Squash mosaic virus(SqMV; Avgelis and Katis, 1989), Papaya ringspot virus(PRSV; Bateson et al., 1994; Grafton-Cardwell et al., 1996) Zucchini yellow mosaic virus (ZYMV; Dukic et al., 2002; Papayiannis et al., 2005; Vucurovic et al., 2009), Kyuri green mottle mosaic virus(KGMMV; Choi, 2001; Tan et al., 2000), Cucurbit aphid-borne yellows virus(CABYV; Guilley et al., 1994) 등 39종 이상의 바이러스에 감염되는 것으로 알려져 있다(Lecoq et al., 2003). 그 중에서도 CABYV는 프랑스에서 1992년에 처음 발생이 보고되었고(Lecoq, 1999), CABYV 감염 멜론의 경우 40% 정도의 상품 수량이 감소한다고 보고된 바 있지만(Lecoq, 1999; Lecoq et al., 2012) 아직까지 국내에서 발생되었다는 보고(Ryu, 2009)는 없다. 따라서 본 실험은 우리나라 멜론재배지에서 발생되고 있는 황화엽 증상의 발생 원인과 황화엽 발생이 근활력 및 광합성에 미치는 영향을 다양한 각도에서 구명하고자 수행되었다.
재료 및 방법
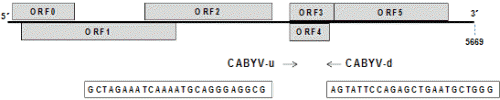
경북 구미, 전북 남원 및 충남 청양지역에서 황화증상을 보이는 멜론 식물체를 채집하고 국내에 보고되어 있는 박과 감염 바이러스 8종(Table 1)에 대하여 전자현미경과 RT-PCR 검정을 수행하였다. 또한 국내 미보고된 새로운 바이러스의 진단을 위해 차세대유전체염기서열분석(Next generation sequencing, NGS)을 이용하여 바이러스 감염여부를 진단하였다. NGS 및 Bioinformatics 분석은 (주) C&K Genomics (Seoul, Korea)에 의뢰하여 수행하였다. NGS는 Illumina사의 프로토콜에 따라 Hiseq2000을 이용하여 Read length 101bp paired-end로 수행되었으며 약 16.8Gbp의 데이터를 생산하였다. 생산된 NGS데이터의 분석을 위해 NCBI에 있는 식물 바이러스의 sequence DB를 구축하였고 이 Database에 Short read aligner인 Bowtie2(Langmead and Salzberg, 2012)를 이용하여 생산된 NGS데이터를 Align하였다. Read mapping Coverage > 90%, average mapping depth > 10×를 만족하는 reference virus sequence 중 가장 높은 mapping coverage와 depth를 보이는 시퀀스를 후보 바이러스로 선발하였다. 이렇게 선발된 virus sequence와 NGS 데이터를 이용하여 genome re-sequencing을 수행하였으며(Carver et al., 2010; Li et al., 2009; McKenna et al., 2010), 변이를 발굴하고 이 변이와 reference virus sequence를 이용하여 샘플 내에 포함되어 있는 virus 유전체의 염기서열을 재구성하였다. 재구성된 virus sequence는 계통 유연관계 확인을 위하여 기존에 알려진 같은 종류의 virus complete sequence와 함께 phylogenetic 분석에 사용되었다. Phylogenetic tree는 Mega 6.0(Tamura et al., 2013)을 이용하여 수행되었으며 muscle sequence alignment, neighbor Joining method with bootstrap 1,000×, maximum composite likelihood 기반의 consensus tree로 분석되었다. Bioinformatics 분석을 통해 진단된 바이러스의 RT-PCR 진단법 확립을 위해서 CABYV 특이 프라이머를 선발하였다(Fig. 1). RT-PCR은 total RNA 1μL을 사용하여 10μmol 농도의 정방향과 역방향 primer를 각각 0.5μL, DW 7μL와 2x RT-PCR premix 10μL(Genetbio SR-8000, Korea)를 첨가하여 전체 20μL로 반응을 수행하였다. RT-PCR 조건은 50°C, 30분의 역전사 반응과 95°C, 10분의 변성 후, 95°C에서 30초, 55°C에서 30초, 72°C에서 1분 과정을 35회 반복 후, 마지막으로 72°C에서 5분간 반응시켰다. PCR 산물은 1% agarose gel에서 60분간 전기영동 후 ethidium bromide로 염색하여 UV광 하에서 확인하였다.

멜론 황화엽증이 발생된 개체의 잎과 정상개체의 잎을 대상으로 광합성능력과 뿌리의 활력을 조사하였다. 광합성특성은 광합성측정기(LI-6400, Portable photosynthesis system, LI-COR Inc., Nebraska, USA)를 사용하여 정상엽과 황화엽을 2주씩 3반복으로 조사하였다. 광합성 측정기의 측정 조건은 온도 25°C, 상대습도 60%, CO2 농도는 400ppm, 광량은 1,000µmol・m-2・s-1로 하였다. 뿌리의 활력 조사는 Berridge et al.(2005)의 방법으로 하였는데, 발생된 포장 내에서 황화엽 증상(Fig. 2)을 보인 것과 정상적인 주를 대상으로 2주씩 3반복으로 조사하였다. 뿌리 채취는 수확 3주 전에 정식부위를 중심으로 반경 50cm, 깊이 30cm 부위의 뿌리를 채취하여 흐르는 물에 씻은 후 측정에 이용하였다. 뿌리 시료는 세근을 약 0.5cm 길이로 절단하여 균일하게 혼합한 후 0.1g을 취하여 test tube에 넣고 1mL의 증류수와 혼합한 후 Premix WST-1 cell proliferation assay system(Takara Inc., Tokyo, Japan) 시약 10µL를 가하였다. 이것을 암상태로 25°C에서 3시간 동안 반응시킨 후 ELISA reader(Microplate Spectro-photometer, EonTM, BioTek Inc., Vermont, USA)를 이용하여 흡광도 420nm로 분석하였다. Premix WST-1 cell proliferation assay system 시약 10µL를 증류수에 혼합하여 blank로 이용하였다. 세포조직 현미경 촬영은 Chang(1973)의 방법으로 하였는데, 채취한 잎을 1차 고정액 2.5% glutaraldehyde에 넣은 즉시 모든 과정은 4°C에서 진행되었으며 1차 고정 90분간 처리, 0.1M phosphate buffer(pH 7.2)로 15분 간격 4-5회 세척, 2차 고정 1% osmium tetroxide 90분간 처리, 위와 동일한 세척 과정 후 하룻밤을 침지시켰다. 탈수는 상온에서 40, 60, 80, 90, 95% ethanol로 각각 5분씩, 100% ethanol로 5, 15, 15, 30분간 처리로 이루어졌으며 propylene oxide로 치환 후 최종적으로 epon에 포매(embedding)하여 60°C의 오븐에서 4일간 중합시켰다. 중합된 epon block을 초미세절편기(Ultracut R ultramicrotome, Leica Mycrosystems, North Ryde, Australia)를 이용하여 1,500nm의 두께로 시료를 절단하여 periodic acid and Schiff’s reagent 염색법으로 염색한 후 광학현미경(Axioskop 2, Carl Zeiss, Oberkochen, Germany) 100배로 검경하였다.
|
Fig. 3. Symptoms in melons infected with Cucurbit aphid-borne yellows virus. A, Infection on 20 days after transplanting; B, Infection on 60 days after transplanting. |

잎의 무기성분은 수액분석을 위해 절단하였던 개체의 잎을 채취하여 80°C가 유지되는 건조기에서 3일간 건조한 후 분말을 만들어서 분석에 사용하였다. 식물체 분석은 토양화학분석법(NIAST, 2000)의 식물체 분석법에 준하여 원소흡광분석기(AA-6800, Shimadzu, Kyoto, Japan)를 사용하여 분석하였다.
통계분석은 SAS 프로그램(SAS 9.2, SAS Institute Inc., USA)을 이용하여 t 검정으로 수행한 후 그 결과를 표에 나타냈다.
결과 및 고찰
경북 구미지역에서 채취된 황화증상을 보이는 멜론에 대하여 전자현미경 검경 및 국내 보고(Ryu, 2009)된 박과 감염 바이러스 8종(CMV, MNSV, CGMMV, SqMV, WMV, KGMMV, PRSV, ZYMV)에 대해 RT-PCR한 결과, 바이러스가 진단되지 않았다(data not shown). 이에 국내에 보고되지 않은 새로운 바이러스로 의심되어 차세대유전체염기서열분석(NGS)을 이용하여 진단한 결과, 박과진딧물매개바이러스(CABYV)로 판정되었고, 전체 contig sequence를 계통 분석한 결과 CABYV 일본에서 보고된 바이러스와 상동성이 높은 것으로 확인되었다(Table 2).
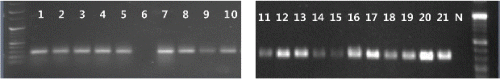
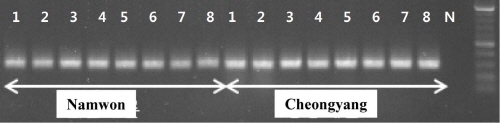
CABYV 진단 프라이머 중 CABYV 특이 프라이머를 선발하여 황화증상(Fig. 3)을 보이는 구미에서 채취한 멜론 21점을 검정한 결과, 20점이 CABYV에 감염된 것을 확인할 수 있었고, 나머지 1점은 다른 요인에 의해서 황화증상을 보인 것으로 판단된다(Fig. 4). 또한 전북 남원과 충남 청양지역으로부터 황화 증상을 보이는 시료 각각 8점을 CABYV 특이 프라이머를 이용하여 RT-PCR한 결과, 모두 CABYV 감염이 확인되었다(Fig. 5).
Table 2. List of candidate complete genome sequencing of melon sample. |
|
|
Fig. 4. RT-PCR results of melon using the CABYV specific primers. N, negative control. |
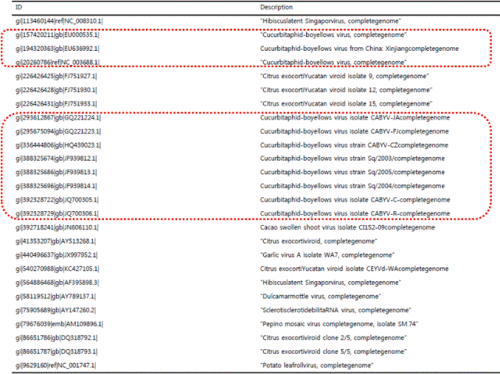
CABYV는 Luteoviridae과 Polerovirus속에 속하는 직경 23nm의 구형바이러스로 진딧물에 의해서 영속전염(순환형)되는 것으로 알려져 있으며, 프랑스(Lecoq, 1999)에서 처음 보고된 이후 이탈리아(Tomassoli and Meneghini, 2007), 스페인(Juarez, 2004), 튀니지(Mnari, 2005), 이집트(Omar and Bagdady, 2012), 미국(Lemaire et al., 1993) 및 네팔(Dahal et al., 1997), 레바논(Abou-Jawdah et al., 1997), 중국(Xiang et al., 2008) 등 아시아 지역에서 발생 보고가 있었다. CABYV에 감염된 멜론은 잎에서는 퇴록반점, 모자이크, 황화 등의 증상을 일으키며, 이병된 과실에서는 불규칙한 네트 형성으로 상품성이 전혀 없으며 초기 감염주는 과실 형성이 되지 않는 것으로 알려져 있다. 또한 CABYV는 호박, 수박, 오이, 상추 등 넓은 기주범위를 가지고 있고, 보독 진딧물은 영속 전염됨에 따라 주변 및 먼 거리 이동으로 인한 확산 가능성이 매우 높을 것으로 판단된다.
멜론 정상엽과 황화엽의 광합성 특성은 Table 3과 같다. 광합성 능력은 정상엽의 경우 12.36µmol・m-2・s-1 정도이었고, 황화엽은 4.09µmol・m-2・s-1로 정상엽과 비교하여 1/3수준 정도로 낮았다. 이와 같은 결과는 황화증상으로 인하여 잎 내 엽록소가 파괴되어 클로로필 함량이 낮아져서 광합성 능력이 떨어진 결과로 판단되며, 멜론의 황화엽 발생시 광합성 능력이 낮아졌다는 Takeshita(2004)와 Park 등(2011)의 보고와 유사한 결과이다.
뿌리의 활력은 정상주의 뿌리는 0.48mg・g-1이었으나 황화주는 0.28mg・g-1로 정상주의 뿌리 활력이 황화주보다 1.7배정도 높은 것을 확인할 수 있었다. Takeshita(2004)는 뿌리의 양이 적은 품종이 황화증상의 발생에 영향을 미칠 가능성이 높다고 하였는데, 본 연구의 황화엽 발생주와 정상주의 뿌리 양은 큰 차이를 보이지 않았다. 그러나 정상주보다 낮은 뿌리 활력이 황화증상이 나타난 개체의 양수분 흡수에 영향을 미쳤을 것으로 판단된다.
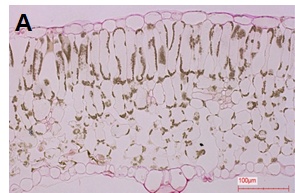
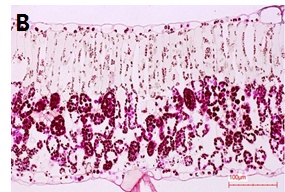
정상주의 잎과 황화엽 발생개체의 잎을 채취하여 광학 현미경으로 잎의 세포조직을 검경한 결과는 Fig. 6과 같다. 정상주의 경우에는 잎 내에 전분이 축적되지 않고 모두 전류된 상태로 보이고, 황화엽증 발생주의 잎은 통도조직 주변을 중심으로 전분이 많이 축적되어 있는 상태로 전류가 되지 않은 것으로 사료된다. 즉, 황화증상 개체의 잎은 바이러스의 전이를 막기 위하여 세포의 원형질 연락사(plasmodesmata)를 막아 광합성 산물이 전류되지 못하고 잎에 축적상태로 남아 있는 것으로 생각된다. 이것은 원형질 연락사 개폐에 관하여 바이러스 감염시 원형질 연락사가 닫힌다는 보고와 같은 맥락으로 판단된다(Epel, 2009; Lee et al., 2011; Lee, 2014).
멜론의 정상주와 황화엽증 발생주의 잎을 채취하여 무기성분을 분석한 결과는 Table 4와 같다. 모든 성분에서 황화엽이 정상엽보다 함량이 낮았고, 특히 마그네슘, 철분, 아연, 나트륨, 망간 성분 함량에 유의차가 큰 것으로 나타났다. 이와 같이 황화엽의 무기성분 함량이 정상엽보다 낮은 것은 황화증상의 원인이라기보다 황화엽 발생의 결과라고 추정된다(Takeshita, 2004). 이러한 결과를 토대로 최근에 국내의 멜론 재배지에서 급속하게 발생하고 있는 황화엽 증상은 생리적인 원인보다는 진딧물에 의한 바이러스 이병에 의한 원인이 더 큰 것으로 판단된다. 따라서 황화엽 증상 피해 확산을 줄이기 위해서는 바이러스 매개충인 진딧물을 사전에 방제하는 것이 좋을 것으로 판단되며 정확한 원인을 구명하기 위해서는 뿌리량, sink-source 크기에 따른 발생 등 다양한 각도에서 연구가 필요하다.
