

서 언
호접란(Phalaenopsis spp.)은 전세계적으로 가장 대중적인 난과 작목 중 하나이며 분화작목으로서 인기가 크게 높아지고 있다(Chen and Chen, 2011; Hou et al., 2011; Royal FloraHolland, 2018). 세계시장에서 대표적인 화훼 소비국인 미국의 2019년 난 거래량은 3천8백만본(USDA, 2020)으로 2010년 대비 43% 증가하였고, 거래액은 1.3억불 증가한 것을 통해 미국의 난 시장규모가 점차 커지고 있음을 알 수 있다. 특히 최근 미국에서 호접란 인기가 급증하면서 수요가 꾸준히 증가하여 대만과 네덜란드로부터 많은 묘가 수입되고 있다(aT, 2017). 미국에서는 일반적으로 개화가 가능한 성묘를 수입한 후 바로 저온처리를 통해 3–4개월 동안 개화를 유도하여 출하하는 시스템이다(Wang, 2007). 배지에 심겨진 성묘를 미국에 수출하기 위해서는 국가간 검역협상 타결이 필요하며, 국내에서는 2017년 12월 미국과 검역 협상이 타결되어 배지에 심겨진 호접란 묘의 수출 가능한 여건이 마련되었다(APQA, 2017).
화분에 심겨진 성묘는 상당한 부피를 차지하기 때문에 선박을 통해 해상으로 수출하는 것이 항공편을 이용하는 것보다 물류비를 절약할 수 있어 효과적이다(Hou et al., 2010; Cinantya et al., 2013). 그러나 선박 수송시 미국까지 운송기간이 2–4주이상 소요되며 이 과정에서 식물체는 광이 없고 수분관리가 되지 않는 등의 스트레스 환경에 놓이게 된다(Cinantya et al., 2013). Wang(2007)에 따르면 장기간의 암수송은 호접란 식물체 잎의 황화, 탈엽, 연부병 발생 등 묘 손실을 야기시키며 이는 수출 농가의 경제적 손실로 이어진다. 장기 선박 수송에 대응하여 수송과정에서 발생할 수 있는 부패, 잎 피해 증상 등을 줄이고 품질 감소를 최소화 할 수 있는 기술개발로 수출 분화의 상품성을 높일 필요가 있다(Fan et al., 2009).
수출묘의 품질에 영향을 미치는 여러 요인중 수송 중 80%이상의 습도는 식물체의 증산과 곰팡이 병발생에 영향을 미치는 주요한 요인이다(Ferrante et al., 2015). 호접란은 주로 수태배지에 식재하여 재배 및 수출되고 있으며, 수태는 보수력이 우수하여(Hwang and Jeong, 2007) 밀폐된 환경에서 수송시 수태의 높은 수분량이 수송 중 습도를 높여 잎의 황화와 병부 발생을 증가시킬 수 있다. 반면에 선적전 건조를 통해 배지내 수분량이 현저히 낮을 경우, 수송 기간 중 식물체와 배지의 증발산을 통한 수분 손실로 심각한 탈수 현상이 나타날 수 있으며 이는 분화의 품질과 상품성을 저하시키는 원인이 된다(Jones, 2002; Kim and Iersel, 2011). 따라서 이 연구에서는 수태 배지에 식재된 호접란 수출묘의 수송 중 손실 감소와 품질 유지를 위해 수송 전 배지 건조를 통한 수태 배지내 적정 수분함량을 정량화하고자 하였다.
재료 및 방법
실험 재료
1차 실험은 백색 화색의 대형종 Phalaenopsis ‘V3’와 분홍색 빅립 화형의 소형종 P. ‘Lovely Angel’ 품종을 충남 태안 소재의 농장(SangMi Orchids, Taean, Korea)에서 구입하여 실험 재료로 이용하였다. 이는 개화가 가능한 12개월묘로 ‘V3’ 품종은 평균 엽수 5–6장, ‘Lovely Angel’ 품종은 4–5장을 보유하였으며 두 품종의 평균 엽간거리(Leaf span)는 각각 46 ± 3cm, 30 ± 3cm였다. 직경 10cm (420mL)의 투명 비닐화분에 100% 수태(Sphagnum moss, Chille)배지로 식재한 식물체를 대상으로 하였다. 2차 실험은 1차 실험과 동일한 규격의 Phalaenopsis ‘V3’ 품종을 대상으로 하였다.
배지건조 및 모의암수송 처리
1차 실험은 국립원예특작과학원 유리온실(경도 °N35)에서 2018년 5월 4일부터 0(무처리), 1, 2, 3, 4주 동안 단수하면서 식물체가 심겨진 화분의 수태배지를 자연적으로 건조하였다. 이때의 온실내 평균 온도와 상대습도는 각각 25 ± 4/20 ± 1°C, 63 ± 14%였으며, 광도는 평균적으로 288µmol·m-2·s-1 PPF 수준이었다. 건조처리 완료 후 암조건 및 운송온도인 17°C하에서 20일동안 모의수송(Simulated dark shipping, SDS)을 실시하였으며 이때의 상대습도는 75 ± 2%였다. 모의수송 완료 후 냉방처리가 가능한 온실에 입실하여 품질조사를 실시하였고, 이 기간 동안 평균 온도와 상대습도는 22 ± 4/19 ± 2°C와 63 ± 12%였다. 관수는 모의수송 완료 5일 후부터 실시하였다.
수출시뮬레이션을 통한 실증을 위해 컨테이너를 이용하여 2차 실험을 진행하였다. 2018년 8월 22일부터 0(무처리), 1, 2, 3, 4주 동안 단수하면서 화분내 수태배지를 온실환경에서 자연적으로 건조시켰다. 이때의 온실내 평균 온도와 상대습도는 각각 26 ± 4/20 ± 4°C, 75 ± 15%였으며, 광도는 평균적으로 315µmol·m-2·s-1 PPF 수준이었다. 건조처리 기간 완료 후, 실제 수출용 종이박스[65cm(L) × 44cm(W) × 높이20cm(H), 통기공 상하면 12개]에 식물체를 14개씩 2단으로 포장하였으며, 처리당 3박스씩 20ft 냉장컨테이너에 적재하여 30일간 모의암수송을 실시하였다(Fig. 1). 컨테이너는 온도 18°C, 환기구 80%로 설정하였다. 이후 1차실험과 동일하게 저온실(평균 온도23 ± 4/19 ± 2°C, 상대습도 53 ± 12%)에 입실하여 모의수송 완료 5일 후부터 관수하였으며 품질조사를 실시하였다.

엽록소형광분석
엽록소형광 반응 측정을 위해 상위 두번째 잎을 30분간 암적응 시킨 상태에서 PAM 엽록소형광 측정기(PAM-2000, Heinz Walz, Effeltrich, Germany)를 사용하여 측정하였다. 처리별로 9개의 식물체를 임의로 선발하여 Quenching kinetics analysis를 통해 측정하였으며, 최소 형광값(Fo)과 최대형광값(Fm)을 유도하기 위한 광원은 각각 148µmol·m-2·s-1와 1,250µmol· m-2·s-1였다. 이로부터 ΦPSII(Quantum yield), qP(photochemical quenching), qN(on-photochemical quenching) 등의 형광변수를 산출하였다(Yoo et al., 2012). 최대양자수율인 Fv/Fm(maximum quantum efficiency)값은 Fv = Fm–Fo을 통해 산출하였다.
데이터 수집 및 통계 분석
수태 배지의 용적수분함량(Volumetric water content, VWC)을 측정하기위해 5TM 센서(Decagon Devices, Pullman, WA, USA)를 사용하였으며, 수태 배지의 함수량 변화에 따른 5TM센서의 출력값 변화를 파악하고 보정하였다. VWC는 화분당 두 곳에 꽂아 Procheck 휴대용 데이터로거(Decagon Devices, Pullman, WA, USA)를 통해 출력된 평균값을 사용하였다. 모의암수송 및 온실 재배시 환경은 호보(HOBO, Onset Computer, Co., USA)를 이용하여 온도와 상대습도를 측정하였다. 상대적엽록소함량은 SPAD meter(SPAD 502, Konica Minolta Sensing Inc., Sakai, Osaka, Japan)로 상위 두번째 잎을 측정하여 나타내었다. 꽃대수는 화분당 개수로 조사하였고 꽃대길이는 가장 긴 꽃대의 길이를 측정하여 평균길이로 나타내었으며, 꽃대당 꽃수 및 꽃의 길이와 폭을 조사하였다. 꽃눈출현소요일수와 개화소요일수는 모의수송완료 후 온실입실시기를 기준으로 꽃대의 길이가 0.5cm가 되는 시점과 첫 꽃이 완전히 개화한 날까지의 소요일수를 조사하여 평균일수로 나타내었다. 2차 실험에서는 모의암수송 완료 25일 후 황화로 인한 화분당 탈엽수를 조사하였다.
1차 실험의 경우 처리당 10개체가 되도록 완전임의배치하여 실험을 진행하였으며, 2차 실험은 처리별 4반복으로 각 반복당 28개체 배치하였다. 통계분석은 SAS 프로그램(SAS 9.4, SAS Institute Inc., USA)을 이용하여 분산분석(ANOVA)후, 5% 유의수준에서 처리 평균에 대해 Duncan의 다중범위검정으로 분석하였다.
결과 및 고찰
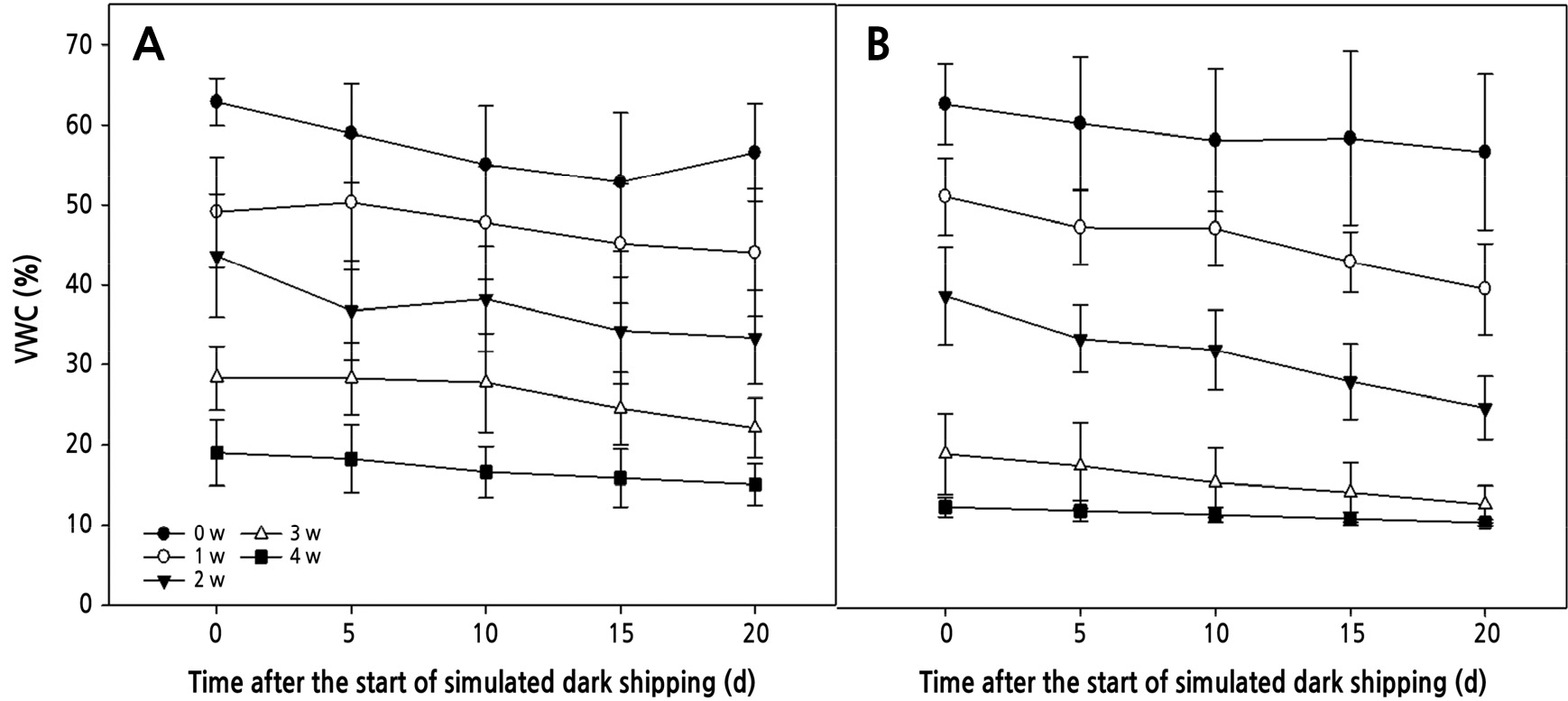
수태 배지의 용적수분함량(VWC)은 건조기간이 길어짐에 따라 감소하여 모의암수송 직전 VWC가 건조 무처리(0주)에서 두 품종 모두 63%였고, 1주에서 4주까지 건조기간이 길어짐에 따라 ‘V3’ 품종은 49, 44, 28, 19%로 감소하였으며, ‘Lovely Angel’은 51, 39, 19, 13%로 감소하였다(Fig. 2). 모의암수송 기간 동안 VWC가 점차 감소하는 경향을 보였고, 모의암수송이 끝난 직후 VWC는 처리별 건조수준에 따라 감소폭이 2–14%였으며, 4주 건조시 변화가 가장 미미하였다(Fig. 2).
상대적엽록소함량을 나타내는 SPAD값은 ‘V3’ 품종 0, 1주 건조처리에서 암수송 직전에 각각 57.5, 54.4였으나, 2주 이상 건조했을 때 50이하로 감소하였다(Fig. 3). 암수송 기간 동안에는 0, 1주 건조처리에서 값이 크게 감소하였고, 20일 암수송 후 온실에 다시 입실하였을 때 모든 처리에서 SPAD값이 증가하면서 회복하였다(Fig. 3). ‘Lovely Angel’ 품종에서도 0주 건조처리에서 ‘V3’ 품종과 동일하게 암수송 기간 동안 SPAD값이 큰 폭으로 감소하였다가 온실 입실 후 증가하는 것으로 나타났다(Fig. 3). 엽록소 함량의 감소는 잎이 황화되는 주요한 원인이 되며, 황화된 잎의 증가로 수출묘의 품질은 떨어지게 된다. Ferrante et al.(2015)에 따르면 엽록소의 합성경로는 광의 유무에 따라 조절되며 암조건은 엽록소의 분해를 촉진시키고 엽록소의 생합성 경로를 차단함으로써 잎의 황화를 촉진시킨다고 하였다. 또한 암조건에서 황화된 잎의 수가 증가하는 것은 영양생장을 통해 생성된 동화산물의 양이 감소하는 것과 관련이 있으며(Hou et al., 2010), 광이 부족한 상태에서는 호흡량이 증가하고 체내 당(sugar)이 소모되어 잎의 노화가 가속화된다(Ferrante and Reid, 2006).
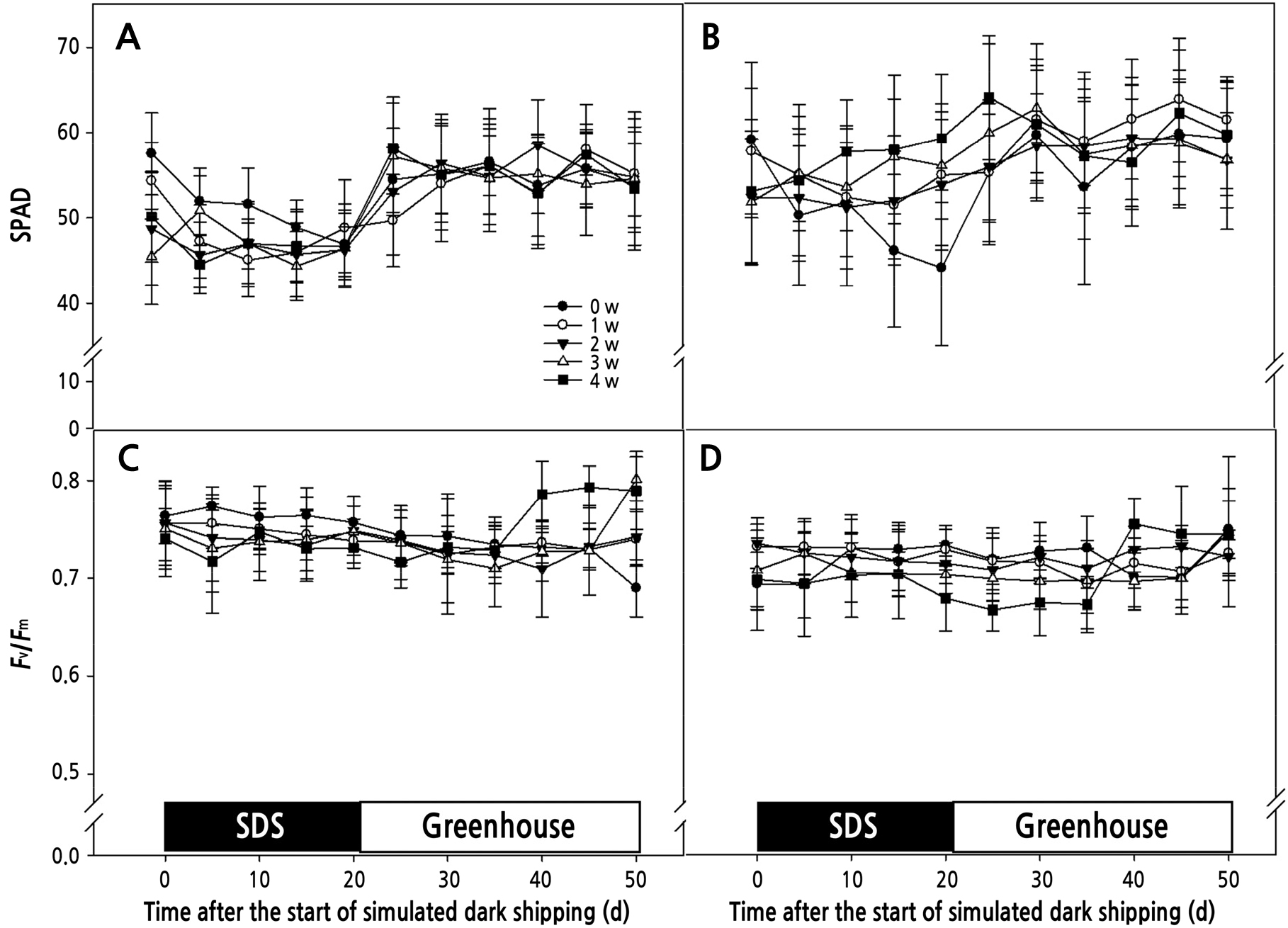
엽록소 형광 반응은 다양한 환경 조건에서 Phalaenopsis의 광합성 능력을 판단하는데 효과적인 것으로 알려져 있다(Lin and Hsu, 2004; Hsu, 2007; Lee et al., 2017). Fv/Fm값의 경우 암수송과 온실 입실 후 30일간 건조처리에 상관없이 ‘V3’ 품종은 0.69–0.80, ‘Lovely Angel’은 0.68–0.77범위로 유지되었으며(Fig. 3), 온실 입실 50일 후에도 비슷한 수준으로 품종간 차이만 있을 뿐 처리간에 유의적인 차이는 보이지 않았다(Table 1). 일반적으로 광계Ⅱ의 최대 광화학적 광합성 효율을 나타내는 Fv/Fm값은 0.7–0.8범위에서 식물체의 건전한 상태를 나타낸다(Bolhar-Nordenkampf et al., 1989). 따라서 Phalaenopsis의 경우 암조건이 처리에 상관없이 광계Ⅱ의 유의적인 손상을 일으키지 않는 것을 알 수 있으며, 이는 Hou et al.(2010)의 보고에서 21일간 암처리에도 무처리와 동일하게 Phalaenopsis가 0.8수준의 값을 가지며 건전한 광합성 효율을 유지한 것과 비슷한 결과였다. 또한 Dendrobium에서도 21일간의 암수송 전후 모두 Fv/Fm값이 0.77–0.83 수준으로 유지되는 것으로 나타났다(Cinantya et al., 2013). 온실 입실 50일 후 광계Ⅱ의 광합성능력을 판단하는 지표로써 ΦPSII(Quantum yield), qP (Photochemical quenching) 조사결과에서는 ‘V3’ 품종이 ‘Lovely Angel’ 품종보다 높은 경향을 보였으나 처리간의 유의적인 차이는 없었다(Table 1). 수송 후 손실정도를 판단하는 탈엽이나 부패증상은 1차 실험에서 처리에 상관없이 크게 발생하지 않았다. Phalaenopsis는 CAM(Crassulacean acid metabolism)형태의 광합성 기작을 보이는 식물(Endo and Ikusima, 1989; Guo and Lee, 2006)로써 비교적 두꺼운 잎과 뿌리를 가지고 있어 수분 유실 방지 능력이 있고 광에 대한 요구도가 낮아 장기 암수송 환경에 상당한 적응력을 가지고 있는 것으로 판단된다(Su et al., 2001).
Table 1.
Maximum quantum efficiency (Fv/Fm), actual quantum yields (ΦPSⅡ), photochemical quenching (qP), and non-photochemical quenching (qN) at upper spots of the leaves of Phalaenopsis ‘V3’ and ‘Lovely Angel’ at 50 days after simulated dark shipping. ΦPSⅡ, qP, and qN were measured with actinic light of 200 µmol·m-2·s-1 PPF
| Cultivar | Drying duration (weeks) | Fo | Fm | Fv/Fm | ΦPSⅡ | qP | qN |
| V3 | 0 | 0.419 bz | 2.20 b | 0.809 a | 0.450 a | 0.662 a | 0.562 a |
| 1 | 0.443 b | 2.25 b | 0.802 a | 0.412 ab | 0.612 ab | 0.625 a | |
| 2 | 0.465 b | 2.39 ab | 0.805 a | 0.475 a | 0.700 a | 0.575 a | |
| 3 | 0.459 b | 2.32 b | 0.802 a | 0.444 a | 0.655 a | 0.600 a | |
| 4 | 0.514 a | 2.53 a | 0.796 a | 0.366 b | 0.555 b | 0.622 a | |
| Lovely Angel | 0 | 0.485 a | 2.36 a | 0.794 a | 0.566 a | 0.811 a | 0.566 a |
| 1 | 0.494 a | 2.24 a | 0.779 a | 0.500 a | 0.800 a | 0.577 a | |
| 2 | 0.511 a | 2.30 a | 0.769 a | 0.500 a | 0.775 a | 0.525 a | |
| 3 | 0.497 a | 2.16 a | 0.773 a | 0.528 a | 0.814 a | 0.542 a | |
| 4 | 0.510 a | 2.26 a | 0.766 a | 0.533 a | 0.766 a | 0.500 a | |
| Significance | Cultivar (C) | * | NS | *** | *** | *** | NS |
| Treatment (T) | NS | NS | NS | NS | NS | NS | |
| (C) × (T) | NS | NS | NS | NS | NS | NS |
암수송 후 개화품질을 조사한 결과, 꽃눈출현소요일수는 두 품종 모두 0주 건조처리보다 건조시 촉진되는 경향을 보였다. 다만 ‘Lovely Angel’은 4주 건조처리에서 다른 건조처리구보다 꽃눈출현소요일수가 증가하는 것으로 나타났다(Table 2). 개화소요일수는 ‘V3’ 품종에서 3, 4주 건조처리시 0주 건조처리보다 10–13일 유의적으로 촉진되었고, ‘Lovely Angel’은 1–3주 건조처리시 0주 건조처리구보다 개화가 촉진되었다가 4주 건조처리에서 지연되는 것으로 나타났다(Table 2). 꽃대수는 ‘Lovely Angel’에서 건조시 수치적으로 증가하였으나 유의적인 차이는 없었으며, 꽃대길이, 꽃수의 경우 품종간 차이만 있을 뿐 처리간에 유의성은 나타나지 않았다(Table 2).
Table 2.
Subsequent flowering performance of Phalaenopsis ‘V3’ and ‘Lovely Angel’ depending on drying duration after simulated dark shipping
| Cultivar |
Drying duration (weeks) |
Days to inflorescence emergence |
Days to flowering |
No. of inflorescences |
Inflorescence length (cm) |
No. of flowers |
Flower size (cm) | |
| width | length | |||||||
| V3 | 0 | 55.0 az | 162 a | 1.000 a | 82 a | 8.4 a | 11.7 a | 9.8 a |
| 1 | 47.8 a | 156 ab | 1.000 a | 80 a | 8.4 a | 11.0 a | 9.1 ab | |
| 2 | 45.9 a | 156 ab | 1.000 a | 75 a | 7.6 a | 11.7 a | 8.9 b | |
| 3 | 47.0 a | 152 ab | 1.111 a | 79 a | 7.9 a | 10.9 a | 8.8 b | |
| 4 | 46.1 a | 149 b | 1.000 a | 76 a | 7.1 a | 11.7 a | 9.3 ab | |
|
Lovely Angel | 0 | 47.8 a | 133 a | 1.000 a | 42 a | 11.2 b | 6.2 a | 5.6 a |
| 1 | 38.3 b | 121 c | 1.000 a | 43 a | 11.8 ab | 6.2 a | 5.9 a | |
| 2 | 35.4 b | 123 bc | 1.125 a | 44 a | 11.8 ab | 6.2 a | 5.8 a | |
| 3 | 37.5 b | 127 abc | 1.167 a | 45 a | 11.8 ab | 6.1 a | 6.0 a | |
| 4 | 43.5 ab | 132 ab | 1.125 a | 46 a | 13.6 a | 6.1 a | 6.1 a | |
| Significance | Cultivar (C) | *** | *** | NS | *** | *** | *** | *** |
| Treatment (T) | ** | * | NS | NS | NS | * | * | |
| (C) x (T) | NS | NS | NS | NS | * | NS | NS | |
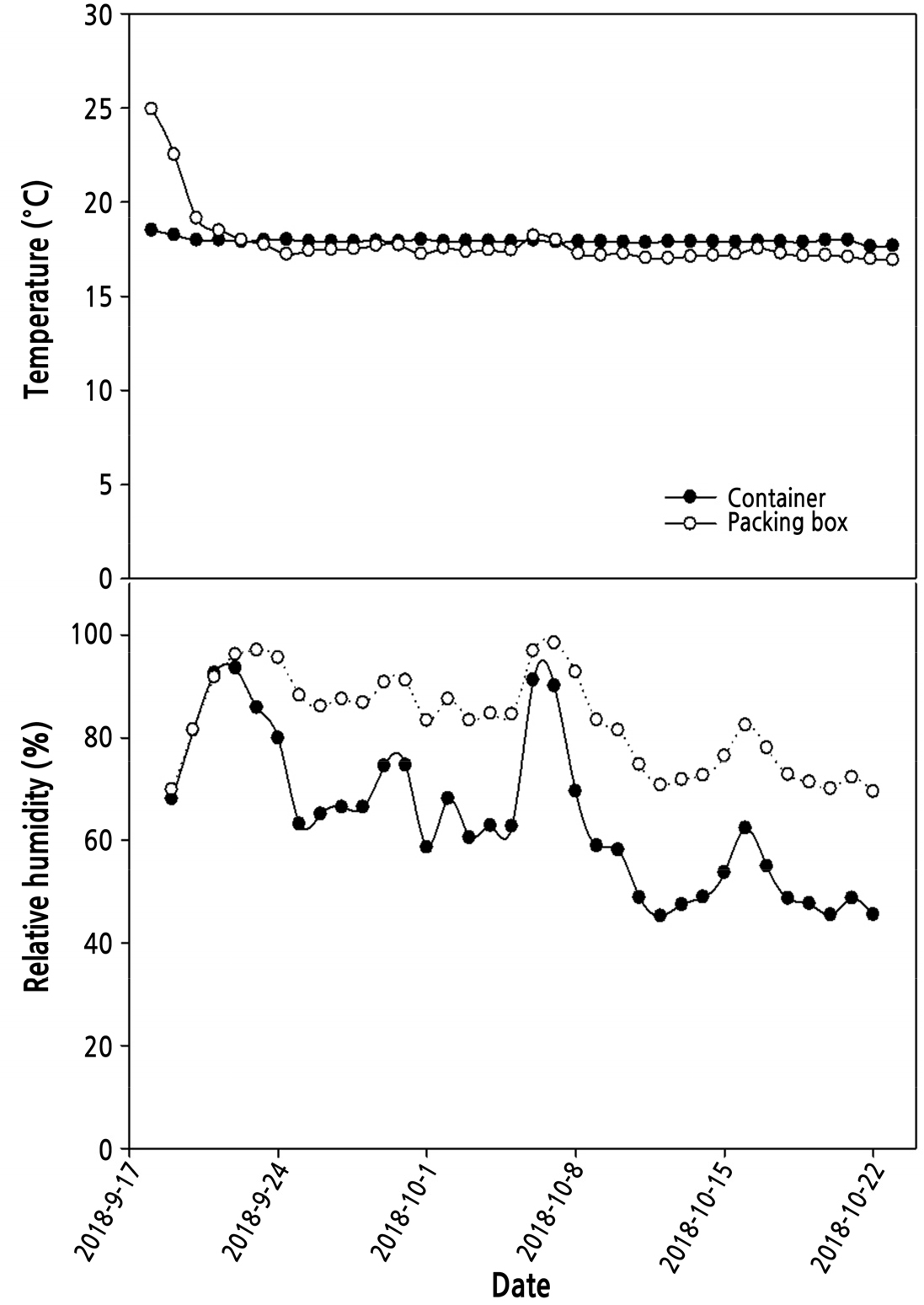
실제 수출환경과 비슷한 조건을 적용하고자 2차 실험을 통해 컨테이너를 사용하여 모의 수출시뮬레이션을 실시한 결과, 모의암수송 기간 동안 컨테이너 내부 온도는 설정온도인 18°C로 유지가 되었으나 포장박스내 온도가 컨테이너 설정온도까지 떨어지는데 2일 정도 소요되었다(Fig. 4). 실제로 종이상자에 식물체를 포장하여 컨테이너로 수출시 설정온도까지 도달하는데 3–4일 소요되는 것과 비슷한 결과였다(Chen and Chen, 2017). 상대습도는 암수송 기간 전반에 걸쳐 박스내 습도가 컨테이너내 습도보다 높게 유지되었으며, 평균 상대습도는 박스에서 83%, 컨테이너는 64%수준이었다(Fig. 4). 이는 식물체와 배지에서 보유하고 있는 수분이 박스내 습도를 높인것으로 판단되며, 수송 중 밀폐된 환경에서 높은 습도와 공기 순환의 장해는 부패 발생을 증가시키고 생리적 장해를 촉진시키는 주요한 원인이 된다(Ferrante et al., 2015). 운송 중 습도는 포장재와 컨테이너의 통풍량을 통해 조절할 수 있으며 수태의 수분량 조절을 통해 수태의 습도를 낮춤으로써 부패, 잎의 황화 등을 감소시킬 수 있다고 하였다(Chen and Chen, 2017).
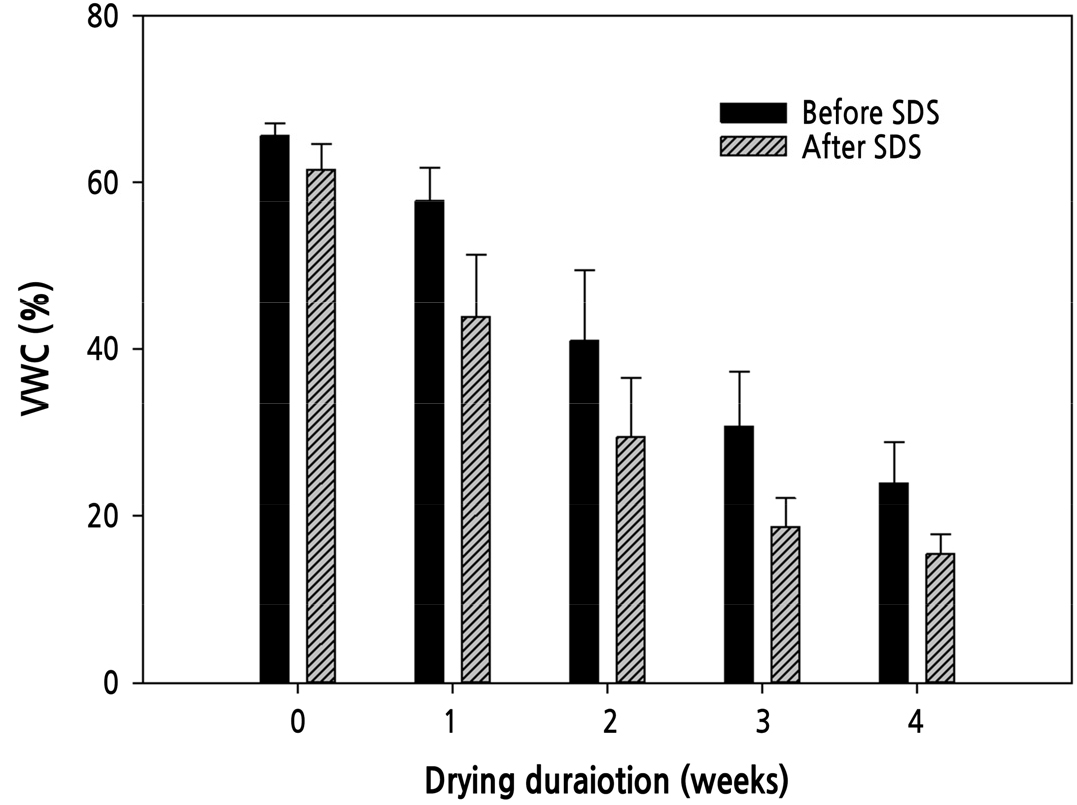
컨테이너 적재전 VWC는 0주 건조처리에서 65.6%였고, 건조기간이 증가함에 따라 VWC가 감소하여 1–4주까지 58, 41, 31, 24%로 감소하였다(Fig. 5). 30일간 모의암수송 경과 후 온실 입실시, 건조무처리(0주)에서는 62%로 배지내 많은 수분을 보유하고 있는 반면, 4주 건조처리에서는 15%까지 감소하였다(Fig. 5).
Fv/Fm값은 1차 실험과 비슷한 수준으로 0.7–0.8범위로 유지되었으며 처리간의 유의적인 차이는 없었다(Table 3). SPAD값은 암수송 직후 0주 건조처리보다 건조처리시 증가하였으며, 온실입실 25일 후 건조처리구에서 고도로 유의하게 엽록소 함량이 높게 나타났다(Table 3). Angelonia에서도 수송 전 배지의 수분함량을 낮게 조절하였을 때 배지의 수분함량이 높았을 때보다 잎의 SPAD값이 증가함으로써 더 진한 녹색잎을 가진다고 하였다(Jacobson et al., 2015). 분화의 일반적인 수확후 생리장해 중 하나가 잎의 황화이며 탈리되는 현상으로 이어지기도 하는데(Ferrante et al., 2015), 컨테이너에서 암수송이 끝난 25일 후 ‘V3’ 품종의 잎 기부가 황화되면서 탈엽수 증가로 이어졌다(Fig. 6). 0주 건조처리에서 탈엽수가 가장 많았으며, 건조처리를 통해 탈엽수가 점차 감소하여 VWC 31%의 3주 건조처리에서 가장 감소하였다. 암 수송 전 VWC 24%의 4주 건조처리에서는 오히려 탈엽수가 증가하는 경향을 보였다(Fig. 6). 이는 높은 강도의 수분스트레스가 오히려 잎의 노화를 촉진한 결과로 생각된다(Islam and Joyce, 2015). 암수송 후 꽃눈출현소요일수는 3, 4주 건조처리에서 0주 건조처리보다 3일, 개화소요일수는 4주 건조처리시 4일 촉진되었다. 꽃수는 3주 건조처리에서 가장 증가하는 것으로 나타났다(Table 4). Jacobson et al.(2015)에 따르면 분화 유통시 수송 전에 관수량을 줄임으로써 수송중의 암환경, 온도변화, 고농도의 에틸렌 발생 등 스트레스 환경에 대한 적응력을 높여 분화의 수명과 개화 품질을 높일 수 있다고 하였다. 장미, 샐비어 등에서 근권부의 수분함량을 감소시킨 식물체는 수분 이용 효율이 높아지고, 수분 소비량을 감소시킴으로써 건조 환경에서 세포의 팽압을 유지하여 수분 스트레스에 대한 저항성이 높아지는 것으로 나타났다(Williams et al., 2000; Nemali and van iersel, 2004). 건조 환경에서 세포의 팽압은 대사작용을 조절하는 주요한 요인이되며, 팽압이 감소할 경우 세포의 분열과 신장이 감소하고 식물의 생장과 발달에 관여하는 대사작용을 억제하는 것으로 보고된 바 있다(Hsiao, 1973).
Table 3.
The relative leaf chlorophyll content (SPAD) and maximum quantum efficiency (Fv/Fm) depending on drying duration in Phalaenopsis ‘V3’ at the end of simulated dark shipping (SDS) and 25 days after the end of SDS in a refrigerated container
|
Drying duration (weeks) | SPAD | Fv/Fm | |||
| at the end of SDS | 25 days after the end of SDS | at the end of SDS | 25 days after the end of SDS | ||
| 0 | 57.7 bz | 57.8 b | 0.810 a | 0.758 a | |
| 1 | 60.9 ab | 62.6 a | 0.807 a | 0.747 a | |
| 2 | 61.3 a | 62.5 a | 0.799 a | 0.750 a | |
| 3 | 60.7 ab | 62.7 a | 0.802 a | 0.737 a | |
| 4 | 63.5 a | 63.4 a | 0.805 a | 0.736 a | |
| Significance | * | *** | NS | NS | |
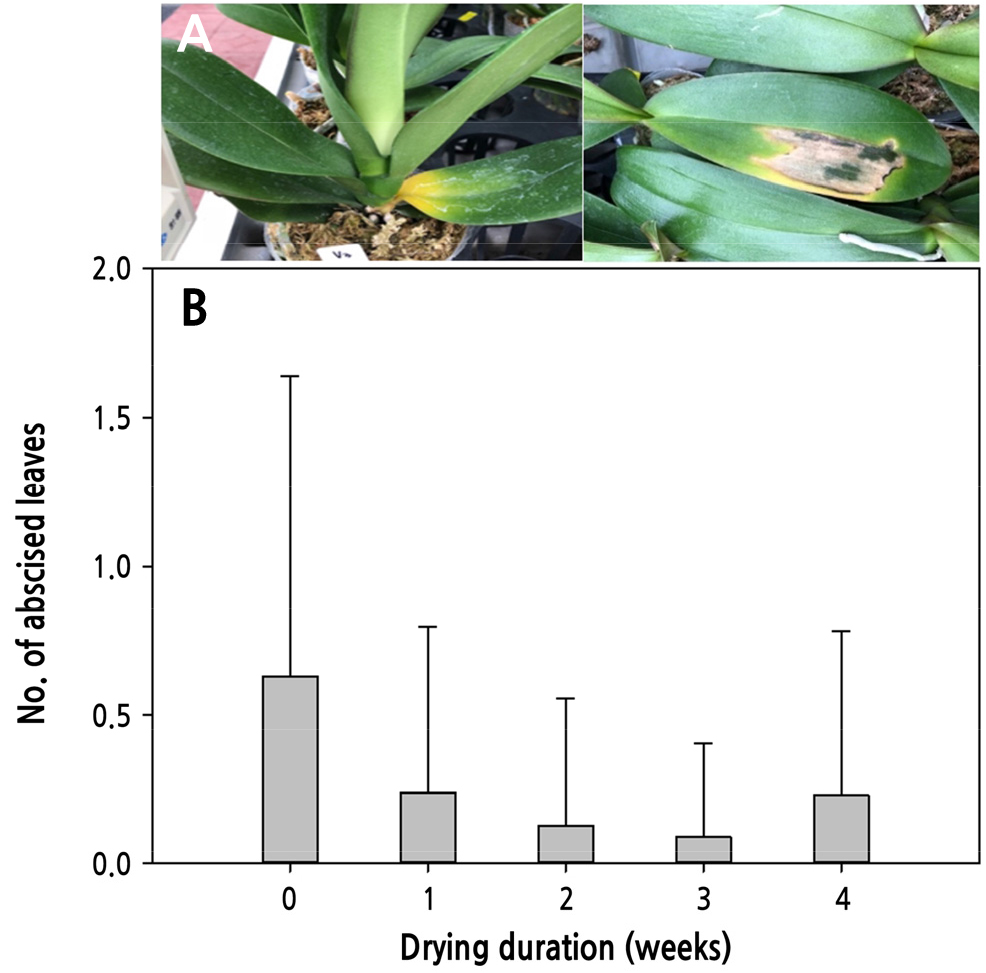
Table 4.
Subsequent flowering performance of Phalaenopsis ‘V3’ depending on drying duration after simulated dark shipping in a refrigerated container
|
Drying duration (weeks) |
Days to inflorescence emergence |
Days to flowering |
No. of inflorescences |
Inflorescence length (cm) | No. of flowers |
Flower size (cm) | |
| width | length | ||||||
| 0 | 37.8 az | 141.1 b | 1.20 b | 76 a | 8.13 bc | 12.6 a | 10.9 a |
| 1 | 37.8 a | 141.8 b | 1.06 b | 81 a | 8.93 ab | 12.8 a | 11.3 a |
| 2 | 38.0 a | 144.0 a | 1.50 a | 76 a | 7.93 c | 12.6 a | 11.1 a |
| 3 | 34.8 c | 142.1 b | 1.14 b | 81 a | 9.07 a | 12.7 a | 11.2 a |
| 4 | 36.3 b | 136.9 c | 1.00 b | 80 a | 8.67 abc | 12.7 a | 11.0 a |
| Significance | *** | *** | ** | NS | * | NS | NS |
종합적으로 Phalaenopsis는 광요구도가 낮고 높은 수분 보유능력을 가지고 있어 장기 암수송에 대한 적응력이 있는것으로 나타났다. 그러나 묘품질을 유지하기 위해서는 수태 배지의 건조를 통해 근권부의 수분량을 조절하여 수송 환경에서 마주하는 건조, 암환경 등의 스트레스 환경에 대한 저항성을 높이는 것이 필요하다. 건조기간은 온실 환경과 화분크기에 따라 상이하므로 수송 전 수태 배지의 VWC를 30%정도로 조절함으로써 잎의 황화와 탈엽이 증가하는 것을 막고 수출 후 환경에 대한 빠른 회복력으로 개화품질을 높일 수 있을 것이다.
