

서 언
재료 및 방법
실험 온실, 작물, 재배 관리
온실 환경, 광합성 요소 측정 및 식물체 엽온 모니터링
증산량 추정
통계분석
결과 및 고찰
온실 환경 변화
식물체 생장 변화
실험 기간 동안 파프리카 증산량 추정
서 언
파프리카(Capsicum annuum L.)는 2023년 기준 수출량은 총 21,700톤, 평년 수출량은 30,002톤으로 국내 총 생산량의 35–40%를 차지하고 있는 과채류의 대표적인 작물이다(KREI 2024). 파프리카는 수분 스트레스에 민감한 작물이며(Smittle et al. 1994), 식물체가 흡수한 수분의 약 1%만 대사활동에 이용되고 나머지 99%는 공기 중으로 증산한다(Salisbury and Ross 1992). 파프리카 재배 시 관수량이 적거나 증산작용이 지나치게 활발하여 배지 내 수분 부족 시 식물체 뿌리에서 이용할 수 있는 흡수량이 적어지고 이러한 상태는 과실 체적 생장과 총생산량에 영향을 미치기 때문에 적절한 관수 조절이 필요하다(De Swaef et al. 2012; Tai et al. 2014).
기존 증산량 추정 모델은 작물 표면에서의 순 일사량(MJ·m-2·day-1), 일평균기온(°C), 포화수증기압(kPa), 실제 수증기압(kPa), 상대습도 등의 환경 요인과 엽면적(지수)을 주로 이용하여 계산하였다(Monteith 1973; Allen et al. 1998). 이 중 엽면적은 식물 생장, 증산, 광합성 효율 등에 직접적으로 관여하는 요소 중 하나로 품질과 총생산량에 영향을 미친다(Heuvelink and Dorais 2005; Jo and Shin 2020). 엽면적을 측정하는 방법으로는 엽장, 엽폭을 조사하여 선형 회귀식으로 계산하는 간접적인 방법(Jolliet and Bailey 1992; Kage et al. 2000)과 식물체 전체를 파괴 조사하는 직접적인 방법이 있다. 하지만 기존 증산량 추정 모델은 노지 작물을 대상으로 하였으며, 엽면적은 별도의 회귀식을 만들어 활용했기 때문에 환경 요인과 생육 요인을 이용한 기존 추정 모델을 현장 적용 시 오차 발생 우려가 있다. 그리고 온실 내에서 수직형으로 재배하는 작물인 파프리카는 생장함에 따라 엽면적, 엽 수 등이 증가하며, 상부 엽 수의 증가로 인해 하부의 광량, 엽온 등 환경 요소는 상대적으로 감소한다. 이로 인한 증산량, 광합성, 양액 흡수량 등의 차이를 나타낸다(Kang et al. 2019; Woo et al. 2024).
따라서 본 연구는 실제 파프리카가 재배되고 있는 온실에서 한 작기 동안 엽온의 시계열 데이터를 대량으로 측정 및 수집하고 이를 이용한 일증산량을 추정하고자 수행되었다.
재료 및 방법
실험 온실, 작물, 재배 관리
본 실험은 파프리카를 재배 중인 전라북도 익산시 왕궁면에 위치한 벤로형 유리온실(N 35°97'16.7", E 127°10'36.9")에서 진행되었다(Fig. 1). 실험에 이용한 품종은 2022년 7월 20일 파종 후 8월 18일에 정식된 ‘나가노(Nagano RZ, Arseed, Korea)’였다. 배양액 조성은 농촌진흥청에서 제시한 파프리카 표준배양액(성분별 당량 표기)을 따랐으며, 누적 광량(100J·cm-2)을 기준으로1회 공급 시 약 90mL/dripper 내외로 하였다. 그리고 배양약의 pH 5.5–6.5, EC 2.5–3.5 dS·m-1 범위에서 생육 단계에 따라 조정하며 공급하였다. 사용 배지는 코이어배지(Daeyoug GS Co., Korea)로 정식한 다음 1차 분지 발생 후 2줄기만 유인하였다. 그리고 착색된 과실을 기준으로 약 2마디 아래 하위엽 전체를 적엽하며 관리하였고, 환경 관리는 일평균기온 평균 21.34 ± 1.80°C, 주간평균기온 22.80 ± 1.66°C로 농가의 관리에 따라 재배되었다(Table 1 and Fig. 3).

파프리카의 총 재배면적은 18,150m2(5,500평)이었으며, 이 중 실험에 이용된 적색 품종의 재배면적은 8,167m2(2,475평)로 재식밀도는 약 3.34주/m2였다.
온실 환경, 광합성 요소 측정 및 식물체 엽온 모니터링
환경데이터인 내부 기온, 외부 광량은 농가에 설치된 환경제어시스템을 사용하여 정식일인 2022년 8월 18일부터 2023년 6월 7일까지 수집된 데이터를 활용하였다. 그리고 식물체의 광합성 요소는 정식되어 있는 식물체 중 최소 4에서 최대 10반복으로 전체 잎을 대상으로 형광 측정기(LI-600, LI-COR, USA)를 이용하여 엽 주변 광량(태양복사량, Photosynthetic photon flux density, PPFD), 엽온(Leaf temperature, LT), 증산속도(Transpiration rate, TR)를 일주일 간격으로 측정하였다.
식물체 높이에 따른 엽온의 변화를 알아보기 위해 식물체 당 하부에 엽온 센서(LT-1T-V2, Bio instruments S.R.L, Republic of Moldova) 1개씩을 분지점 위 3마디(약 90cm) 위치에 고정하였고, 분지점 위 생장점 아래 중간 부위에 센서 1개씩을 설치하였다(Fig. 2). 그리고 식물체가 수직형으로 생장하면서 발생하는 그림자로 인해 모니터링 센서를 식물체 중간에 위치할 수 있게 일정 간격을 주어 이동해주었다. 실험 기간 동안 센서를 이동시킨 횟수는 총 4번이었다.

증산량 추정
실험 기간 동안 일증산량을 계산하기 위하여 Python(version 3.12.4, Python Software Foundation, USA)의 pandas, numpy 라이브러리를 이용하여 아래의 순서에 따라 산출하였다. ① 매주 측정된 식물체의 재식 위치, 높이별로 측정된 엽온(독립변인)과 증산량(종속변인) 간 회귀모형(EQTR, p < 0.05)을 작성하였다. ② 도출된 EQTR에 2분 간격으로 모니터링된 엽온을 대입하여 2분 간격의 증산량(mmol·m-2·s-1)을 도출한 후 120초(2분)를 곱하여 2분 간 증산량을 TR1(mmol·m-2·2minutes-1)으로 나타내었다. ③ ‘②’의 데이터를 일중 증산량 변화 회귀모형(EQDTR)을 적용하여 TR2를 계산하였다. ④ EQDTR에 의해 도출된 데이터를 일별로 누적한 후 1,000(mmol → mol)으로 나누어 일증산량(DATR, mol·m-2·day-1)을 도출하였다. ⑤ DATR 데이터를 약 일주일 간격으로 측정한 정식 후 일수에 따른 식물체 당 엽면적 변화 회귀식(EQDLA, p < 0.05)을 적용하여 식물체 당 일증산량, TR3(DPTR, mol·plant-1·day-1)으로 계산하였다. ⑥ DPTR 값에 순수한 물 1mole = 18g을 적용하여 g·plant-1·day-1 단위로 TR4를 계산하였다.
일중 증산량 변화를 알아보기 위해서 오전 5시부터 오후 5시까지 매 시각마다 식물체 8주를 대상으로 상부, 중부, 하부 3잎을 측정하였다. 그리고 측정된 데이터를 시각별로 평균하고 최대값을 나타낸 시각의 값을 1(100%)로 하여 다른 시각 평균값을 상대 값으로 변환한 후 일중 변화 회귀모형을 작성하였다.
통계분석
집단 간 평균 차이 검증은 95% 신뢰수준(p < 0.05)에서 F-검정을 통한 이분산(Heteroskedasticity test) 또는 등분산(Homoskedasticity test) T-검정을 실시하였다. 주요 환경 요소와 생리 요소 간 관계 분석인 회귀 분석(Regression analysis)은 SPSS 프로그램(Version 19.0 SPSS Inc., Chicago, USA)을 이용하였다. 그리고 회귀 그래프는 Excel 2018(office 365, microsoft Crop., Canada)을 이용하여 작성하였다.
결과 및 고찰
온실 환경 변화
온실 내부의 일평균기온과 외부의 일평균광량의 계절적 영향을 확인하기 위해 월별로 평균한 결과(Table 1), 재배 기간 동안 내부 일평균기온은 19.4–24.5°C로 관리되었고, 외부 일평균광량은 802.5–2,446.2J·m-2였다. 그리고 일평균기온은 평균 변이계수(Coefficient of Variation, CV)가 4.9%로 2022년 8,9,12월과 2023년 4월에 높았다. 일평균광량은 평균 CV가 38.10%로 2022년 8월, 2023년 4–5월에 높았는데 이는 흐린 날과 맑은 날 수가 고르게 분포되었기 때문이었다(KMA 2023). 온실 내부의 기온은 식물체 온도에 직접적인 영향을 미치고, 온실 내부로 들어오는 직달 광은 식물체 온도를 높일 수 있다(Park 2011). 따라서 이러한 월별 경향은 식물체 엽온에 영향을 줄 것으로 생각된다.
재배 기간 동안 온실 내부의 주간평균기온과 주간평균엽온의 변화는 Fig. 3과 같다. 두 온도 요소의 수준은 각각 22.80 ± 1.66°C와 21.63 ± 0.98°C로 기온이 엽온에 비해 약 1.17°C 높았다. 그리고 실험 기간 동안 변화 패턴은 유사한 경향으로 두 요소 간 상관성(Blonder and Michaletz 2018)에서 나타난 것으로 판단된다. 하지만 변화 폭은 엽온보다 기온에서 더 컸다. 일반적으로 광도가 높아짐에 따라 기온과 엽온은 모두 상승하고 엽온이 기온보다 높아진다고 하였는데(Cho and Kwack 1994) 본 실험 기간에서는 반대의 경향을 나타내었다. 이러한 차이는 선행 연구에서는 주간과 야간 전체 시간의 일평균값을 비교하였으나, 본 연구에서는 야간이 제외된 주간 동안의 평균값을 비교하였기 때문이다. 하지만 일평균기온과 일평균엽온을 사용하였을 땐 선행 연구와 같은 결과를 나타내었다(자료 미제시).
Table 1.
Daily inside temperatures and outside solar radiation levels per month in the Venlo-type greenhouse from August of 2022 to June of 2023

식물체 생장 변화
실험 기간 동안 정식 후 일수에 따른 줄기 당 마디수 및 엽수, 식물체 당 엽면적 변화를 회귀분석하였다(Fig. 4). 최종적으로 줄기 당 마디수는 50 nodes/stem였고 엽수는 142 leaves/stem, 그리고 식물체 당 엽면적은 1.53m2/plant였다. 정식 후 일수에 따라 줄기 당 마디수의 증가 속도는 0.18 node/stem/day로 평균 5일에 1마디가 발생하였다. 엽수는 0.5268 leaf/stem/day로 평균 2일에 1엽이 발생하였고, 엽면적은 0.0025m2/plant/day로 평균 40일에 식물체(2줄기) 당 0.1m2씩 증가하였다.

실험 기간 동안 파프리카 증산량 추정
광에 의해 내부 고온 발생 시에는 온도 관리를 위해 차광 스크린을 이용한다. 그리고 운량이 매우 많아 광량이 시각별 큰 차이를 나타내지 않는 날에도 일중 증산량의 최소값과 최대값의 차이는 낮아지는 경향이었다(자료 미제시). 생리적으로는 기공 열림에는 적색광과 청색광이 1차적 신호로서 관여한다(Lurie 1978). 하지만 광에 의해 촉발된 기공열림 이후 증산량에는 수증기압포차가 좌우되며(Wilson and Bunce 1997; Tai et al. 2010) 수증기압포차 증가는 증산 촉진을 가져와 엽온을 저하시킨다(Idso 1982). 이는 내부 광량은 재배 관리에서 변동성이 매우 크지만 엽온은 광량과 기온의 영향을 모두 내포하고 있다고 판단하여 본 연구에서 엽온을 독립변인으로 선택하였다.
실험 기간 동안 매주 파프리카 식물체 분지점부터 상부 방향으로 형광 측정기를 이용해 측정한 엽온과 증산속도를 동일한 위치 잎마다 평균한 값을 이용하여 회귀분석하였다(Fig. 5). 수직형으로 생장하는 파프리카의 특성 상 엽에 직달하는 광량이 많은 상부에 가까이 갈수록 엽온 또한 높아지는데, 엽의 반복수가 적어지면서 28°C를 기준으로 고온으로 갈수록 증산속도의 편차가 커지는 경향이었다. 회귀분석 결과, 엽온과 증산속도 간 지수함수로 도출하였고 R2값이 0.77(p < 0.01)로 유의하게 나타났다.

Fig. 5.
Regression analysis between the leaf temperature and transpiration rate of paprika leaves in the Venlo-type greenhouse from August of 2022 to June of 2023. Dots are the average values of data measured every week on leaves at the same location above the branch point during the experimental period. ***Significant at p < 0.001.
잠열 에너지의 근원인 일사량은 식물체 표면의 온도를 증가시켜 기온과의 차이를 발생시키는데 이에 따른 증산량은 식물체 표면 수증기압과 공기 중 수증기압 차이에 비례해 일어난다(Monteith 1973). 일사량에 의한 상승된 엽온은 증산에 의한 잠열 손실로 다시 엽온이 내려간다. 이는 외부로부터 유입되는 직달 광에 의해 엽온이 상승할 시 잎과 대기 간 수분포텐셜 차이로 인해 잎 내부의 수분이 대기로 유출(증산)되기 때문이다(Bange 1953; Idso 1982). 재배 과정 중 작물에 흡수된 수분의 약 96%는 증산 잠열로 대기 중에 방출되어 식물체 내 열에너지 소실 반응에 이용된다는 결과도 있다(Seo et al. 2019). 결국 대기환경요소는 엽온의 변화를 주고 이후 엽온은 증산량에 영향을 미치는 것으로 생각된다.
온실에서 내부 기온을 환경제어를 통해 일정하게 유지한다고 가정하여도 기공 개방과 폐쇄는 일어나기 때문에 엽온으로 증산량을 추정한 회귀모형에 일중 증산량 변화를 적용하기 위하여 일중 증산량 회귀식을 도출하였다(Fig. 6). 그리고 일중 증산량의 최대값을 나타낸 특정 시각을 1(100%)로 하여 다른 시각의 값을 이에 대한 백분율로 나타내었다. 일중 증산량의 최대값은 13시경에 측정되었고, 5시경(6월) 일출 후 8시에서 10시까지는 급격한 증가, 10–13시에 최대, 13시 이후부터 급격한 감소되는 경향이었고, 10시에서 13시 사이에는 유의한 차이를 나타내지 않았다. 13시경의 증산량을 최대값으로 하여 다른 시각을 상대값으로 변환하였을 때 일출에서 9시에는 0.25–0.68, 그리고 14시부터 일몰까지는 시간에 경과에 따라 0.58–0.00이었다. 도출된 상대값을 가지고 회귀분석한 결과 3차 다항식으로 도출되었다. 회귀식에 따르면 최대값은 오전 10시 15분 정도로 나타났다. 일반적으로 일출 후 상승하는 광량은 12–14시까지 증가한 후 일몰까지 감소하는 형태인데, 이를 고려하면 광량과 엽온(Cho and Kwack 1994; Tai et al. 2010; Nam et al. 2017), 엽온과 증산량(Bange 1953)은 정의 상관성을 나타내었다는 선행연구와 동일한 결과로 나타났다. 하지만 일중 변화는 외기 기온이 구분되는 계절적 요인(Table 1)이 작용할 것이며, 일중 증산량의 변화도 시기별 구분을 통해 별도 모형을 작성함과 함께 하루를 증가시간, 유지시간, 감소시간 3 구간으로 나누어 모형을 작성하고 적용한다면 더욱 정확한 결과를 도출할 수 있을 것으로 판단된다.

엽온에 따른 증산량 회귀식(Fig. 5)에 실험 기간 동안 2분 마다 모니터링된 엽온을 입력하여 계산하고 이에 120초(2분)를 곱한 후 일중 증산량 변화(mol·m-2·day-1, TR1)에서 도출된 회귀식(Fig. 6)을 적용하여 일증산량(mol·m-2·day-1, TR2)을 계산하여 월별 평균하였다(Table 2). TR1은 전체 기간 동안 최소 1.98mol·m-2·day-1, 최대 249.76mol·m-2·day-1, 평균 36.38 ± 32.87mol·m-2·day-1였다. 그리고 TR2는 최소 1.11mol·m-2·day-1, 131.16mol·m-2·day-1, 평균 20.44 ± 18.27mol·m-2·day-1로 일중 변화 회귀식을 적용한 TR2보다 적용하지 않은 TR1값보다 평균 15.95 ± 14.60mol·m-2·day-1가 낮아졌다. 특히, TR1, TR2 모두 전체 일수 중 6일이 인근 날에 비해 매우 높은 값을 나타내었다.
Table 2.
Comparison of the average production level among the monthly transpiration rates of paprika plants per month during the experimental period in the Venlo-type greenhouse from August of 2022 to June of 2023
시간의 경과와 함께 식물체는 생장함에 따라서 엽면적이 증가한다. 따라서 Table 2의 TR2의 단위면적(1m2) 당 계산된 값에 엽면적의 변화 회귀모형(Fig. 4)을 적용시켜 식물체 당 일증산량(mol·m-2·day-1)을 계산하였다. 계산한 결과(Table 2, TR3), 식물체 당 일증산량의 평균값은 26.04 ± 23.98mol·m-2·day-1였다. 그리고 가장 낮은 날은 1.55mol·m-2·day-1였고 가장 높은 날은 146mol·m-2·day-1 수준으로 최소값과 최대값의 차이가 매우 컸다.
TR3 값에 순수한 물(H2O) 1mole 무게인 18g을 곱하여 일중 식물체 당 증산된 물의 무게(g·plant-1·day-1)를 나타내었다(Table 3). 증산된 물의 무게는 평균 468.7g·plant-1·day-1, 편차 431.58g·plant-1·day-1이었고, 가장 낮은 날은 27.93g·plant-1·day-1, 가장 높은 날은 2,627.94g·plant-1·day-1수준을 나타내었다. TR4 값을 월별로 평균하였을 때, 식물체가 생장함에 따라 엽수의 증가로 식물체 당 TR4값은 지속적으로 증가하는 경향이었다. 하지만 2023년 4월에는 값이 일시적으로 감소하였는데 이는 온실 내부 기온과 엽온이 낮아 엽온에 따른 증산량 회귀모형(Fig. 5)에 적용할 시 엽면적 증가 대비 엽온의 상대적 영향이 커 값이 낮아졌기 때문이다.
Table 3.
Daily estimated weight of transpired water (EDWTW) of paprika plant per month during experiment in Venlo-type greenhouse from August 2022 to June 2023
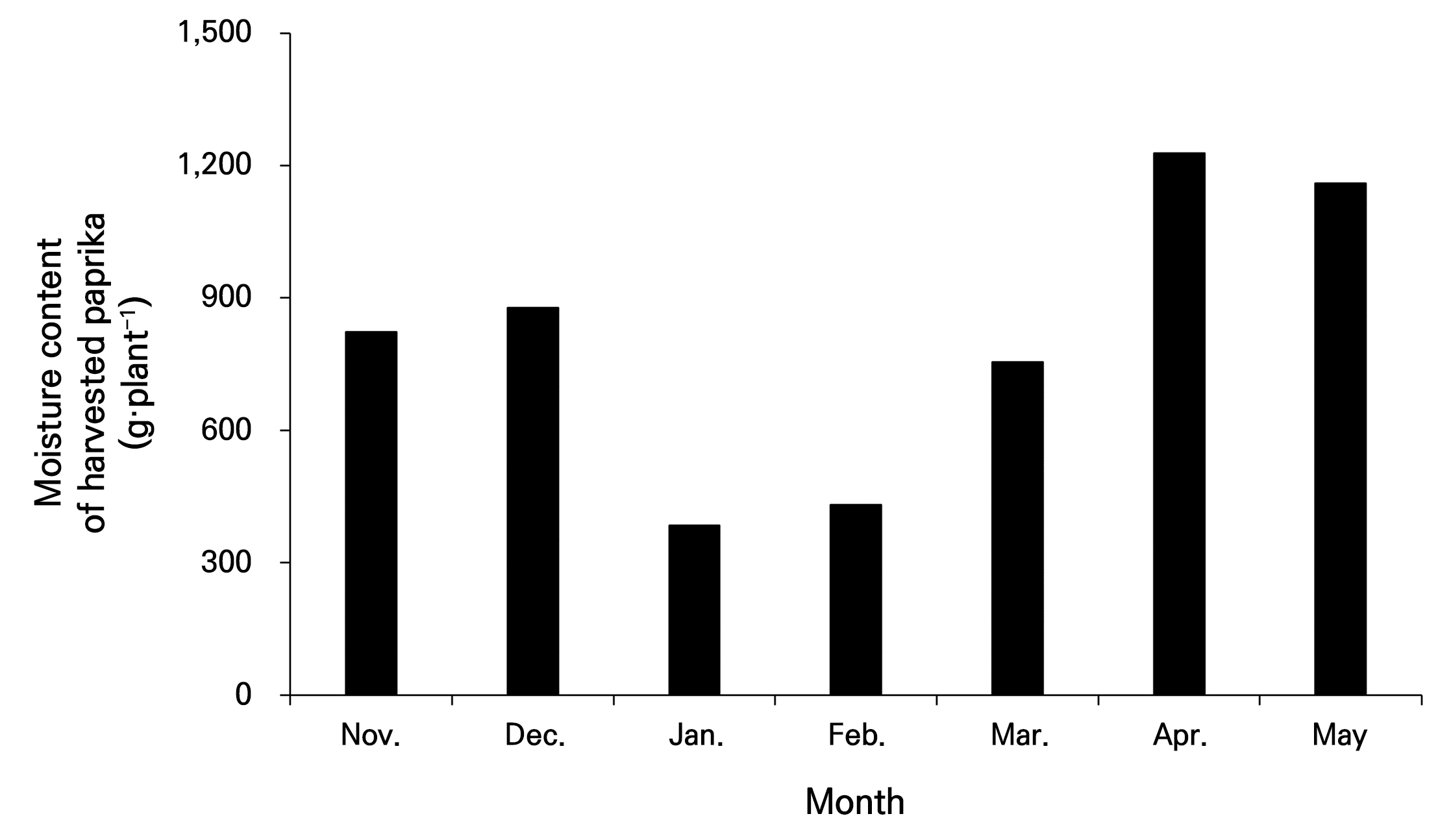
TR 4과 수확 과실의 수분함량과 실제 온실의 작물 흡수량 간 차이를 비교하였다(Fig. 7). 작물 흡수량은 재배 농가에서 관행적으로 계산되는 공급량에서 배액량을 뺀 값으로 비교하였다. 그리고 배액량은 일괄적으로 공급량의 30%로 간주하였다. 식물체 당 일증산량(g·plant-1·day-1)은 TR4에서 작물 흡수량보다 최대 627.97g·plant-1·day-1적게 나타났다. 이러한 차이 정도는 파프리카 작물의 수확량 패턴과 유사한 경향으로 나타났다. 본 실험 기간 동안 온실에서 수확된 작물의 수분 함량을 나타낸 결과(Fig. 8), 실험 온실의 수확 패턴인 11월과 12월 수확량의 증가, 1월에서 2월 수확량 감소, 이후부터 급격히 증가하여 최대 수확량, 5월부터 작기 마무리 시기인 6월까지 감소하는 경향을 따라 수확된 과실의 수분함량 또한 같은 경향을 나타냈다.

Fig. 7.
Changes in the daily absorption of nutrient solution (DANS) and the estimated daily weight of transpired water (EDWTW, Table 3) of paprika plants per month during the experimental period in the Venlo-type greenhouse from August of 2022 to June of 2023.
작물의 양액 흡수량(Accumulated daily absorption of nutrient solution, ADANS)과 수확 과실의 수분함량을 포함한 추정 증산량(Accumulated estimated transpiration including the moisture content of harvested paprika, AETMC)을 누적한 결과(Fig. 9), 최종값은 각각 107,781.1, 10,3148.4g·plant-1·day-1로 ADANS가 AETMC보다 약 4,632.7g·plant-1·day-1로 많았으며, 증가속도는 각각 508.55, 471.31g·plant-1·day-1으로 ADANS가 AETMC보다 약 37.24g·plant-1·day-1빨랐다. 그리고 정식 후 약 227일 후부터 ADANS와 AETMC의 차이는 더 크게 발생하였는데 이는 수확량이 증가하는 4월이지만 수확 과실의 수분함량이 정확하게 측정된 값이 아닌 계산한 것에서 발생하는 오류로 생각된다. 이를 고려할 때 작물의 흡수량과 추정된 증산량과의 차이에는 과실 수확량의 수분량이 가장 크게 관여하고, 이외에 작물의 단위시간 당 생장 줄기 및 엽의 수분량, 그리고 실제 배액량과 본 연구에서 일괄 적용한 공급량의 30% 간 차이, 적심·엽의 양 등이 관여한 것으로 생각된다.

Fig. 9.
Changes in the accumulated values of the accumulated daily absorption of nutrient solution (ADANS) and the accumulated estimated transpiration including the moisture content of harvested paprika (AETMC) during the experimental period in the Venlo-type greenhouse from August of 2022 to June of 2023.
최근 에너지와 관수 측면에서 작물 증산량 연구가 다수 진행되어 오고 있다. 하지만 실제 파프리카의 재배 전주기 동안 실측값을 통한 연구는 없는 상태이다. 본 연구는 이러한 대량 데이터의 확보를 통해 추정한 것에 의미를 갖고 있다. 시기별 과실 수확량, 생장량, 적심·엽의 양, 그리고 실측 배액량, 배지의 수분함량, 시기 및 높이별로 더욱 세분화하여 엽온과 증산량 간, 일중 증산량 변화 등의 회귀식을 도출하여 적용한다면 더욱 정확한 증산량 추정이 가능할 것으로 생각된다.
