

Introduction
Materials and Methods
Plant Materials and Culture Systems
MeJA Treatment
Growth Measurements
Extraction and Analysis of Tilianin and Acacetin by HPLC
Statistical Analysis
Results and Discussion
Effect of EC on Plant Growth
Effect of EC on Tilianin and Acacetin Contents
Combined Effect of MeJA and EC on Phenylpropanoid Levels
Introduction
Agastache rugosa, a species of Korean mint in the family Lamiaceae, is a short-lived, perennial, aromatic herb with light-green foliage comprised of stiff, toothed leaves and showy, purple floral spikes. A. rugosa is distributed throughout East Asia, including China, Japan, Korea, and Siberia, and is used in traditional medicine to treat numerous illnesses, such as bacterial infections, cholera, miasma, nausea, and anxiety (Park et al., 2016). Leaves are commonly consumed in meat marinades, tea, and salad dressings (Gong et al., 2012). A. rugosa produces various secondary metabolites such as flavonoids (Hernandez-Abreu et al., 2013), essential oils (Skakovskii et al., 2010), non-volatile terpenoids and sterols (Estrada-Reyes et al., 2004), lignans (Lee et al., 2002), and phenolic acids (Janicsak et al., 1999). These metabolites have numerous pharmacological and physiological properties, including anti-wrinkle (Kim et al., 2015), anti-viral (Wang et al., 2009), cardiovascular (Hernandez-Abreu et al., 2013), anti-atherogenic (Hong et al., 2001), antimicrobial (Shin, 2004), antioxidant (Guo et al., 2011), and anti-inflammatory effects (Nam et al., 2005), as well as anti-fungal (Shin and Kang, 2003), anti-melanogenesis (Kim et al., 2017), and antibiofilm-antitumor activities (Haiyan et al., 2016).
A. rugosa has gained attention due to its potential food, cosmetic, and medicinal values. Its large-scale cultivation was initiated to supply nectar to honeybees, and to obtain essential oils and phytochemicals for the flavor, perfume, and food industries (Fuentes-Granados, 1997). Not only is the plant itself used in traditional medicine, but plant extracts are in high demand. A. rugosa is cultivated for industrial use, but harvest is limited to twice a year in open fields. The composition and quantity of functional substances can be inconsistent due to seasonal variation in the cultivation environment and seasonal changes in the quality of the plantation (Rao and Ravishankar, 2002). Thus, mass cultivation methods and techniques for isolating its constituent chemicals are needed. Soil-based culture requires arable land, abundant water, fertilizers, and pesticides to achieve high yields. Water is an economically scarce resource, particularly in arid and semi-arid zones. Thus, traditional agricultural methods are being replaced with soilless cultivation systems that use water with greater efficiency. Hydroponic cultivation under well-managed, controlled climatic and growth conditions in nutrient solution has successfully been used to produce leafy vegetables such as radish (Salerno et al., 2005), lettuce, and lamb lettuce (Zanin et al., 2011). Hydroponic cultivation provides multifold advantages compared to traditional cultivation, including low labor cost, precise control of plant nutrition and light intensity, reduced use of pesticides, fewer effects of seasonality (Paulus et al., 2012), improved yield (Nicola et al., 2005), increased phytochemical concentrations (Wu and Kubota, 2008), year-round production in areas with short cultivation cycles, and high-quality, high-density cultivation of plants that are free of weeds and soil particles.
Plants emit airborne signals when challenged by abiotic stresses, such as adverse weather, salinity, drought, and low or high temperature, and by biotic stresses such as herbivores (Pare, 1999) and pathogens. These signals allow plants to communicate with other members of the species in their surroundings, attract specific pollinators, and reduce their susceptibility to harmful insects (Pichersky and Gershenzon, 2002). Methyl jasmonate (MeJA) is a volatile fragrant compound that was first reported in Jasminum grandiflorum flowers and is distributed ubiquitously in the plant kingdom. Jasmonates, such as MeJA and its free-acid jasmonic acid, are cellular regulators involved in various developmental processes, including senescence, seed germination, and fruit ripening (Creelman and Rao, 2002). Jasmonates activate plant defense mechanisms in response to wounding caused by insects (Wasternack and Parthier, 1997). In addition, jasmonates induce the accumulation of various secondary metabolites (Afrin et al., 2015). Some secondary metabolites from A. rugosa such as flavonoids, particularly the phenylpropanoids tilianin and acacetin, are potent anti-atherogenic compounds, which prevent narrowing of the arteries caused by plaque build-up (Hong et al., 2001). Atherosclerosis adversely affects the kidneys and causes peripheral artery and coronary artery disease and stroke.
No studies have examined the combined effect of applying MeJA and using nutrient solution with a high electrical conductivity (EC) on hydroponically grown A. rugosa plants. Thus, we evaluated the influence of the EC of the nutrient solution and fragrant MeJA on secondary metabolite production in A. rugosa. We determined the optimum EC value, MeJA concentration, and quantities of the flavonoids, acacetin, and tilianin, accumulated during hydroponic cultivation.
Materials and Methods
Plant Materials and Culture Systems
A. rugosa seeds were purchased from Danong Seed Co., Ltd. (Seoul, Korea) and sown in a crushed rook-wool media tray (240 holes, UR Rockwool Co., Ltd., Suwon, Korea). The trays were placed in a culture container (1,200 mm long × 7,000 mm wide × 110 mm high) for germination and raising seedlings. Three culture containers (1,200 mm long × 7,000 mm wide × 110 mm high) were installed in a module with three lighting systems. The distance between the container and the lighting system was 600 mm. Three fully-equipped modules were set up in a plant culture room with dimensions of 5 m length × 3 m width × 4.5 m height, maintained at 22±1°C (day) and 18±1°C (night) using an air conditioner (AF30FSAM1EEN, Samsung Co., Ltd., Suwon, Korea). Fluorescent lamps (TL5 14W/865 Philips, Amsterdam, Netherlands) were installed above the culture container to provide a photosynthetic photon flux density (PPFD) of 120± 10 µmol·m-2·s-1 for raising seedlings and 180±5 µmol·m-2·s-1 for growing plants to the bottom of the tray. These PPFDs were averages measured at nine points above the canopy level of the container. The photoperiod was 16-h/8-h for 4 weeks. After the appearance of the hypocotyl, half-strength Otsuka House 1 nutrient solution (OHNS) [16.8 NO3-N, 1.8 NH4-N, 8.2 Ca, 5.1 PO4-P, 3.0 Mg, and 8.6 K (me·L-1) containing several micronutrients] was provided during the seedling growth stage. Ninety-six 35-day-old seedlings (four-leaf stage) were selected for EC treatments with OHNS at three ECs: 1.0, 2.0, and 4.0 dS·m-1.Thirty-two seedlings for each treatment were transplanted into the culture container at a planting distance of 120 mm in a deep flow technique culture system. The nutrient solution was circulated intermittently for 20 min at intervals of 10 min for 4 weeks.
MeJA Treatment
Three culture-containers previously used for EC treatment were partitioned into four compartments with white board papers. Eight 3-week-old plants were placed into each compartment for each EC and MeJA treatment. The same volume of MeJA (392707, Sigma Aldrich, St. Louis, MO, USA) at concentrations of 0, 10, 20, or 40 µM was sprayed with a hand sprayer at 4 weeks after transplantation onto the canopy of A. rugosa twice per week thereafter.
Growth Measurements
Growth parameters of twenty plants were measured. At the end of the culture period, the leaves were detached from the plants. Stem length, leaf length, and leaf width were measured with a digital caliper (Digipa, Mitutoyo Co., Ltd., Tokyo, Japan). The third internode length from the apical meristem of each plant, petiole length, and the longest root length, were measured every week. Chlorophyll content of the plants after 4 weeks transplantation was measured by a portable chlorophyll meter (SPAD 502, Minolta Co., Ltd., Tokyo, Japan). Root volumes were measured using a mass cylinder filled with water. Fresh shoot and root weights were determined using a digital balance (ARG224 OHAUS, Sigma-Aldrich) after absorbing surface water by pressing between folds of lab wipes (Kimwipes S200, Japan Paper Crecia Co., Ltd., Tokyo, Japan). Shoots and roots were dried in an oven (HB-502M, Hanback Sci, Suwon, Korea) at 67°C for 7 days, and then weighed. All measurements were taken four weeks after transplantation. The tilianin and acacetin contents were measured in three plants subjected to EC treatment or MeJA treatment using high-performance liquid chromatography (HPLC).
Extraction and Analysis of Tilianin and Acacetin by HPLC
Shoot samples frozen in liquid nitrogen were first ground to a fine powder using a mortar and pestle. 0.1 g of powder sample was mixed with 1.5 mL of 80% methanol ethanol for 1 h at 60°C. The sample was centrifuged, and the supernatant filtered through a 0.45-µm polypropylene syringe filter (Acrodisc Syringe Filters, Pall, Port Washington, NY, USA) and subjected to HPLC analysis. Analysis was performed using a C-18 column (250 × 4.6 mm, 5 µm; RStech, Daejon, Korea) at 30°C. The mobile phase was a gradient mixture of acetonitrile, methanol, and 0.2% acetic acid. The flow rate was maintained at 1.0 mL·min-1, the injection volume was 20 µL, and the detection wavelength was 280 nm. Concentrations of tilianin and acacetin in the samples were calculated using a standard curve. Leaves and stems were analyzed for both concentrations in unit mg and contents in biomass (per plant). All mean values of tilianin and acacetin were obtained from three plant replicates.
Statistical Analysis
Each experiment was conducted twice with completely randomized designs. All data were subjected to analysis of variance followed by Tukey’s multiple range test at p ≤ 0.05 (SPSS version 24 software, SPSS Inc., Chicago, IL, USA).
Results and Discussion
Effect of EC on Plant Growth
A. rugosa plants were grown in a hydroponic culture system with different applied ECs. The EC of the nutrient solution affected leaf, stem, petiole, and root growth (Fig. 1). All growth parameters varied with the EC of the nutrient solution. Leaf length and width (Fig. 1A), internode length (Fig. 1B), petiole length (Fig. 1B), and stem length (Fig. 1C) were increasingly greater in nutrient solutions with increasing ECs up to 2.0 dS·m-1, but were lower at 4.0 dS·m-1. The same pattern was observed for shoot fresh and dry weight (Fig. 1E). The maximum values of the value of leaf length and leaf width were not match with Fig. 1A, stem length, internode length, and petiole length were 7.49, 6.18, 42.62, 8.89, and 3.95 cm, respectively, at an EC of 2.0 dS·m-1. Shoot fresh weight and dry weight were 32.85 and 5.17 g, respectively, at 2.0 dS·m-1. By contrast, root length and volume (Fig. 1D) increased with increasing EC, as did root fresh (Fig. 1F) and dry weights (Fig. 1F). Root length (30.0 cm) and root volume (12.0 mL) were the greatest when the plants were cultivated in nutrient solution with an EC of 4.0 dS·m-1. The maximum values for root fresh and dry weights at 4.0 dS·m-1 were 11.48 and 1.13 g, respectively.
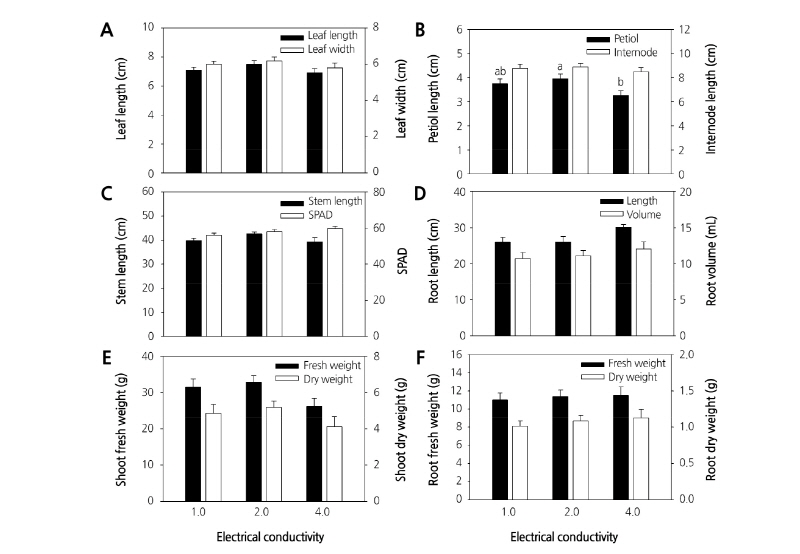
Fig. 1. Bar graph showing the effects of EC in nutrient solution on Agastache rugosa plant parts. Leaf length and leaf width (A), petiole length, internode length (B), stem length and SPAD (C), root length and volume (D), shoot fresh weight and dry weight (E), root fresh weight and dry weight (F). The plants were grown in a hydroponic culture system with nutrient solution at ECs of 1.0, 2.0, and 4.0 dS·m-1 in Otsuka House 1 and illuminated with fluorescent lamps for 4 weeks after transplantation. The data represent the means and the vertical bars indicate standard errors (n=20). Data points with different letters for the same measurement are significantly different according to Tukey’s multiple range test at p ≤ 0.05.
When nutrients in the nutrient solution are present at higher than optimal concentrations, the nutrients in solution increase salinity, raising EC. Salinity is a major abiotic stress. Negative effects of high salt concentration include delays in plant growth and development, inhibition of enzymatic activities, and reduction of photosynthetic rate (Lee et al., 2013). Salinity is the presence of electrolytic mineral solutes at concentrations that harm plants. Salinity can severely limit plant growth and production (Oliveira et al., 2013). High solute concentrations inhibit plant growth in two ways. First, they reduce osmotic potential in the rhizosphere as solute accumulation (Degl’Innocenti et al., 2009), reduce water uptake and turgor pressure (Munns, 2008; Zhang et al., 2006). Second, nutritional imbalances can develop (Zhu, 2007) as the concentration of nutrient elements and their ratios limit uptake and translocation of other elements, thus influencing plant growth (Tabatabaei, 2006).
In this study, leaf length and width, stem length, internode length, and petiole length first increased to maximum values at an EC of 2.0 dS·m-1, but were lower when the EC exceeded 2.0 dS·m-1. The optimal growth of these parts occurred at an EC of 2.0 dS·m-1. The shoot is the part of the plant most sensitive to increased solute concentrations (Karimi et al., 2009). Growth inhibition when EC is higher than 2.0 dS·m-1 is related to changes in the osmotic potential because of increased ion concentrations around the roots. The reduced plant growth observed for A. rugosa at a high EC may have resulted from reduced cell expansion, because of diminished water uptake and turgor pressure. The reduced growth at elevated EC may also have resulted from internal injury, due to accumulation of solutes in the leaves to levels exceeding the ability of cells to compartmentalize these solutes in the vacuole. Moreover, over-accumulation of solutes such as Na+ and Cl– cause deficiencies in other important nutrients such as Ca2+ and K+, which adversely affects plant growth (Marschner, 1995). High solute concentrations often induce hormonal signals originating from the roots, mainly in the form of abscisic acid, which causes stomatal closure (Gill and Tuteja, 2010).
It has been reported that EC in single-truss tomato plants increased from 1.8–2.0 dS·m-1 to approximately 8.1–9.2 dS·m-1 and chlorophyll concentration increased in leaves by 14% (Romero-Aranda et al., 2001). The total chlorophyll content decreased in nutrient solution with high EC (Dewir et al., 2005). In this study, chlorophyll content peaked at an EC of 4.0 dS·m-1. A high EC of 2.2 dS·m-1 significantly decreased the photosynthetic capacity of potato plantlets (Piao et al., 2004). Carbon assimilation occurs via photosynthesis and is necessary for plant growth and yield. Photosynthetic capacity can be affected by stomatal factors, non-stomatal factors, or both (Saibo et al., 2009). High soil salinity causes stomatal closure and a reduction in a photosynthesis (Saleem et al., 2011; Yan et al., 2012). The optimum EC of a balanced nutrient solution is approximately 1.5 dS·m-1 (Dewir et al., 2005). Some plants are more susceptible to EC and each plant species has a specific EC threshold, beyond which yield is decreased. Electrolytes influence electro-physiological parameters involved in controlling cell expansion and coordinating the growth underlying tropisms and mutations (Spalding and Cosgrove, 1993). However, optimum EC levels vary by plant species, season, growth stage, and water quality.
Effect of EC on Tilianin and Acacetin Contents
At 4 weeks after transplanting, A. rugosa plants grown in the hydroponic culture system with nutrient solutions of different EC were analyzed for secondary metabolite content. The total contents of both tilianin and acacetin varied with ECs. The variation pattern in tilianin and acacetin with ECs of the nutrient solution was similar. The quantity of tilianin and total tilianin was calculated per gram of dry weight and per plant based on the dry weight. The amount of tilianin per gram dry weight of A. rugosa increased with increasing EC. Tilianin content per gram dry weight was 1.35 mg·g-1 DW at 1.0 dS·m-1 and reached a maximum of 2.10 mg·g-1 DW at 4.0 dS·m-1 (Fig. 2A). Acacetin per gram dry weight was 19.08 mg·g-1 DW at 1.0 dS·m-1 and reached a maximum of 21.84 mg·g-1 DW at 4.0 dS·m-1 with an intermediate content of 17.14 mg·g-1 DW at 2.0 dS·m-1. Thus, acacetin content first decreased with increasing EC and then increased (Fig. 2A). Total tilianin and acacetin content per plant based on the dry weight of plants grown hydroponically in nutrient solutions with different ECs varied. The tilianin concentration was 6.52 g/plant at an of 1.0 dS·m-1, 9.67 g/plant at 2.0 dS·m-1, and 8.65 g/plant at 4.0 dS·m-1. Initially, tilianin concentration increased, and then remained constant. By contrast, the total acacetin continually increased from 41.17 g/plant at 1.0 dS·m-1 to a maximum value of 50.00 g/plant at 4.0 dS·m-1 with an intermediate value of 44.27 g/plant at 2.0 dS·m-1 (Fig. 2B).
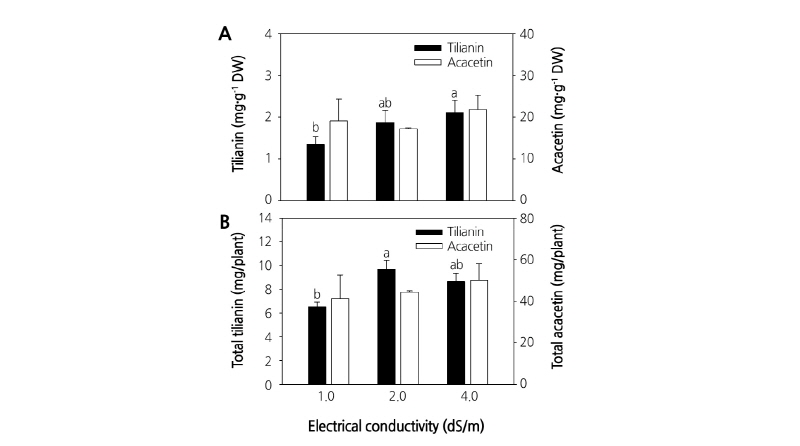
Fig. 2. Tilianin and total acacetin contents varied with EC. A: Secondary metabolite (mg) per dry weight (g). B: Metabolite (mg) per plant. Agastache rugosa was grown under hydroponic culture system at 1.0, 2.0, and 4.0 dS·m-1 electrical conductivities in Otsuka House 1 Nutrient Solution and illuminated with fluorescent lamps for 4 weeks after transplantation. The data represent the means and the vertical bars indicate standard errors (n=3). Data points with different letters for the same measurement are significantly different according to Tukey’s multiple range test at p ≤ 0.05.
Comparison of the secondary metabolites revealed that the amount of total acacetin accumulated in A. rugosa was always more than that of tilianin. Tilianin in A. rugosa grown in nutrient solution of EC 2.0 and 4.0 dS·m-1 was 39% and 56% higher than that of plants grown at EC 1.0 dS·m-1, respectively. However, acacetin in A. rugosa grown at EC 2.0 decreased by 10% but increased by 15% at EC 4.0 dS·m-1 compared with that of plants grown at EC 1.0 dS·m-1 for 4 weeks. Compared with EC 1.0 dS·m-1, accumulation of secondary metabolites in the whole plants, based on dry weight, increased by 48 and 36% for tilianin and 8 and 21% for acacetin at EC 2.0 and 4.0 dS·m-1, respectively. These results indicate that nutrient solution with a higher EC may induce accumulation of tilianin and acacetin in hydroponically grown A. rugosa, without radical growth reductions.
Numerous environmental factors such as CO2 concentration, temperature, humidity, light, water supply, and mineral salts, influence plant growth and directly impact biochemical pathways, which affect secondary metabolite production (Ramakrishna and Ravishankar, 2011). Excess Na+ and Cl– in the soil solution creates osmotic stress and causes plants to accumulate osmotically active compounds to lower osmotic potential (Ahmad and Sharma, 2008). Excessive salt concentrations trigger plants to control ion partitioning between different sink tissues (Tyerman et al., 1997). Salt-driven osmotic stress induces oxidative stress caused by reactive oxygen species. Plants have developed a complex antioxidant defense system containing small molecules such as ascorbate, reduced glutathione, flavonoids, carotenoids, tocopherol, and some antioxidant enzymes such as catalase, peroxidase, ascorbate peroxidase, and superoxide dismutase (Azevedo-Neto et al., 2006). Some studies have examined the enhancement of secondary metabolite production with increasing EC of the nutrient medium in hydroponic culture. The high EC of 4.5 dS·m-1, compared to the standard low EC of 2.3 dS·m-1, enhanced the lycopene concentration in tomato when grown hydroponically (Wu et al., 2004). Lycopene synthesis was promoted by osmotic and/or salt stress caused by the high EC (Wu and Kubota, 2008).
In addition, plants divert the synthesis of carbohydrates to produce secondary metabolites under saline conditions. It has been shown that moderate salinity induces the normal saline tolerance pathway to increase total phenolic compounds contents (Salem et al., 2014). Increasing the salinity to 10 g·L-1 NaCl increased the flavonoid contents of safflower (Gengmao et al., 2015).
Combined Effect of MeJA and EC on Phenylpropanoid Levels
A. rugosa seedlings were grown under different EC of nutrient solution with foliar application of MeJA at different concentrations at regular intervals, twice in the last week before harvest. Tilianin and acacetin accumulation in plants were analyzed at 4 weeks after transplantation into the hydroponics culture system. Both tilianin and acacetin contents were influenced positively by both increasing concentrations of MeJA and increasing EC of the nutrient solution (Fig. 3). In nutrient solution at each EC, increasing the MeJA concentration to 20 µM increased tilianin levels to a maximum value. The levels of tilianin were 1.61, 2.49, and 2.75 mg·g-1 DW at 1.0, 2.0, and 4.0 dS·m-1, respectively. Accumulation of tilianin was 1.5- and 1.7-fold higher at 2.0 and 4.0 dS·m-1, respectively, compared to that at 1.0 dS·m-1.
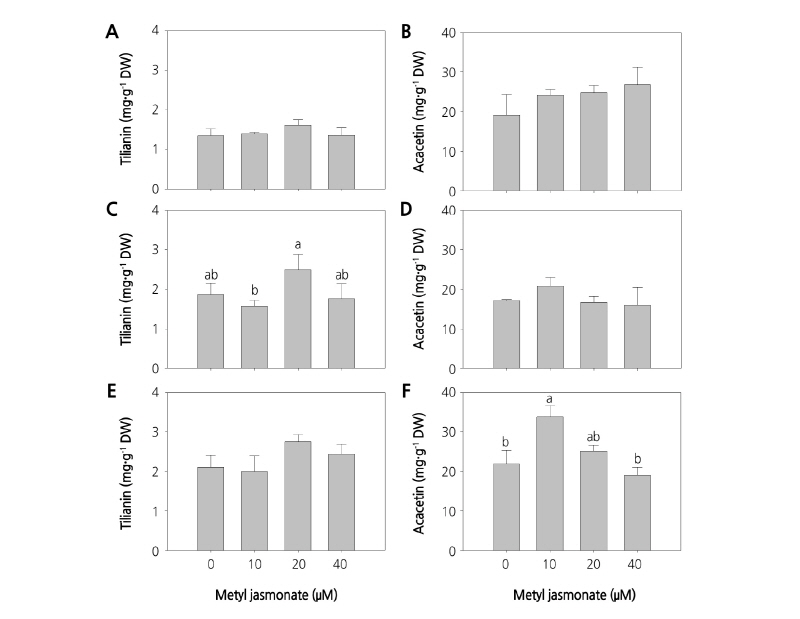
Fig. 3. Tilianin and total acacetin contents varied with MeJA application in Agastache rugosa. The plants were sprayed twice with 0, 10, 20, and 40 µM methyl jasmonate from the 4th week after transplantation and grown in a hydroponic culture system with 1.0 dS·m-1 (A, B), 2.0 dS·m-1 (C, D), and 4.0 dS·m-1 (E, F) conductance, and illuminated with fluorescent lamps, for 4 weeks. The data represent the means and the vertical bars indicate standard errors (n=3). Data points with different letters for the same measurement are significantly different according to Tukey’s multiple range test at p ≤ 0.05.
Variation of acacetin in the extract sample differed from that of tilianin. At the lowest EC of 1.0 dS·m-1, total acacetin levels gradually increased with increasing concentrations of MeJA and peaked at 26.83 mg·g-1 DW at 40 µM of MeJA. This trend was not observed at 2.0 and 4.0 dS·m-1. At both conductivities of 2.0 and 4.0 dS·m-1, total acacetin first increased to a maximum when 10 µM MeJA was used, and then decreased when 40 µM was used. The maximum values of total acacetin were 20.86 and 33.81 mg·g-1 DW at 2.0 and 4.0 dS·m-1 following spraying with 10 µM MeJA. The highest value of tilianin and was 2.75 mg·g-1 DW at 20 µM of MeJA and 4.0 dS·m-1, whereas the highest value of total acacetin was 33.81 mg·g-1 DW at 10 µM of MeJA and 4.0 dS·m-1 EC of nutrient solution.
Plants synthesize several classes of phenolic compounds via the shikimate pathway. A. rugosa produces several types of essential oils, sesquiterpenes, diterpenes, triterpenes, flavonoids, and carotenoids. Tilianin and acacetin are flavonoids. Tilianin is found in various plants as a glucose-glycoside compound of acacetin. Tilianin shows anti-inflammatory (Nam et al., 2005), antiatherogenic (Hong et al., 2001), antihypertensive, and vaso-relaxant effects (Hernandez-Abreu et al., 2009).
JA and its ester MeJA are signaling molecules that play key roles in plant growth, development, and response to biotic and abiotic stresses. Plants exposed to stress produce MeJA, which induces the expression of defensive genes and activates the defense response in plants. Phenolic acids and their derivatives play key roles in this defense response process (Creelman and Mullet, 1997). Elicitors interact with plant membrane receptors and generate signaling compounds, which subsequently activate specific genes of highly inducible enzymatic systems involved in secondary metabolite biosynthesis. Generally, MeJA is an elicitor that enhances the biosynthesis of flavonoids and phenolic acids. Application of 50 µM MeJA to A. rugosa increased phenolic acid production and phenylpropanoid biosynthetic gene expression to a greater extent than did any other concentration of MeJA tested (Kim et al., 2013). MeJA was used to induce the abundant production of flavonoids in plant cell cultures (Fang et al., 1999). MeJA treatments induced phenylalanine ammonia-lyase activity, which led to greater accumulation of total phenolic compounds in romaine lettuce (Lactuca sativa L.) and sweet basil (Ocimum basilicum L.) (Kim et al., 2006; Kim et al., 2007). Furthermore, MeJA treatment dramatically increased the levels of some secondary metabolites in Nicotiana attenuata leaves (Keinänen et al., 2001); increasing the relative abundances of methyl benzoate and methyl salicylate in tuberose (Kanani and Nazarideljou, 2017); enhancement certain bioactive compounds, such as ascorbic acid (AsA) and superoxide dismutase (SOD) in ‘queen’ pineapple PAC tissues (Boonyaritthongchai and Supapvanich, 2017); stimulated the production of phenolic acids, particularly ferulic, chlorogenic, and syringic acids, in Ginkgo biloba (Szewczyk, 2008); induced the biosynthesis of phenolic acids and flavonoids and ginsenoside increasing in Panax ginseng (Ali et al., 2007; Um et al., 2017); and upregulated sorgoleone biosynthesis in sorghum (Uddin et al., 2013) and the production of rosmarinic acid in A. rugosa (Kim et al., 2013).
In conclusion, the growth of A. rugosa was lower when cultured in nutrient solution with elevated ECs, possibly because of reduced cell expansion, which is often the result of diminished water uptake and turgor pressure. High EC and spraying with MeJA are individually important factors affecting secondary metabolite accumulation. In addition, the effect of MeJA was more pronounced when plants were grown in high EC nutrient solution. Our findings suggest that high EC nutrient solution and spraying with low concentrations of MeJA together strongly activate phenylpropanoid biosynthesis genes.
