

서 언
감귤은 국내외 과수 산업에서 가장 중요한 작물의 하나이다(FAO, 2012; MFAFF, 2011). 최근의 기후 변화에 따른 기상 이변, 돌발 병해충의 발생, 노동력 부족, 다양한 과실의 공급 증가 등은 세계 감귤 산업의 지속 발전에 다양한 위협 요인이 되고 있다. 특히 고품질, 친환경, 기능성 및 다양성에 대한 소비자들의 관심이 급격히 높아지고 있다. 이들 다양한 위협 요인과 소비자 요구에 신속히 대응하기 위해서는 다양한 변이의 창성과 우수 품종의 선발이 요구되고 있다. 이에 따라 다양한 종내 및 종간 교배가 이루어지고 있다. 그런데 우수한 과실 형질을 나타내는 교배 실생 개체라 하더라도 과실의 종자가 많다면 신품종으로서의 가치는 잃게 될 것이다(Recupero et al., 2005). 그러므로 최근 고품질 무핵 품종의 육성을 위한 삼배체 육종이 국내외적으로 가장 중요한 육종 목표의 하나가 되고 있다(Zhu et al., 2009).
무핵 삼배체의 육성에는 이배체와 사배체 간의 정역 교배가 가장 보편적으로 이용된다(Ferrante et al., 2009; Recupero et al., 2005). 감귤 및 근연종의 대부분은 이배체(2n = 18)이다. 삼배체 육종을 위해서는 유전체가 배가된 사배체의 양성이 필수적이다. 사배체의 양성은 콜히친 처리에 의한 인위 유도(Dutt et al., 2010; Wakana et al., 2005), 체세포 융합에 의한 인위 유도(Grosser et al., 1994, 2000; Medina-Urrutia et al., 2004) 및 주심배 자연 발생 사배체의 선발(Ferrante et al., 2009; Kaneyoshi et al., 2008; Oiyama and Kobayashi, 1991; Song et al., 2011; Usman et al., 2006)로 구분할 수 있다. 이들 중에서 주심배 배수체의 자연 발생은 운향과의 감귤아과에 속하는 감귤속 식물은 물론 근연속인 탱자속 및 금감속에서도 보고되어 왔다(Cameron and Frost, 1968). 이들 사배체의 발생 빈도는 2-5% 내외로 알려지고 있다. 일반적으로 사배체 감귤 및 탱자는 잎이 두껍고 크며, 줄기의 생장은 느리고 왜화성이며 직립성은 떨어진다. 과실의 수량은 낮고, 과실은 다소 크나, 과피가 두껍고 양낭의 수가 적은 편이다. 그러나 일부 사배체에서는 이와 반대의 경향을 나타내기도 한다(Cameron and Frost, 1968). 일부 감귤 동질 사배체에서의 형태적 변이에 대해 보고된 바 있으나(Romero-Aranda et al., 1997; Wakana et al., 2005), 아직까지도 변이의 정도와 그 기작에 대한 연구는 거의 알려진 바가 없다.
한편 일반 식물에서도 다양한 동질 배수체 및 이질 배수체가 보고된 바 있고, 일부 배수체들은 기존 이배체와 다른 표현형을 나타내기도 하고(Gunasekare and Ranatunga, 2003; Osborn et al., 2003), 변이를 보이기도 한다(Ranatunga and Gunasekare, 2003; Ye et al., 2010). 아직까지도 이들 기작에 대해서는 잘 알려져 있지 않으나 염색체 수의 증가와 더불어 중복된 대립 유전자 및 전사 조절 인자들의 발현 증대, 유전자 재배열, 후성 유전에 의한 유전자 발현 변화에 기인하는 것으로 보고되고 있다(Osborn et al., 2003; Sémon and Wolfe, 2007). 후성 유전은 유전체 염기 서열의 변화가 아닌 유전체의 염기 서열의 메틸화, 히스톤의 변형, 염색질의 구조 변화 등에 기인한다. 일반 식물에서도 이질 배수체와는 달리 동질 배수체의 후성 유전의 영향에 대한 연구 보고는 거의 없는 실정이다(Osborn et al., 2003; Sémon and Wolfe, 2007). 따라서 본 연구에서는 자연적으로 발생한 탱자 동질 사배체들로부터 몇 가지 수체 특성 표현형의 변이 정도와 유전체 메틸화의 변이 정도를 분석하여 후성 유전의 하나인 유전체 메틸화가 동질 사배체의 표현형 변이와 관계되는 하나의 요인으로 작용할 수 있음을 구명하고자 하였다.
재료 및 방법
식물 재료
제주특별자치도 서귀포시에 위치한 국립원예특작과학원 감귤시험장에 재식된 이배체 탱자 주심배 유래의 3년생 사배체 탱자(Poncirus trifoliata (L.) Raf.) 14주와 대조군인 이배체 탱자 3주를 시험수로 이용하였다.
염색체 및 동질 사배체 분석
배수성 확인을 위해 Dutt et al.(2010)의 방법에 따라 염색체 분석을 수행하였다. 탱자 사배체 식물로부터 유엽을 채취하여 2mM 8-hydroxyquinoline 용액에서 전처리 한 후 고정액(메틸알코올:초산 = 3:1)으로 처리하였다. 고정된 시료는 세포 분해 효소액(2% cellulase Onezuka RS, 1% macerozyme 및 0.3% pectolyase Y-23)으로 37°C에서 1시간 처리한 후 슬라이드글래스 상에 세포들을 단리시켰다. 슬라이드 시료는 2% Giemsa 용액을 이용으로 염색하였으며, 현미경 하에서 염색체 관찰을 통해 탱자의 배수성과 이수성 여부를 확인하였다.
동질 사배체 여부를 확인하기 위하여 Chromomycin A3 (CMA) 염색에 의한 핵형 분석을 실시하였다. Befu et al. (2000)의 방법에 따라 McIlvaine’s buffer와 5.0mM MgCl2 완충용액을 전처리한 후 0.12mg・mL-1 CMA에 30분간 처리한 다음 4, 6-diamidino-2-phenyindole를 20분간 처리한 후 형광 현미경(LEICA DMRBE, Germany)으로 핵형 분석을 통해 동질 사배체 여부를 확인하였다.
생장 특성 분석
포장에 재식된 사배체 탱자로부터 수고, 수폭, 원가지 수, 원가지 길이, 분지 각도, 마디 길이 등을 조사하였다. 그리고 생장이 정지된 신초 상부의 잎에서 엽장, 엽폭, 잎의 두께, 엽병 길이를 조사하였다. 잎의 광합성 및 엽록소 지수는 사배체 탱자 상부의 가까운 잎에서 맑은 날 오전 10시경에 측정하였다. 광합성 측정은 LI-6400 portable photosynthesis system(LI-COR, Ltd., Lincoln, NE, USA)을, 엽록소 지수는 SPAD-502(Minolta, Tokyo, Japan)를 이용하여 측정하였다. 또한 사배체 탱자의 잎 이면에 일반 매니큐어를 바른 후 건조시킨 다음, 이를 핀셋으로 분리하고 슬라이드글래스 위에 거치하여 현미경 하에서 기공 특성을 관찰하였다.
Global Cytosine DNA 메틸화 분석
Global cytosine DNA 메틸화 분석은 Demeulemeester et al.(1999)과 Jia et al.(2011)의 방법을 일부 변형하여 수행하였다. 사배체 탱자의 잎으로부터 DNA를 CTAB법으로 추출하고, HClO4를 처리하여 DNA를 분해한 후 gradient HPLC (LC-20A HPLC series, Shimadzu, Kyoto, Japan)에 의한 nucleoside chromogram을 분석하였다. 분석 컬럼은 ODS C18(4.6 × 250mm, 5µm, Shimadzu)을 사용하였고, 이동상은 7% 아세트산 + 3% 아세토니트릴 + 증류수의 A용매와 증류수로 이루어진 B용매로 사용하였다. 유속은 분당 1mL, 온도는 40°C, 최대 온도는 85°C로 설정하였다. Global cytosine DNA 메틸화 정도(%)는 {[methylated DNA cytosine/(non- methylated + methylated)] × 100}으로 계산하였다.
통계 검정
통계 처리는 Sigmastat 3.5(SYSTAT software Inc., USA)를 이용하여 수행하였다.
결과 및 고찰
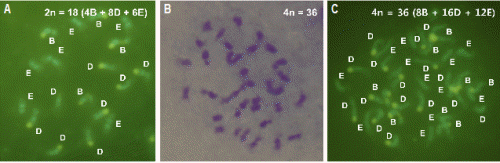
Flow cytometry 분석에 의해 이배체 탱자의 주심배 실생으로부터 선발한 2년생 사배체 탱자 14주의 유엽으로부터 염색체 및 핵형 분석을 수행하였다(Fig. 1). Flow cytometry 분석과 Giemsa 염색에 의한 염색체 수의 분석 결과, 사배체 식물체 14주는 모두 이수성이 없는 2n = 4X = 36 식물체로 확인되었다. CMA 염색에 의해 핵형을 분석한 결과, 이배체 탱자는 4B + 8D + 6E인 반면, 사배체 탱자는 모두 8B + 16D + 12E로 나타났다.
이배체 탱자로부터 사배체 식물체의 발생은 수정된 이배체 배의 초기 발육과정 동안 체세포의 염색체 배가 또는 주심조직의 염색체 배가된 세포 기원의 무성생식 주심배 발달로 가능할 수 있다. 일반적으로 만다린, 오렌지, 자몽, 라임, 탱자 등의 감귤류에서 사배체 식물의 발생은 체세포 유래의 주심조직에서 무수정 생식에 의해 일어나는 것으로 알려져 있다(Usman et al., 2006). Aleza et al.(2011)은 22개의 감귤, 탱자, 또는 이들 잡종에서 얻어진 580개 사배체 식물의 SSR 분석을 통해 이들 사배체의 기원은 자식 또는 타식에 의한 수정 접합체의 염색체 배가가 아닌 주심조직 세포의 염색체 배가에 의해 얻어지는 것을 제시한 바 있다. 감귤 CMA 염색은 감귤의 핵형을 구분하여 양친형의 동정 및 잡종성을 확인하는데 유용하게 이용되어 왔다(Barros e Silva et al., 2010; Yamamoto and Tominaga, 2004; Yamamoto et al., 2007). 본 연구에서 수행된 이배체 탱자의 CMA 핵형 분석 결과는 이전에 보고(Miranda et al., 1997)와 일치하는 4B + 8D + 6E였으며, 사배체 탱자의 CMA 핵형은 8B + 16D + 12E 로 이배체가 배가된 형태를 보여주었다. 그러므로 이들 사배체 탱자는 이전에 보고되어 온 바(Aleza et al., 2011; Usman et al., 2006)와 같이 수정배의 염색체 배가가 아닌 무성생식 주심배 유래의 동질 사배체인 것으로 확인할 수 있었다.
동질 사배체 탱자 14주의 생장 특성을 분석하기 위해 수고, 수폭, 원가지 수, 원가지 길이, 분지 각도, 마디 길이, 그리고 잎의 특성을 조사하였다(Tables 1 and 2). 동질 사배체 탱자 14주의 수고는 평균 94.7cm로, 수폭은 평균 55.5cm로 이배체보다 다소 작은 편이었다. 동질 사배체 탱자의 원가지 수는 1-8개로 다양하게 나타났는데, 대부분이 5개 이상의 원가지를 가지고 있었으나, 3개 이하의 극히 적은 분지 수를 나타내는 개체의 비율이 35.7%로 관찰되었다. 원가지 길이, 분지 각도 및 마디 길이에 있어서도 이배체 탱자와 사배체 탱자 간의 뚜렷한 차이는 나타나지 않았다. 그러나 분지 각도에 있어서는 매우 좁게 나타나는 것과 매우 넓게 나타나는 일부 개체가 관찰되었다. 동질 사배체 탱자 14주의 수형을 보면 직립형은 6주, 반직립형은 6주, 개장형 2주로 조사되었다. 이에 반해 대조군인 이배체 탱자는 모두 직립형로 관찰되어 이배체 탱자의 특성상 수형은 직립형으로 자라는 특징이 있는 것으로 생각된다. 그러나 이배체 탱자의 조사 집단이 작았기 때문에 이배체 탱자에서의 직립형 특성이 사배체와 구분할 수 있는 표현형인지는 분명하지 않았다.
잎의 길이는 다양하게 나타났으며, 사배체 탱자 10주가 이배체 탱자보다 길게 나타났다. 엽폭에 있어서는 사배체 탱자 내 변이가 다양하게 나타났음에도 불구하고 모든 개체에서 이배체의 엽폭보다 크게 나타났다. Usman et al.(2006)와 Wakana et al.(2005)에 의하면 이배체의 경우 사배체보다 엽폭이 1.13-1.5배가 좁다고 하는데, 본 연구 결과에서도 대조군인 이배체 탱자보다 사배체 탱자 14주 모두 엽폭은 넓은 경향을 보였으며, 그 비율은 1.04-1.51% 범위에 있었다. 잎의 두께는 사배체 탱자 14주 중 50%가 이배체보다 작거나 같았고, 50% 이상은 두꺼운 것으로 나타났으며, 엽병 길이는 사배체 탱자 1개체(T5)에서의 15.1cm를 제외하고는 이배체 탱자보다 짧은 것으로 조사되었다. 전체적으로 볼 때, 나무 크기, 엽폭, 엽병 길이 등은 보편적인 사배체 식물의 특징을 보이는 편이었으나, 수형과 잎의 길이, 엽의 두께에 있어서는 전형적인 특징을 보이지 않은 편이었으며, 보편적인 특성에 있어서도 상당한 표현형의 변이 폭을 나타내는 것을 보여 주고 있어 동질 사배체 탱자에서 수체 형질마다 표현이 다르게 나타나는 것을 알 수 있었다.
사배체 탱자 상부의 가장 가까운 잎에서 측정한 광합성률과 엽록소 지수를 Tabal 3에 나타내었다. 사배체 탱자의 광합성률은 17-21μmole CO2・m-2・s-1 범위에서 나타났고, 사배체 탱자 간에도 광합성률에 차이가 있었다. 이배체 탱자의 광합성률과 비교할 때 일부 사배체 탱자(T4, T12 및 T13)를 제외하고는 높은 수준이었다. SPAD 값에 의한 엽록소 지수에 있어서 사배체 식물에서는 전체적으로 48-75의 범위로 나타났는데, 이는 이배체 식물체의 엽록소 지수인 62와 비교할 때 분리되는 경향처럼 보였으나, 분명하지는 않았다. 감귤에서는 일반적으로 사배체의 잎은 이배체의 잎보다 진하여 SPAD 값이 높으나 광합성율은 낮은 경향을 보이는 것으로 알려져 있는데(Allario et al., 2011; Romero-Aranda et al., 1997), 본 연구결과와는 차이가 있는 것으로 환경과의 상호작용에 따른 변이 등은 상세한 연구가 필요하다고 생각된다.
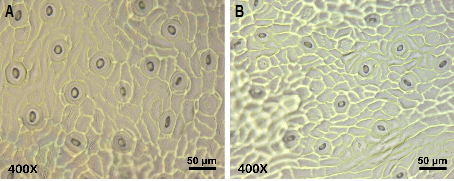
사배체 탱자 간의 기공의 크기와 밀도를 비교한 결과를 Table 4와 Fig. 2에 나타내었다. 사배체 탱자의 기공 밀도는 97-166mm2의 범위로, 공변세포 길이는 28-50μm의 범위에서 나타났다. 기공 밀도는 사배체 탱자에서 모두 이배체 탱자에서보다 낮았으며, 기공 크기도 1개체(T4)만 제외하고는 모두 크게 나타났다. 그러나 개체에 따라서는 T6의 경우에서와 같이 대조군 이배체 탱자의 기공 밀도에 비해 46% 수준으로 매우 낮은 것도 조사되었다. 이는 감귤의 경우 이배체와 비교하여 사배체에서 기공의 크기는 커지고 분포 밀도는 낮아진다는 보고(Song et al., 2011; Usman et al., 2006)와는 차이가 있으나, 차나무의 경우 삼배체에서 오히려 이배체보다 기공의 크기도 작고 분포 밀도가 증가하는 보고(Gunasekare and Ranatunga, 2003)와는 일치하는 경향으로 나타났다.
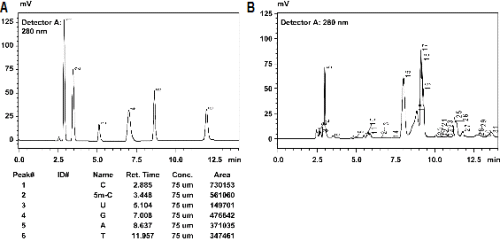
Gradient HPLC 분석에 의한 동질 사배체 탱자의 global cytosine DNA 메틸화 비율을 분석하였다(Table 5 and Fig. 3). 동질 사배체 탱자의 global cytosine DNA 메틸화를 분석한 결과 6.4-17.6% 범위로 나타났으며, 개체 간 메틸화 정도에 변이가 존재함을 확인할 수 있었다. 또한 이배체 탱자의 메틸화 수준과 비교하였을 때, 14주 사배체 탱자의 절반이 2배 이상 높게 나타났고, 거의 대부분이 이배체 탱자보다 높은 경향을 나타내며, 전체적으로 볼 때 유의하게 높게 나타나는 것을 알 수 있었다. 이것은 염색체 배가에 의해 발생되는 gene redundency를 줄이기 위한 것으로 생각된다.
동질 배수체에서도 이질 배수체에서 나타나는 유전체 재배열과 메틸화에 의한 유전자 발현의 조절이 이루어지고, 이들 유전체의 구조적 변화와 메틸화 정도는 종과 배수화 정도에 따라 다르게 나타나는 것으로 알려지고 있다(Aversano et al., 2013; Wang et al., 2009). 그러나 야생 감자(Aversano et al., 2013), 수박(Wang et al., 2009), 감귤(Aleza et al., 2011), 향모속 식물(Lavania et al., 2012), 애기장대(Yu et al., 2010) 등 대부분의 동질 배수체 연구에서 유전자 배열의 구조적 변화는 매우 미미하여 배수체에서 나타나는 표현형의 변이는 메틸화와 관계된다고 하였다. 특히 향모속 6개 종의 동질 사배체에서 나타나는 표현형의 변이가 사배체에서 증가된 메틸화와 관계되며(Lavinia et al., 2012), 야생 감자에서 사배체들간 나타나는 메틸화 정도의 상당한 변이가 표현형의 변이와 관계된다고 하였는 바, 본 연구결과와 매우 유사하였다. 그러나 Wang et al.(2009)은 이배체, 삼배체 및 사배체의 수박에서 메틸화의 정도는 유사하나 분포는 다르게 나타난다고 하였는 바, 사배체 탱자에서 나타나는 표현형의 변이가 메틸화 분포와 관계될 수도 있다고 생각되며, 이에 대해서는 추가적인 연구가 필요한 것으로 보아진다.
|
Fig. 3. Chromatograph of global cytosine DNA metylation from standard solution (A) and an autotetraploidy trifoliate orange (B). |
본 연구 결과 탱자에서 같은 동질 사배체일지라도 수고, 수폭, 원가지 수, 원가지 길이, 분지 각도, 마디 길이, 잎의 특성, 광합성률, 엽록소 지수, 기공 밀도 등이 개체마다 표현형 변이가 나타나는 것을 알 수 있었다. 이배체 탱자와 사배체 탱자를 비교하였을 때는 표현형마다 비슷한 경향을 보이기도 하고, 다른 경향을 보이기도 하는 것을 알 수 있었다. 또한 동질 사배체의 global cytosine DNA 메틸화 정도는 이배체에 비해 증가하는 경향이었다. 그러나 식물체 생장 특성에 대한 표현형 변이와 global cytosine DNA 메틸화 간의 상관을 찾아내지는 못하였다. 동일한 이배체 탱자로부터 선발된 주심배 유래의 동질 사배체라 하더라도 global cytosine DNA 메틸화 수준에 따라서는 하나의 게놈에 수많은 에피게놈(epigenome)이 존재할 수 있으며, 이에 따라 각 개체는 유전자 발현 양상에 차이를 보이게 되며, 각기 다른 표현형을 나타내는 것으로 보인다. 그러므로 유전체 메틸화 수준이 앞에서 조사했던 동질 사배체 탱자의 생장 특성, 광합성률, 엽록체 지수, 기공 밀도 등 다양한 형질 표현형 변이를 유발하는 하나의 요인으로 작용하고 있는 것으로 생각된다. 그러나 식물체의 생장 특성의 발현에는 유전적인 요인이외에도 온도, 양분 공급 등 다양한 환경적 요인들이 작용할 수 있기 때문에 추후 더욱 상세한 연구가 필요하다고 판단된다.
