

Introduction
Materials and Methods
Propagation and Postharvest Treatments of Bulblets
Bulblet Planting and Bulb Harvesting
Data Collection and Statistical Analysis
Results
SER after Bulblet Planting
SED after Bulblet Planting
BFW at Harvest Time
BER at Harvest Time
Correlation between the Fresh Weights of Bulblets Planted and Bulbs Harvested
Discussion
Introduction
Lilies native to Korea are part of the original species of Asiatic hybrids. Owing to their unique ornamental value, they can be used as cut flowers or for differentiation and gardening (Kim et al., 2015; RDA, 2021). Earlier, native lilies were propagated by seed propagation and scaling. The bulb production efficiency of native lilies is low because bulbs are obtained by scaling bulblets derived from collected seeds. Therefore, the systematic production and distribution of high-quality bulbs is absent in Korea (Kim et al., 2015).
The general methods of the propagation of horticultural varieties of lily can be divided into seed propagation and vegetative propagation. As vegetative propagation methods, propagation by bulblets and bulbils, scaling, and tissue culturing have been widely studied (Kim and de Hertogh, 1997; Daneshvar Royandazagh, 2019). Among these methods, scaling and tissue culturing are effective for mass reproduction. Native lilies produce good seeds by natural pollination, and there is little variation between individuals, meaning that it is possible to cultivate bulbs by seed propagation. However, this method requires two to four years to cultivate flowering bulbs from seeds. The bulb production process of lilies begins with bulblets and, depending on the species and purpose, these small buds can be sold as flowering plants only after they have been enlarged for two to four years.
Studies of native or wild lilies have concentrated on cultivation methods, physiology, ecology (Eum et al., 2011; Jeong, 1998a, 1998b; Son and Yun, 1998; Salehi et al., 2019; Mazor et al., 2021), tissue culture technology, and propagation (Kim et al., 2002, 2003, 2011; Goo et al., 2004; Eum et al., 2006, 2008, 2010; Ko et al., 2006), among other aspects. However, these studies were carried out independently and were not closely related to each other. Therefore, it is urgent and critical to establish a sound bulb production system for the conservation and utilization of wild lilies.
To produce high-quality flowering bulbs efficiently, it is necessary to develop a method to increase the efficiency of bulb enlargement in greenhouses and open fields. According to previous studies, larger bulbs are produced with larger seed bulbs or bulblets. In three cultivars of Lilium Oriental hybrids, the values of the fresh weight and circumference of the bulbs at harvest were higher with larger seed bulbs (Kim et al., 2013). In Triteleia laxa, the fresh weight of the planted corms increased as the weight of the harvested corms increases (Han et al., 1991). In Tulipa gesneriana, the BFW at harvest was higher with larger planted bulbs (Yoo and Roh, 2014). Therefore, to improve bulb production efficiency, it is necessary to investigate the propagation method and planting time with strategies that significantly enhance the efficiency for each species. efficiency for each species.
This study examined an efficient bulb production method by comparing shoot emergence and enlargement patterns by planting bulblets obtained through tissue culture, scaling, and seed propagation processes in a field in autumn and spring for four native Korean lilies with high development value as ornamental plants.
Materials and Methods
Propagation and Postharvest Treatments of Bulblets
Four species of native Korean lilies were grown in a farm field at Yeungnam University. Lilium amabile Palibin, L. cernuum Komar., L. lancifolium Thunb. (syn. L. tigrinum Ker Gawler), and L. maximowiczii Regel. [syn. L. leichtlinii var. maximowiczii (Regel) Baker] were selected as the experimental materials. These species belong to the Sinomartagon section. Propagation materials from healthy plants were collected, and one-year-old bulblets were obtained through tissue culture, scaling, and seed propagation processes and were prepared as plant materials (Table 1). Tissue-culture-derived bulblets were obtained using the method described by Kim et al. (2011). Small bulbs, one year after scaling and sowing, were used as plant materials for scaling and seed propagation. All bulblets were stored in wet cold storage at 0°C until planting.
To obtain scaling-derived bulblets, the outermost scales of the bulbs were removed and clean scales were separated from the bulbs and cured at room temperature. Then, the scales were immersed in benlate solution with a concentration of 2 g·L-1 at room temperature for 1 h and thoroughly washed with tap water. The scales were inserted to a depth of 2 cm into flower boxes filled with potting media (Sunshine® Mix #5, Sun Gro Horticulture, MA, USA) and moved to a glass greenhouse maintained at 20 ± 3°C. After five months of scaling, bulblets were harvested and stored in wet cold storage at 0°C until planting.
To obtain bulblets derived from seedlings through seed propagation, pods that had turned 80–90% brown and had not yet burst were collected in early autumn, and the seeds were separated from the pods and stored at 4°C. After three months, seedlings that germinated from seeds sown in Petri dishes were transferred to plug trays and grown for eight weeks in a greenhouse.
After planting these seedlings in containers for transplants, the bulblets obtained by cultivation for five months were stored at a low temperature. Three months later, they were planted again, grown in the field, and harvested in the fall. Bulblets for fall planting were planted immediately, and those for spring planting were stored wet at 0°C until planting.
Bulblet Planting and Bulb Harvesting
On October 30 (autumn planting) and February 23 of the following year (day of year, DOY 54; spring planting after low-temperature storage), 200 uniform bulblets produced through the tissue culture, scale propagation, and seed propagation steps were selected, divided into four groups (replicates), and planted in a completely randomized design in the open field (35º82’73.5’’ N, 128º75’51.7’’ E) of Yeungnam University Farm, Gyeongsan-si, Gyeongsangbuk-do, South Korea. Table 1 shows the fresh weights of the bulblets by species and the propagation method just before planting. After planting, they were grown using the conventional bulb cultivation method (RDA, 2021), and the bulbs were harvested when the leaves of the lily plants grown in the field were half dried. The average, maximum, and minimum temperatures from bulblet planting to shoot emergence are shown in Fig. 1.
Table 1.
Fresh Weights before the Planting of Bulblets Obtained through Tissue Culturing, Scaling, and Seeding in Native Korean Lilies
| Species | Common name | Bulblet fresh wt. (g) | ||
| Tissue culture | Scaling | Seeding | ||
| Lilium amabile Palibin | Friendly or Korean lily | 0.4 ± 0.02 | 0.8 ± 0.07 | 1.0 ± 0.09 |
| Lilium cernuum Komar. | Nodding lily | 0.3 ± 0.02 | 0.8 ± 0.08 | 0.5 ± 0.04 |
| Lilium lancifolium Thunb.z | Tiger or Easter lily | 0.2 ± 0.01 | 1.3 ± 0.11 | 0.5 ± 0.05 |
| Lilium maximowiczii Regel.y | Maximowicz’s orange lily | 0.3 ± 0.01 | 1.8 ± 0.15 | 0.7 ± 0.06 |
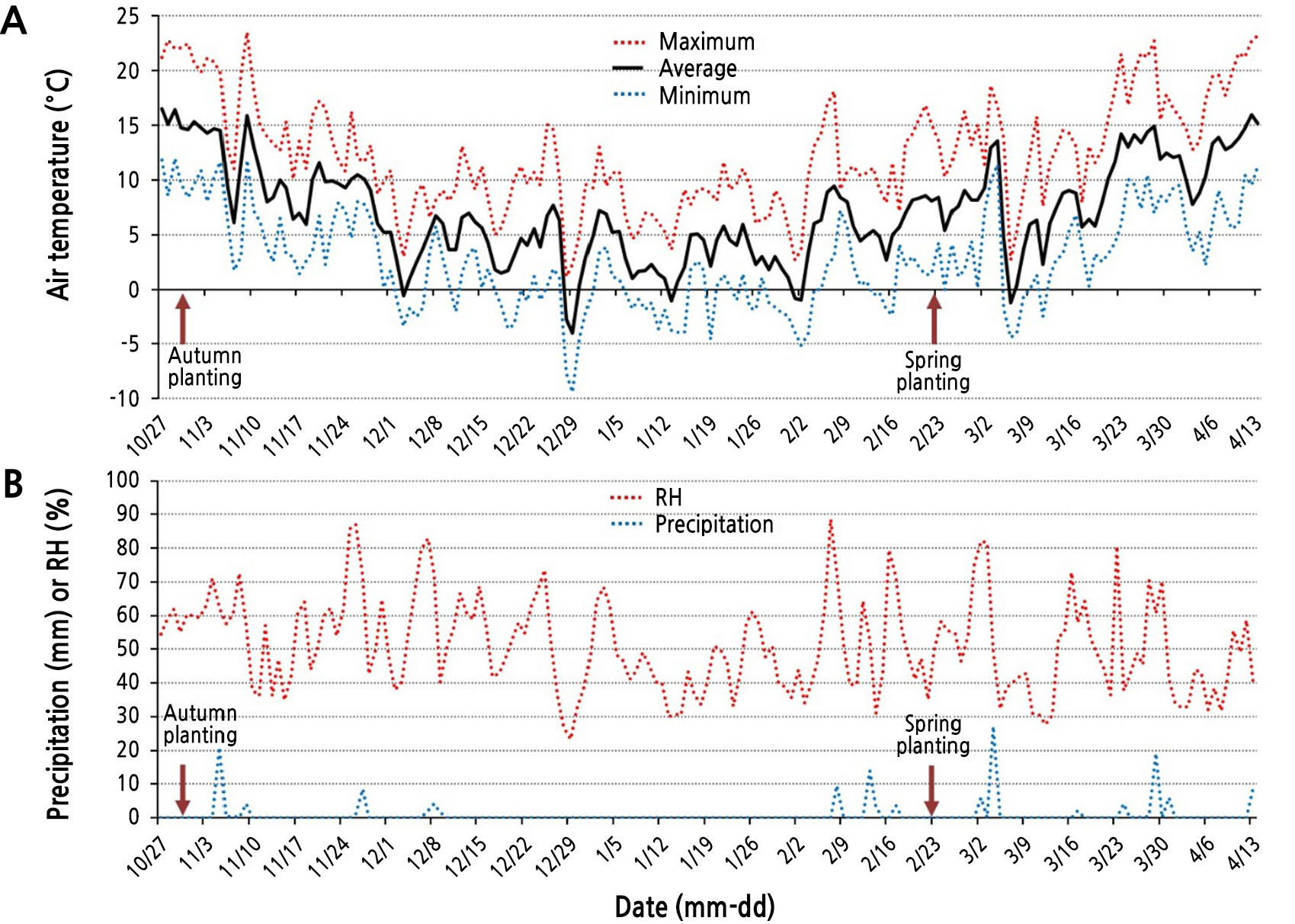
Fig. 1.
Changes in the daily temperature (A), relative humidity (RH) and precipitation (B) from the autumn planting date (October 30, 2016) to the shoot emergence date (April 13, 2017) in this study. The total precipitation from the autumn planting date to February 23 (spring planting date) was 72.7 mm.
Data Collection and Statistical Analysis
The time of shoot emergence was defined as the time shoots with stems or scale-like leaves appeared on the media from the bulblets planted in the autumn and spring and were observable with the naked eye. After the spring planting of the bulblets, the number of bulblets with visible shoots and the date of the shoots were recorded every day for 12 weeks. Using these values, the shoot emergence rate (SER) and average shoot emergence date (SED) were calculated. The SER was expressed as a percentage (%) of the number of bulblets with emerged shoots compared to the number of bulblets planted. The SED was expressed by converting the average date (mm-dd) on which shoots appeared to the day of the year (DOY).
To compare the degree of bulb enlargement by species, the propagation method, and the bulblet planting season (time), the individual fresh weight of each harvested bulb was measured, with this value then divided by the fresh weight before planting (Table 1) to express the bulb enlargement rate (BER). In addition, the correlation between the bulblet fresh weight before planting and the bulb fresh weight (BFW) after harvesting was analyzed. An analysis of variance of the data was performed using SAS (SAS Institute Inc., Cary, NC, USA) and the mean differences in the treatments were separated using Tukey’s honestly significant difference test at a significance level of p < 0.05. A regression analysis was also conducted using SigmaPlot V10 (Systat Software Inc., USA).
Results
SER after Bulblet Planting
The SER of bulblets showed marginally different patterns depending on the species of Korean native lilies, and the propagation method and planting season of the bulblets also had different effects on the SER (Fig. 2). The SER of bulblets derived from tissue culturing was 88% or more regardless of the planting period, but shoots emerged in only 62.5% of bulblets of L. cernuum planted in the fall (Fig. 2). That is, spring planting significantly increased the SER compared to autumn planting only in L. cernuum (Fig. 2B), but there were no significant differences between planting seasons in other species.
For the scaling specimens, one-year-old bulblets planted in autumn and spring showed relatively low SERs (Fig. 2A and 2B) at 50% and 66.7%, respectively, for L. cernuum. The SER of L. amabile bulblets planted in autumn was the lowest at 45% (Fig. 2A). In contrast, the SERs of all other treatment groups were equal to or exceeded 96%. In the comparison between planting seasons within species, the SER of bulblets planted in spring only for L. amabile was higher than those planted in autumn (Fig. 2A).
For seed propagation, shoots appeared in more than 87.5% of seedling-derived bulblets regardless of the species and/or planting period (Fig. 2). In particular, shoots emerged in the bulblets of L. amabile planted in spring and autumn, in L. lancifolium planted in spring, and in L. maximowiczii planted in autumn (SER = 100%). Within the species, there were no differences in the SERs between the planting seasons in any of the four species.
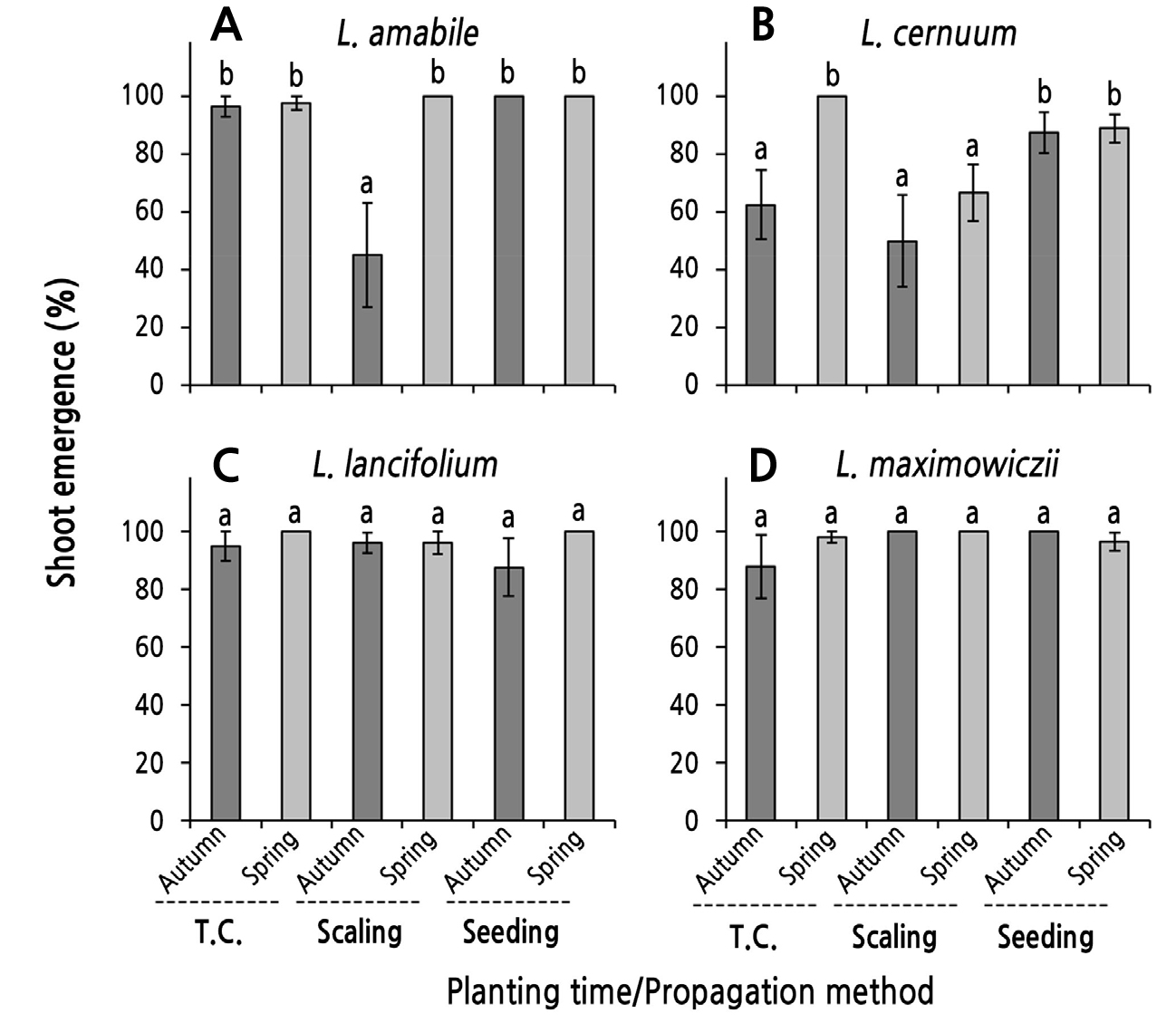
Fig. 2.
Shoot emergence rate (SER) as influenced by the planting season of the bulblets obtained through tissue culturing (T.C.), scaling, and seeding in four species of native Korean lilies. Bulblets were planted on October 30, 2016 (autumn) and were planted on February 23, 2017 after a chilling treatment (spring). Vertical bars represent the standard errors (n = 4). Within-graph means followed by the same letter are not significantly different according to Tukey’s honestly significant difference test at p < 0.05.
SED after Bulblet Planting
The SED of bulblets also differed marginally depending on the species of lilies native to Korea, and the propagation method and planting season had different effects on the SED (Fig. 3). The SED of the tissue culture-derived bulblets was earlier in L. lancifolium than in the other three species. The planting season (time) did not affect the SED in L. cernuum or L. lancifolium (Fig. 3B and 3C). In contrast, for the L. amabile and L. maximowiczii specimens, bulblets planted in autumn appeared 10 to 12 days earlier than those planted in spring (Fig. 3A and 3D).
The SEDs of one-year-old bulblets derived from scaling were earlier in the following order: L. lancifolium > L. amabile ≈ L. maximowiczii > L. cernuum (Fig. 3). Comparing the SED according to the planting season, shoots emerged five days earlier in the bulblets of L. amabile and L. cernuum planted in autumn than those planted in spring, whereas there was no statistically significant difference between planting seasons in the other two species (Fig. 3A and 3B).
The SEDs of bulblets derived from seedlings were earlier in the following order: L. lancifolium > L. amabile > L. maximowiczii ≈ L. cernuum (Fig. 3). The SED of L. amabile was around March 30 (DOY 89) regardless of the planting season. In contrast, for L. cernuum and L. lancifolium, shoots emerged 10 to 11 days earlier in the bulblets planted in autumn than those planted in spring. For L. maximowiczii, autumn planting delayed the SED by eight days compared to spring planting, but there was no significant difference between the planting seasons (Fig. 3D).
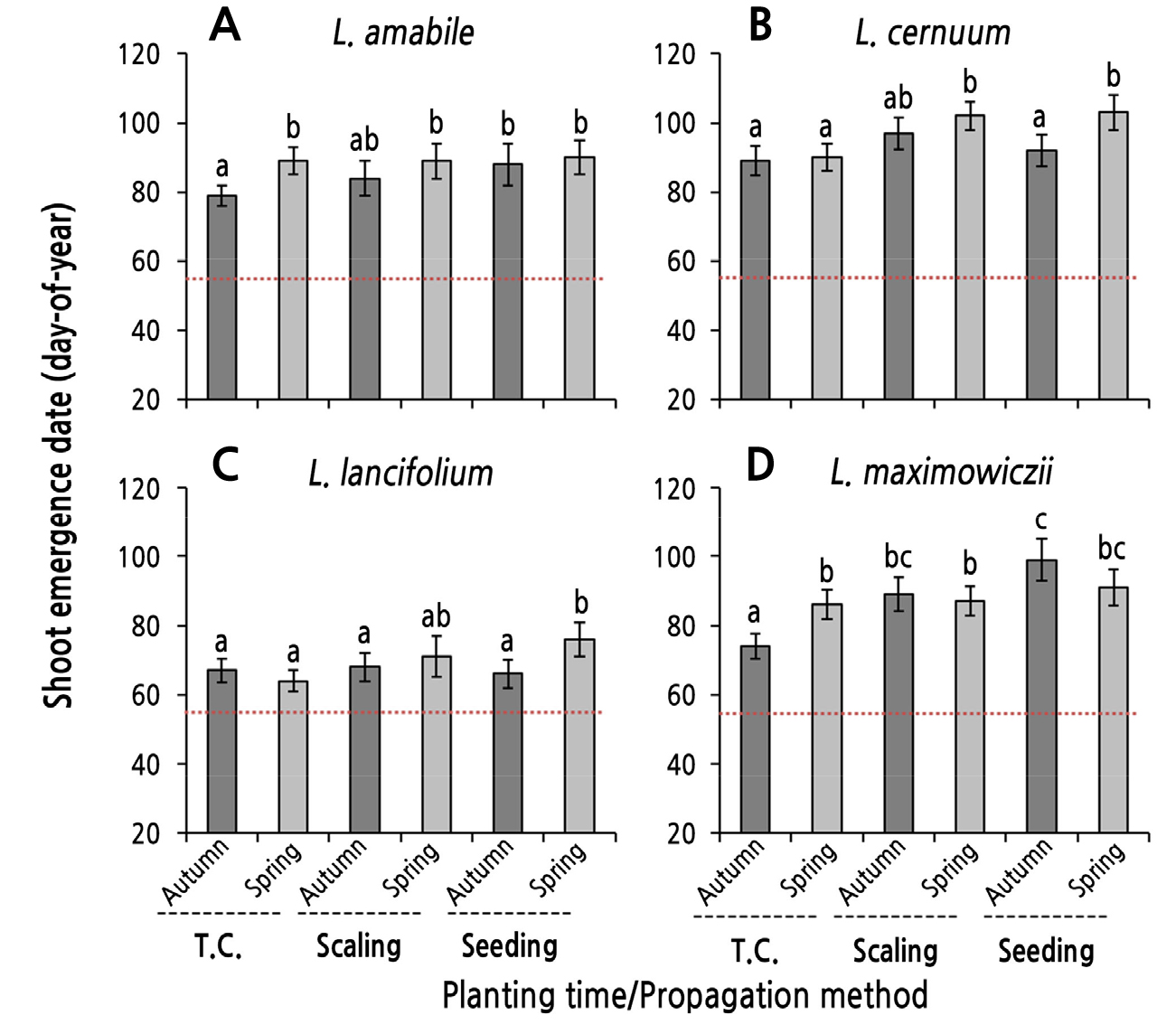
Fig. 3.
Shoot emergence date (SED; day-of-year, DOY) as influenced by the planting season in bulblets obtained through tissue culturing, scaling, and seeding in native Korean lilies. Bulblets were planted on October 30, 2016 (autumn) and were planted on February 23 (DOY 54, red dotted lines), 2017 after a chilling treatment (spring). Vertical bars represent the standard errors (n = 4). Within-graph means followed by the same letter are not significantly different according to Tukey’s honestly significant difference test at p < 0.05.
BFW at Harvest Time
The fresh weight of bulbs harvested after planting bulblets in the open field was affected by the propagation method and the planting season. According to the propagation method, the BFW of L. cernuum was lowest, followed by L. maximowiczii, L. amabile, and L. lancifolium (Fig. 4). When comparing the propagation method within species, L. amabile had heavier bulbs at harvest in the following order: seed propagation > scaling > tissue culture. In L. cernuum, L. lancifolium, and L. maximowiczii, the BFW was higher in the following order: scaling > seed propagation ≥ tissue culture (Fig. 4). The BFW at harvest may be related to the fresh weight of the propagation materials (bulblets) before planting. The fresh weight during harvesting was higher as the weight of the propagation material increased (Fig. 6).
The BFW at harvest according to the planting season of the bulblets tended to be higher in spring than in autumn in all species, but statistically significant differences between planting seasons were found only in some species and propagation methods (Fig. 4). In the tissue cultures of L. cernuum, L. lancifolium, and L. maximowiczii, which had heavier propagation materials, the BFW at harvest for the spring-planted bulblets was significantly higher than that of autumn-planted bulblets (Fig. 4B, 4C, and 4D). In addition, in the bulblets derived from seedlings, the BFW from the spring planting group was higher than that from the autumn planting group in three species but not L. amabile. In contrast, there was no difference between the planting seasons in all species in the tissue-culture-derived bulblets compared to the other propagation methods (Fig. 4).
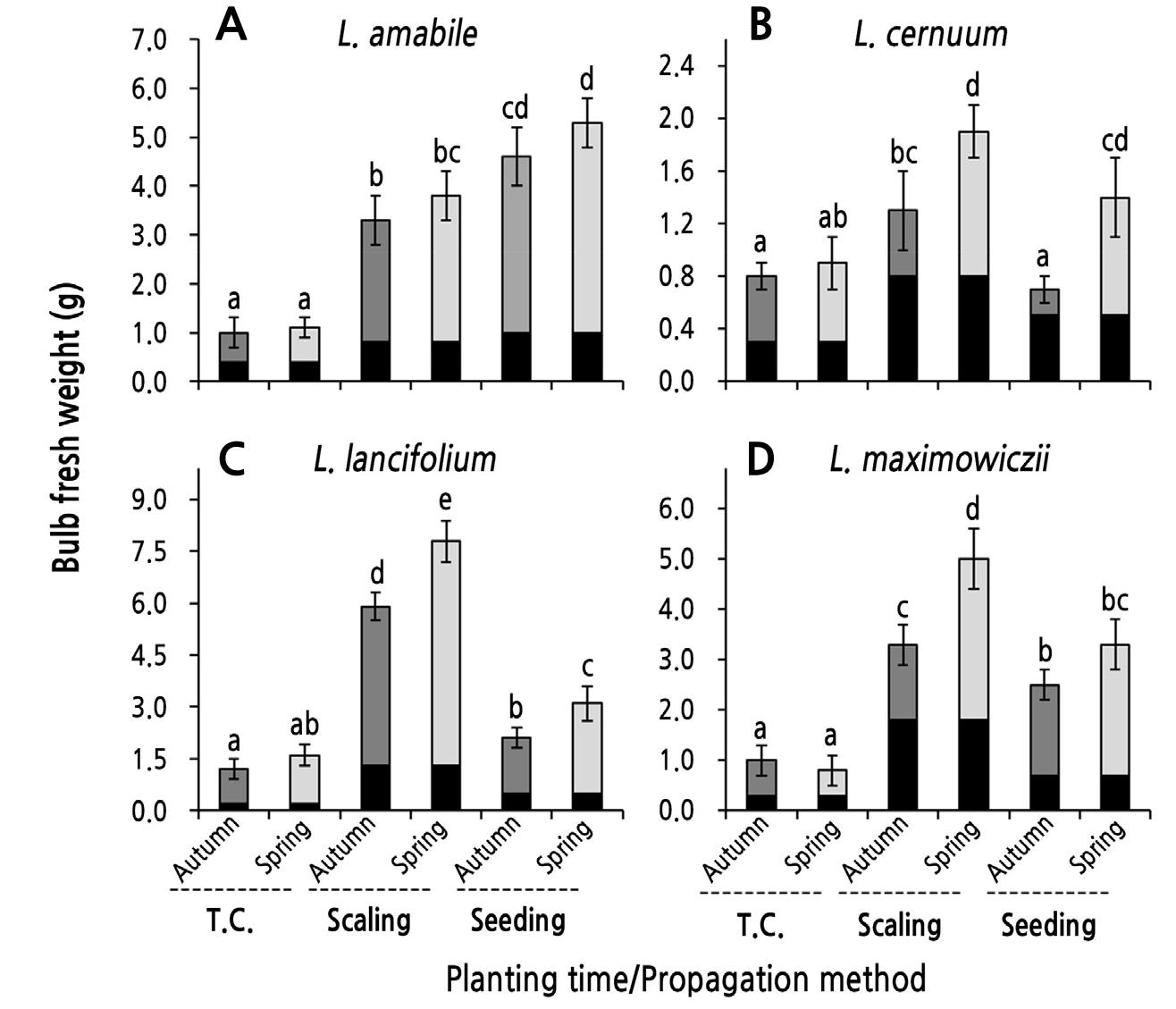
Fig. 4.
Bulb fresh weight (BFW) after harvesting as influenced by the planting season of bulblets obtained through tissue culturing, scaling, and seeding in four species of native Korean lilies. The black parts of the bars represent the fresh weights just before planting. Bulblets were planted on October 30, 2016 (autumn) and were planted on February 23, 2017 after a chilling treatment (spring). Vertical bars represent the standard errors (n = 4). Within-graph means followed by the same letter are not significantly different according to Tukey’s honestly significant difference test at p < 0.05.
BER at Harvest Time
The bulb enlargement rate (BER [%] = [BFW at harvest/bulblet fresh weight just before planting) × 100] was highest overall in the following order: L. lancifolium > L. maximowiczii > L. amabile > L. cernuum, and the BER of L. lancifolium was more than twice that of L. cernuum (Fig. 5). In terms of bulb enlargement according to the planting season, spring planting generally increased the BER compared to autumn planting.
The BER of L. amabile was higher in the following order: seedling > scaling > tissue culture, but there was no difference in the BER between the planting seasons within each propagation method (Fig. 5A). There was also no difference in the BER among the propagation methods in L. cernuum and L. lancifolium (Fig. 5B and 5C). In terms of the planting season, spring planting significantly increased the BER compared to autumn planting, except for the tissue-cultured L. cernuum (Fig. 5). In L. maximowiczii, the BER of spring-planted bulblets derived from scaling and seedlings was higher than that of the autumn-planted case, but there was no difference in the BER between planting seasons of bulblets derived from tissue culturing (Fig. 5D).
Overall, in L. lancifolium, spring planting for all the propagation methods increased the BFW at harvest by about six times compared to the bulblet fresh weight when they were planted, which was the most significant result in terms of the BER (Fig. 5C). In contrast, the BER for autumn planting was less than five times that for spring planting in all propagation methods in L. lancifolium. In L. cernuum and L. maximowiczii, the BERs of bulblets obtained by scaling tended to be low, and the bulblet fresh weight during the planting season was heavier (Fig. 5B and 5D).
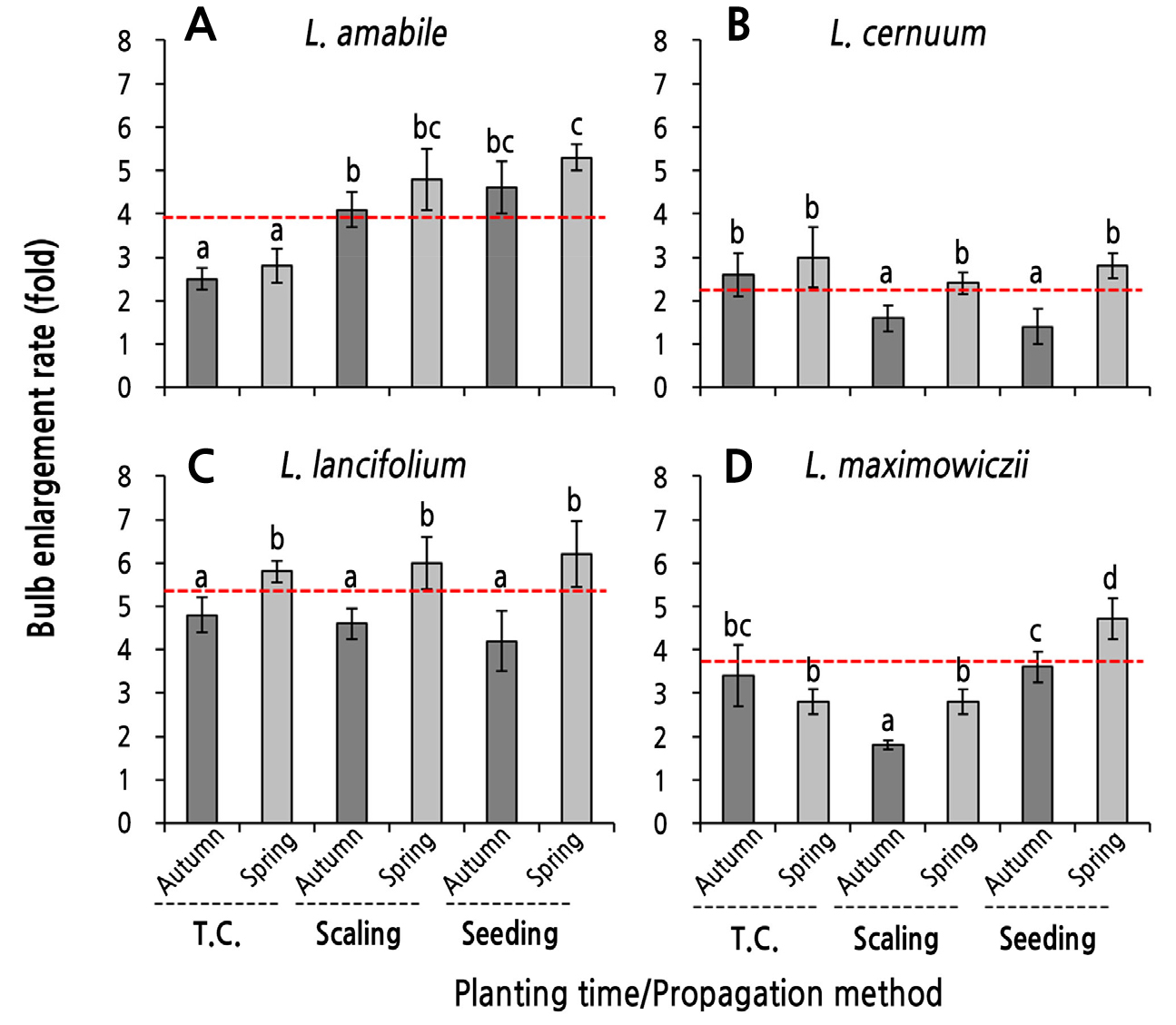
Fig. 5.
Bulb enlargement rate (BER) as influenced by the planting season of bulblets obtained through tissue culturing (T.C.), scaling, and seeding in four species of native Korean lilies. Bulblets were planted at on October 30, 2016 (autumn) and were planted on February 23, 2017 after a chilling treatment (spring). The enlargement rate (fold) = the bulb fresh weight at harvest/bulblet fresh weight before planting. Red lines represent the mean bulb enlargement rates of each species. Data of fresh weights before planting and after harvesting were derived from Table 1 and Fig. 4. Vertical bars represent the standard errors (n = 4). Within-graph means followed by the same letter are not significantly different according to Tukey’s honestly significant difference test at p < 0.05.
Correlation between the Fresh Weights of Bulblets Planted and Bulbs Harvested
A correlation analysis between the BFW just before planting and that immediately after harvesting in the four lilies native to Korea suggested that the relationship was similar for all species (Fig. 6). In particular, L. amabile (y = 6.46x − 1.56, R2 = 0.98, p < 0.001) and L. lancifolium (y = 5.03x + 0.26, R2 = 0.93, p < 0.001) showed a linear positive correlation (Fig. 6A and 6C). In L. cernuum, which had a relatively light bulblet before planting, as the bulblet fresh weight increased, the rate of increase in the BFW after harvesting showed a secondary functional increase (y = 1.67x2 − 0.33x + 0.80, R2 = 0.58, p < 0.05; Fig. 6B). In contrast, L. maximowiczii, which had a relatively large bulblet fresh weight before planting, showed a curve in which the increase in the BFW after harvest decreased as the initial bulblet fresh weight increased (y = −2.58x2 + 7.58x − 1.14, R2 = 0.86, p < 0.001; Fig. 6D).
Based on the data overall from the four wild lilies, the correlation between the fresh weights of the bulblets before planting and the bulbs after harvesting was shown to be a quadratic functional increase (y = −2.52x2 + 8.00x − 1.28, R2 = 0.67, p < 0.01; Fig. 6E). In the range in which the bulblet fresh weight before planting increased from 0.2 g to 1.2 g, the BFW after harvesting increased linearly, with an inflection point appearing at about 1.5 g and the increasing trend slowing. The fresh weight of the bulblets obtained by tissue culturing was lower overall, whereas that of bulblets obtained by scaling was higher (Table 1).
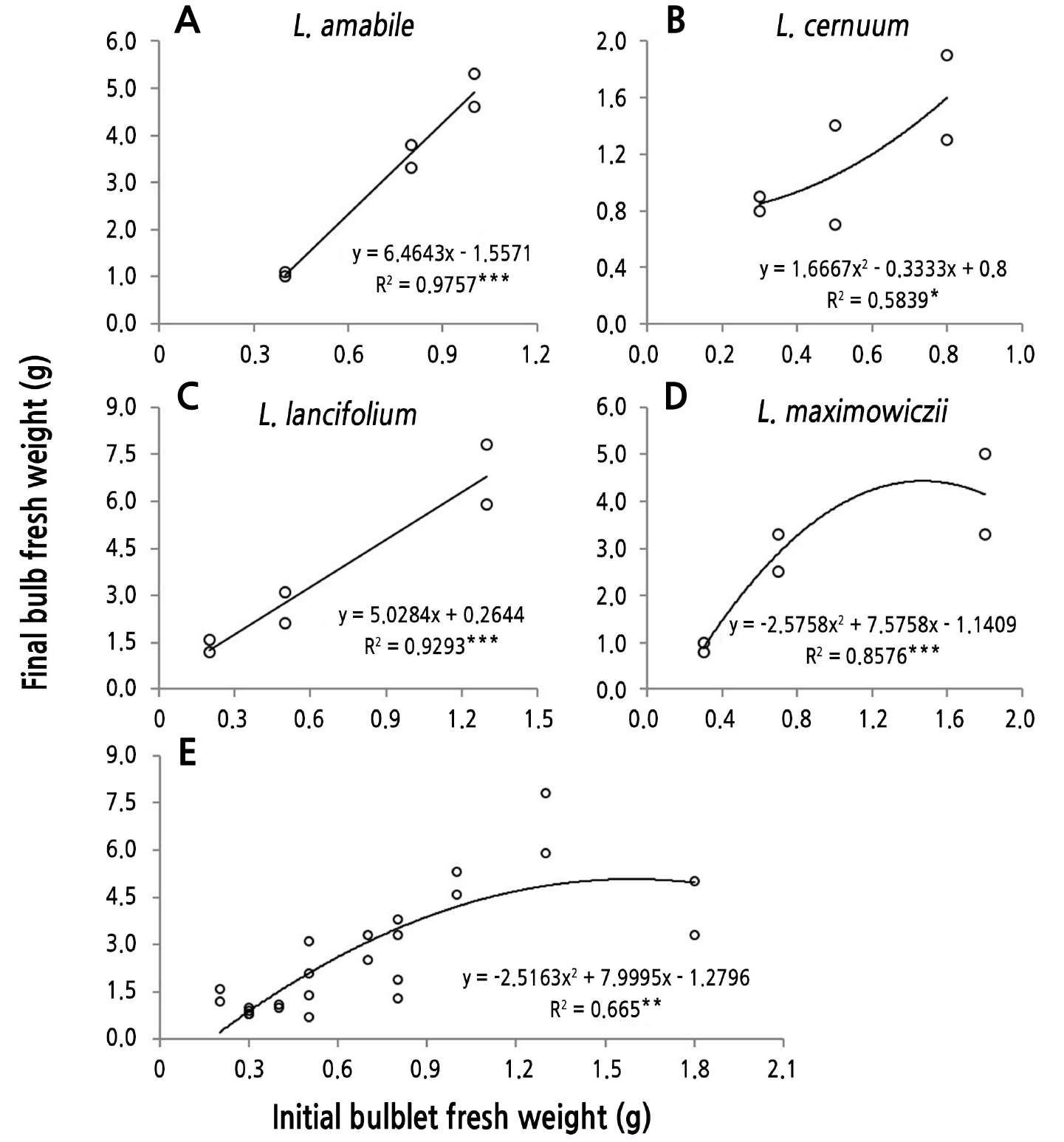
Fig. 6.
Relationships between the initial fresh weights of bulblets (derived from Table 1) and the final bulb fresh weight (BFW) at harvest (derived from Fig. 4) for each species of native Korean lilies (A-D) and the combined values of all species (E). Bulblets were obtained by tissue culturing, scaling, and seeding.
Discussion
In this study, one-year-old bulblets obtained by three propagation methods were planted in an open field in autumn (wintering in the open field) and in the following spring (planting was done after a 0°C wet low-temperature treatment), targeting four species of native Korean lilies belonging to the Sinomartagon section. The shoot emergence rate (SER), shoot emergence date (SED) of the planted bulblets, fresh weight, and the enlargement rate of the harvested bulbs were compared among the treatments. The results suggest that shoot emergence from bulblets and the enlargement of harvested bulbs differ according to the species, propagation method, and planting season (Figs. 1, 2, 3, 4, and 5).
Shoot emergence after sprouting is essential for one-year-old bulblets to start growing and to show enlarged bulbs. In this study, the overall SER was lower in L. cernuum than in other species. In particular, when bulblets obtained by tissue culturing were planted in autumn (62.5%) and those obtained by scaling were planted in autumn (50%) and spring (66.7%), the SER was significantly lower (Fig. 2B). When those obtained by scaling for L. amabile were planted in autumn, the SER was 45%, which was the lowest rate among all treatments for all species (Fig. 2A). However, the species and treatments not mentioned above showed a high SER of more than 87.5% (Fig. 2).
The SER of bulblets derived from the scaling of L. amabile and the tissue culturing of L. cernuum differed between planting seasons, and spring planting significantly increased the SER compared to autumn planting (Fig. 2A and 2B). With regard to the SED, unlike SER, there were no significant differences in these bulblets between planting seasons (Fig. 3A and 3B). Treatment groups with no difference in the SER but a significant difference in the SED between planting seasons consisted of bulblets derived from the tissue culturing of L. amabile and L. maximowiczii and the seedlings of L. cernuum and L. lancifolium. In all of these bulblets, spring planting delayed shoot emergence compared to autumn planting (Fig. 3). However, in bulblets obtained by the seed propagation of L. amabile, the scaling of L. cernuum, the tissue culturing and scaling of L. lancifolium, and the scaling and seed propagation of L. maximowiczii, the planting season did not have a significant effect on the SER or the SED (Figs. 2 and 3).
Thus, there were no significant differences in the SER and SED between planting seasons for all propagation methods for all species. However, the SER and SED for bulblets planted in autumn tended to be low and earlier, respectively, compared to those planted in spring. This is likely due to the temperature and precipitation that bulblets planted in autumn (October 30) encountered in the open field until February 22, which is the spring planting time (Fig. 1). That is, from February 3, 20 days before spring planting, as the temperature rose, there were 15 days with an average temperature of 5°C or higher, while there were only five days in which the minimum temperature fell below freezing during the same period (Fig. 1A). There were 16 days with the highest temperature above 10°C and five days with the highest temperature above 15°C. There was also natural precipitation of 30 mm during this period (Fig. 1B). In other words, it is inferred that the bulblets planted in autumn encountered environmental conditions (e.g., temperatures, precipitation amounts) permissible for shoot emergence at 15 to 20 days while the bulblets for spring planting were still stored in the freezer at 0°C.
On the other hand, it is also possible that some bulblets did not reach the point of shoot emergence because they were exposed to various stressful environments, such as rapid temperature fluctuations and drought in the open field for 116 days from the autumn planting date to the spring planting date (Fig. 1). Autumn is the typically recommended time to plant bulbs for the flowering of Lilium Asiatic hybrids (RDA, 2021). It is also desirable to plant the bulbs in the fall or as early as possible, even in spring, for those in the wild lilies of Sinomartagon section, which are the progenitors of the hybrids. Bulblets planted in autumn take root and grow to a certain extent, naturally enter dormancy due to low temperatures, and after overwintering resume growth in the spring of the following year. Therefore, the enlargement of bulblets planted in autumn is superior to that of bulblets of the same size planted in spring. However, bulblets produced by scaling or micropropagation are small in size and have very low adaptability to weather conditions or soil environments; therefore, careful cultivation management is required (RDA, 2021).
The fresh weight of harvested bulbs also depended on the propagation method and the planting season, with these outcomes differing depending on the species as well (Fig. 4). First, because the fresh weight of bulblets (seed bulbs) just before planting differs depending on the propagation method, it is natural that the fresh weight of the bulb at harvest would vary. However, overall, the fresh weight of bulbs harvested after spring planting was greater than that from autumn planting. In particular, the spring planting of bulblets derived from the scaling and seed propagation of L. cernuum, L. lancifolium, and L. maximowiczii showed significantly increased postharvest BFW outcomes compared to those from the autumn planting (Fig. 4B, 4C, and 4D). In addition, the BER of the bulblets mentioned above was significantly higher in the spring-planted individuals than in the autumn-planted case (Fig. 5B, 5C, and 5D). Additionally, spring planting of bulblets derived from the tissue culture of L. lancifolium significantly increased the BER compared to autumn planting (Fig. 5C). In L. amabile, the BFW and BER for each propagation method were higher in spring planting than in autumn planting, but there was no statistically significant difference between the planting seasons (Figs. 4A and 5A). It is estimated that spring planting is more advantageous for bulb enlargement than autumn planting because bulblets for spring planting can safely overwinter in storage to avoid damage to the bulbs caused by pests, drought stress, and low temperatures during winter in the open field.
In the comparison between the species, the BER was higher in the following order: L. lancifolium > L. amabile > L. maximowiczii > L. cernuum (Fig. 5). L. lancifolium and L. amabile, with relatively large BERs, are reportedly genetically closer than the other species (Song and Lee, 2017). Overall, bulb enlargement of L. cernuum did not match those of the other species. With regard to tissue culturing, L. cernuum was more sensitive to environmental conditions than L. maximowiczii and L. amabile (Eum et al., 2006), and the in vitro bulblet enlargement of L. cernuum was less than those of L. maximowiczii and L. amabile (Eum et al., 2008). It is inferred that the genetic characteristics of this species are expressed in the field growth and bulb enlargement processes.
The fresh weight of the bulblets just before planting and the BFW when harvesting them showed a nearly proportional relationship regardless of the species (Fig. 6). However, the slope decreased as the bulblet fresh weight exceeded 1.5 g (Fig. 6D and 6E). In the Lilium Oriental hybrids ‘Sorbonne’, ‘Siberia’, and ‘Casa Blanca’, the values for the fresh weight and circumference of the bulbs were higher with larger seed bulbs (Kim et al., 2013), as shoots with higher plant heights, leaf numbers, and greater leaf areas grow from larger seed bulbs, and these shoots enlarge the bulb given the greater amount of photosynthesis. In Triteleia laxa ‘Queen Fabiola’, the fresh weight of the planted corms increased as that of the harvested corms increased (Han et al., 1991). In the Tulipa gesneriana ‘Kees Nelis’ and ‘Ile de France’, the BFW at harvest was higher with larger planted bulbs (Yoo and Roh, 2014).
The sprouting percentages of L. cernuum and L. amabile belonging to the Sinomartagon section were 100%, and those of L. lancifolium and L. maximowiczii were also high at 88.0% and 73.0%, respectively, whereas those of L. distichum, L. hansonii, and L. tsingtauense in the Martagon section were relatively low at 34.0–54.0% (Kim and Kim, 2009). In other words, we suggest that seed propagation can be utilized for bulb production because native Korean lilies in the Sinomartagon section have a high germination rate. However, if there are large differences among individuals with regard to the horticultural characteristics of bulbs when bulblets obtained by seed propagation are used as seed bulbs, vegetative propagation methods such as scaling or tissue culturing should be used.
In conclusion, in the bulb production of four Korean native species belonging to the Sinomartagon section - regardless of the propagation method for obtaining bulblets - wet cold storage without the planting of the bulblets in autumn, subsequently planting them in an open field or greenhouse in the spring for intensive management, will be the key to good bulb production. In addition, it is necessary to establish a bulb production system for native lilies given their high ornamental value, to breed hybrid cultivars, and to improve their utilization as garden plants.
