

Introduction
Materials and Methods
Nursery Growth of Mother Plants and Propagation of Daughter Plants
C. fructicola Inoculations
Mineral Nutrient Content and Data Analysis
Results and Discussion
The Growth Characteristics of the Daughter Plants were Influenced by the Type of Substrate
ACR-Inhibiting Effects of the Different Substrates
Mineral Nutrient Uptake of the Daughter Plants
Introduction
‘Seolhyang’ strawberry, a Korean-bred cultivar, accounts for 78.4% of all strawberry cultivars grown in Korea (KREI, 2014). One of the most serious problems with this cultivar is its susceptibility to anthracnose crown rot (ACR), a destructive epidemic caused by a fungal pathogen that commonly occurs in strawberry nurseries (Nam et al., 2014). The ACR pathogen, Colletotrichum gloeosporioides (Nam et al., 1998), which colonizes strawberry leaves, petioles, runners, and crowns, was reclassified as C. fructicola (Nam et al., 2013), while the ACR pathogen that colonizes strawberry fruits was identified as C. acutatum (Nam et al., 2008).
With the recent popularity in forcing cultivation of ‘Seolhyang’ strawberries, there has been an increasing trend in raising transplants in rain-sheltered pots to prevent outbreaks of ACR and to increase early yields, resulting in a rapid increase in the use of growing substrates. There are various substrates available for potted transplants, such as clay sand, coir dust, rice hulls, and mixed media. Due to a wide variety of physicochemical properties in the different substrates, the choice of substrate has a large impact on the quality of the transplants and the occurrence of disease (Park et al., 2015). Although the demand for lightweight substrates has been increasing, the major components, such as coir dust and peat moss, are imported. This means that problems with substrate supply could occur, depending on import-related circumstances. An alternative is rice hulls, an organic material abounding in Korea that has advantages in terms of uniformity, workability, and stable supply. In particular, expanded rice hulls (ERH) have improved physicochemical properties with a distinctive feature of enhanced soil aeration (Kim et al., 2001; Lee et al., 2000). According to previous studies, strawberries grown on ERH-containing substrates showed symptoms of temporary nitrogen deficiency during the maturation process due to the high C/N (carbonto- nitrogen) ratio of ERH-containing substrates, which decreases tissue N contents and accelerates flower bud differentiation in strawberries (Yun, 1996; Hwang et al., 2003).
Son (2009) reported that strawberries took up increased amounts of silicon due to the high concentrations of silicon in ERHs. In addition, Bélanger et al. (1995) noted that the silicon absorbed by strawberry roots moves to the shoot and induces pathogen resistance by stimulating defense mechanisms. Nam et al. (2006) analyzed the relationship between the increase in nutrient uptake by strawberry plants and the occurrence of ACR and reported that increased uptake of nitrogen and potassium increased outbreaks of ACR. Likewise, Long et al. (2000) reported that high nitrogen and potassium concentrations in plants stimulated spore germination and appressorium formation in the rice blast fungus, and that silicate had an inhibitory effect on rice blast disease and cucumber powdery mildew (Bélanger et al., 1995; Seebold et al., 2001).
Due to the increasing popularity of cultivating strawberry transplants on ERH substrates, it is important to provide strawberry farmers with practical guidelines on ERH-based cultivation techniques and to clarify the mineral uptake characteristics of ERH-grown transplants and the ACR-inhibiting effects of ERHs. The aim of this study was to provide field-applicable data on the inhibitory effect that ERH substrates have on ACR in the cultivation of ‘Seolhyang’ strawberry.
Materials and Methods
Nursery Growth of Mother Plants and Propagation of Daughter Plants
The experiments were conducted in a transplant greenhouse of the Buyeo-gun Agriculture Technology Center and an experimental greenhouse at the Nonsan Strawberry Experiment Station in 2013 and 2014. ‘Seolhyang’ strawberry mother plants were grown in elevated beds (height 0.85 m x width 1.8 m) at 18-cm intervals in two rows. The root substrate for mother plant growth was the mixture of coir dust and perlite (4:6, v/v). Plants were fertilized with the Korean standard nutrient solution (N-P-K-Ca-Mg-S = 13-3-6-6-3-3 me∙L-1) two to three times a day at a concentration of 0.50-0.65 dS·m-1 from late March to late July, and then lowered to 0.40-0.50 dS·m-1 for the remainder of the growing season.
Four different types of substrate were used for daughter plant growth in this experiment: expanded rice hull (ERH) with the improved physical and chemical properties produced by compression, expansion, and grinding of the rice hulls to particle sizes of 1.6 mm or less (Daewon Industry Inc., Chilgok, Korea); commercial substrate for strawberries (CSS) (Seoul-bio Co., Ltd., Eumseong, Korea); clay sand (CS; the 85% of particles were in the size of 1.49 to 2.85 mm); and loamy sand (LS). The pH of the substrates was 7.1, 6.0, 6.5, and 6.6 and the electrical conductivity (EC) was 0.26, 0.19, 0.10, and 0.22 dS·m-1, respectively.
The mother plants planted on March 20th started to produce runners beginning around May 10th and daughter plants in early July. Irrigation began on July 8th to encourage root development of the daughter plants. The daughter plants were severed from the mother plants by cutting the runners on August 5th. The average temperature in the greenhouse was maintained at 25.5°C (range: 21.0-34.5°C), and the photosynthetic photon flux was maintained at 312 μmol·m-2·s-1. On August 20th, the daughter plants were transferred to 24-holed seedling pots (Hwa-Seong Industry Inc., Okcheon, Korea) containing the four different substrates and growth characteristics, including plant height, number of leaves, leaf area per plant, crown width, number of primary roots, root weight, and fresh weight, were measured. Leaf area was measured using a portable area meter (LI-3100, LI-COR Inc., USA).
C. fructicola Inoculations
Seventy-two daughter plants in the three- to four-leaf growth stage were collected from the different substrates for C. fructicola inoculations. The selected daughter plants were transferred to the ACR experimental greenhouse in the Nonsan Strawberry Experiment Station. On August 23rd of both years, each daughter plant was spray-inoculated with 1 mL of a conidial suspension (1 x 105 spores/mL) of C. fructicola CGF100713 obtained from the Nonsan Strawberry Experiment Station. The inoculated daughter plants were incubated under high-temperature (28°C) and high-humidity (higher than 90% in relative humidity) conditions for 48 h. The inoculated daughter plants were then placed in a greenhouse maintained at temperatures ranging from 25°C to 30°C and a relative humidity of greater than 80%. The placement of each of the three replicates of the four substrates was randomized. Additional fertilizer was applied to each plant once a week with irrigation throughout the experimental period in 100 to 150 mL of a 2,000 X dilution of Ajuro Combi 100 (N-P2O5-K2O = 30-10-10) + 2MgO + trace elements (DOFAGRO Ltd., Pyeongtaek, Korea). The incidence of ACR was measured seven times in 2013 (once a week from September 20th to October 31st) and nine times in 2014 (once a week from September 3rd to October 31st). The degree of ACR was measured on a disease index scale of 0-4 (0 = no symptoms; 1 = symptoms on less than 50% of the petioles and leaves; 2 = symptoms on greater than or equal to 50% of the petioles and leaves; 3 = wilt; 4 = dead) (Nam et al., 2006).
Mineral Nutrient Content and Data Analysis
To investigate the changes in the mineral nutrient contents of the plants from each substrate treatment, the total amount of nitrogen (TN), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), and silicon (Si) were analyzed in plant tissue. Mineral analysis was conducted three times in 2014 at 15-day intervals (July 28th, August 13th, and August 29th). In each sampling date, the newly fully expanded third leaves and petioles of daughter plants in each treatment were collected and analyzed. Mineral content analysis was performed in compliance with the Agricultural Science and Technology Research & Analysis Standards specified by the Rural Development Administration (RDA, 2003). Duncan’s multiple range test (p < 0.05) was conducted for the mean comparisons using SAS 9.2 (NC, USA)
Results and Discussion
The Growth Characteristics of the Daughter Plants were Influenced by the Type of Substrate
The daughter plants grown on the CSS substrate had the highest values for plant height, leaf number, leaf area, crown diameter, and fresh weight (Table 1). Plants grown on the CS substrate showed the lowest values for plant height, leaf area, crown diameter, and fresh weight. Plants grown on the ERH and LS substrates showed a similar growth of aerial part. ERH and CS substrates yielded higher values for primary root number and root weights over the CSS and LS substrates.
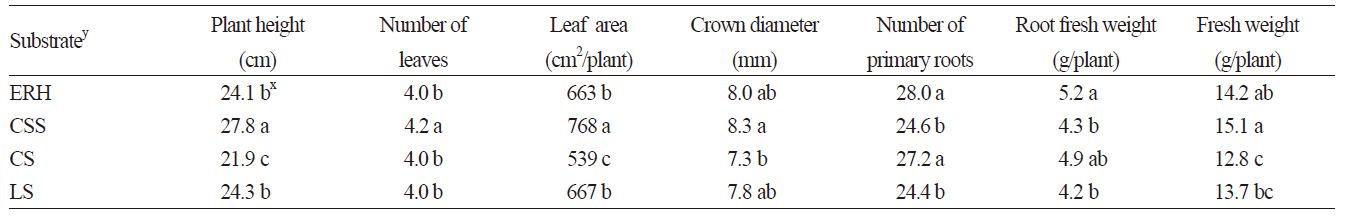
Lee et al. (2000) reported obtaining high-quality fruit and vegetable plants with low leaf nitrogen content and excellent root systems by growing them on an ERH substrate and credited these qualities to the high C/N ratio and porosity of the ERH substrate. In our study, the daughter plants grown on the CSS substrate showed excellent growth of the aerial parts, presumably due to the high concentrations of NH4-N and NO3-N in the CSS substrate, but their roots showed limited development. The high concentrations of NH4-N and NO3-N in CSS substrate was reported by Park et al. (2015) who carried out the experiments using same kinds of substrates in raising daughter plants of strawberry. This is consistent with previous findings that the higher the nitrogen concentration in the root zone, the lower the root yield (Marschner, 2012; Kim et al., 2012). Additionally, the physicochemical properties of the substrates are thought to have had significant effects on the growth of the daughter plants in our previous study (Park et al., 2015) and this study.
ACR-Inhibiting Effects of the Different Substrates
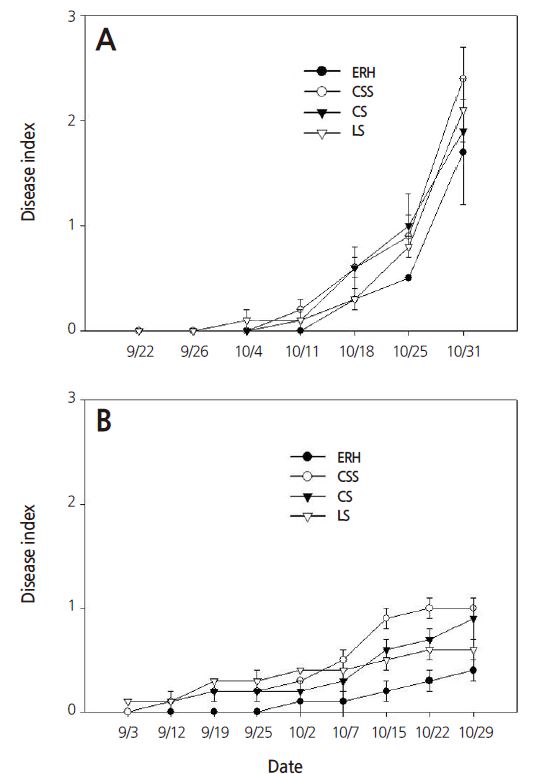
The disease index of the daughter plants grown on the CSS substrate was the highest among all of the substrate treatments in both years (Fig. 1). Those grown on the CS and LS substrates showed the lowest incidence of ACR, but no statistical differences were observed among these three treatments. By contrast, the plants grown on the ERH substrate showed the slowest disease progress on October 11, 2013 and October 7, 2014, differing significantly from plants grown on the other three substrates. While the incidence of ACR rapidly progressed after October 11, 2013, the mortality rate remained at a low level until late October, 2014. Nam et al. (2011) reported that the mortality rate varies greatly depending on weather conditions. During the experimental period in our study, there were many hot and humid days between mid-September and mid-October in 2013, whereas the weather in 2014 was relatively mild (data not shown).
Mineral Nutrient Uptake of the Daughter Plants
The mineral contents of the daughter plants were analyzed three times at 15-day intervals starting from July 28, 2014. Plants from the CSS substrate treatment were found to have the highest nitrogen content with 1.0 to 1.3% nitrogen (Table 2). The ERH, CS, and LS substrate treatments did not show any significant differences in nitrogen content on July 28th. However, on August 13th and August 29th, plants from the ERH treatment had a nitrogen content of 1.02 and 0.85%, respectively, which was lower than that of plants subjected to the other treatments. All of the plants from the substrate treatments tended to show a decrease in nitrogen content as the measurement period progressed. Phosphorus and potassium contents were the highest in the CSS treatment, followed by the LS, ERH, and CS treatments and this trend continued throughout the measurement period. The daughter plants grown on the CSS substrate had a low calcium content, but no differences were seen in calcium content in the other three treatments, and magnesium content was not elevated in any of the treatments. Silicon content was much higher in the plants from the ERH substrate treatment compared to the other treatments.
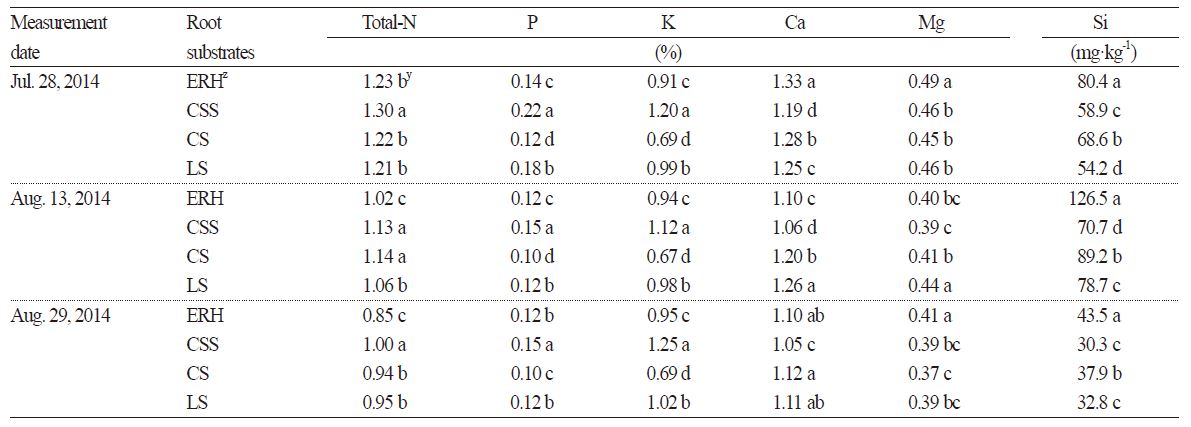
Yun (1996) reported that ERH composted for 60 days lost about 8.0% of the organic materials, which could explain the nitrogen deficiency symptoms seen in the plants grown on the ERH substrate. Kim et al. (2001) noted that ERH-based hydroponic tomatoes showed a sharp decrease in NO3 concentration due to the high C/N ratio of the substrate, which persisted for 20-25 days after the initial delivery of nutrient solutions. In our study, the CSS substrate, in which coir dust is a major of component and a large amount of NH4-N and NO3-N were incorporated during root substrate formulation as pre-planting nutrient charge fertilizer, was thought to accelerate the incidence of ACR due to the increase in the plants’ uptake of nitrogen and potassium caused by continuous potassium elution from the coir dust (Nam et al., 2006). In contrast to the results of our experiment, Park et al. (2015) analyzed the mineral content of transplants grown on the same four substrates and reported that the transplants grown on the CSS substrate had a high calcium and magnesium content and that the nitrogen content of the ERH treatment was similar to that of the other treatments. This result may be due to the fact that the daughter plants were severed from the mother plants 15 days earlier than in our experiment. Kim et al. (2012) noted that the mineral content in the leaves of daughter plants varied widely depending on the time that runners were cut; the later the runner-cutting took place, the higher the nutritional dependence on the mother plants. The slight differences between the findings in our study and Park et al. (2015) regarding the substrate-dependent growth status of the daughter plants may be due to be the differences in the runner-cutting time and the growth monitoring period. The contents of phosphorus, calcium, and magnesium in all of the four substrate treatments was within an acceptable range when checked against the optimal standards proposed by Jeong (2007) for ‘Maehyang’ strawberry (P = 0.30-0.75%, Ca = 1.60-2.25%, Mg = 0.30-0.65%). The concentration of silicon in the plant tissues of ERH treatment remained high until August 13, but then decreased 30 days after the daughter plants were disconnected from the mother plants by cutting of runners. Jun et al. (2006) reported that silicon uptake rapidly increased one month after a soil drench application of silicon during hydroponic cultivation of strawberry. The mechanisms of silicon uptake need further investigation.
From these results, we propose that the markedly lower incidence of ACR noted in the daughter plants grown on the ERH substrate may be due to the low nitrogen content of the ERH substrate (Nam et al., 2006) and the high silicon uptake that may have activated defense mechanisms (Bélanger et al., 1995). The results of this study may be extended to a more in-depth investigation into the mechanisms of mineral uptake in ERH-grown strawberry transplants.
