

서 언
재료 및 방법
작물의 재배관리
생육 조사
생육량 비선형 및 선형 회귀분석
Gompertz 생장함수
지수선형 생장함수(expolinear growth function)
통계분석
결과 및 고찰
서 언
국내 스탠다드 절화 국화는 재배면적, 생산액, 판매량 등에서 국내 화훼시장의 중요한 위치를 차지하고 있다(MAFRA, 2020). 국내에서 재배되고 있는 스탠다드 절화 국화의 대표 품종은 ‘백선(Iwanohakusen)’, ‘신마(Jinba)’, ‘백마(Baekma)’가 있다(Pak et al., 2012; Yoo and Roh, 2015). 특히 ‘백마’는 하추국으로 기존의 품종에 비해 절화수명이 긴 특징으로 주로 수출재배에 이용된다(Choi et al., 2009; Yoon et al., 2014; Roh and Yoo, 2021). ‘백선’은 하국 또는 하추국으로 무측지성 특징을 가지고 있어 생력 재배에 적합한 품종으로 알려져 있다(Shin et al., 2005; Hisamura et al., 2016). 스탠다드 국화의 외형적 품질 요소는 꽃의 색깔, 절화장, 생체중 등이 있다(Karlsson and Heins, 1994, Kim et al., 2012; NAQS, 2020). 국화의 절화장은 절간의 수와 평균 절간장에 의해 결정되고(Pearson et al., 1995), 생체중은 광합성량과 수분함량에 의해 영향을 받는다(Kiehll et al., 1992). 국화의 발육량은 마디와 마디 사이를 의미하는 절간(internode)이 출현하기 위한 발육률인 절간 출현율과 최대 절간 수와 관련이 있으며, 절간 출현율은 재배 온도에, 최대 절간 수는 영양생장 기간에 의존하게 되므로 국화의 발육량은 온도와 장일처리 기간의 조합에 의해 영향을 받게 된다(Carvalho et al., 2002; van der Ploeg et al., 2007). 생체중은 재식밀도와 재배환경인 근권부 수분함량, 수광량 증가에 영향을 받는다(Kiehll et al., 1992; van der Ploeg et al., 2007; Lee et al., 2009). 국내 절화 국화에 관한 주요 연구는 절화품질 및 수명(Yoo and Roh, 2012; Roh et al., 2018), 개화 시기 조절(Kim et al., 2006; Roh and Yoo, 2021) 등에 관한 연구가 수행되었지만, 재배과정에 따른 생육환경 정보 및 생육량의 동적 변화 특성에 대한 연구는 미흡한 실정이다. 특히 스탠다드 국화 ‘백마’와 ‘백선’은 봄 또는 초여름에 정식하여 여름과 가을에 채화하는 특정 시기를 중심으로 연구되었고, 적정한 생육상태를 유지하고 시장규격에 적합한 절화 생산을 위해 품종, 생육단계, 정식시기 등 다양한 환경조건에서 국화의 동적 생육특성에 대한 연구가 요구되고 있다(Yoo and Roh, 2012, 2014; Kwon et al., 2014; Roh and Yoo, 2016; Yoo et al., 2016; Kim et al., 2020).
국화의 동적 생육에 대한 해석 및 정량화를 위해 묘사적 모델 sigmoid 생장모델인 Gompertz 생장함수, Richards 생장함수, Logistic 생장함수와 지수선형(expolinear) 생장 모델을 이용한 연구가 수행되었다(Goudriaan and van Laar, 1994; Lee et al., 2003; Cha et al., 2014; Kim et al., 2020). 묘사적 생장모델은 시계열에 따른 식물체의 생육량을 해석할 수 있는 파라미터를 추정할 수 있고, 파라미터는 식물의 다이내믹한 생육에 대해 상대 생장률, 최대 생장률 도달한 정식 후 일 수, 절대생장률, 선형적 생장으로 전환되기 위해 필요한 일수를 알 수 있다(Goudriaan and Monteith, 1994; Thornley and France, 2007). 재배 환경조건 아래 작물의 동적생육에 관여하는 생육 파라미터 분석을 통해 스탠다드 하국·하추국 절화 국화의 생육과정을 해석하는 것은 국화의 안정적 생산을 위한 작물 생장 모델의 기반 자료로 사용될 수 있다(Lee, 2002; Lee et al., 2009). 최근 시계열에 따른 식물체의 생육 변화량, 재배환경자료 수집, 생육과정 중 이미지 자료 등의 빅데이터를 이용하여 생장량 예측, 품질 예측을 위한 딥러닝 기반 학습모듈 개발이 이뤄지고 있다(Na et al., 2017; Yang et al., 2019; Kim et al., 2020; Lee et al., 2020). 이에 대한 기반 자료로 온실의 환경과 식물의 생육은 정적 변수가 아닌 동적 변수에 의해 식물체의 다이내믹 생육 반응에 대한 연구가 활용될 수 있다(Karlsson and Heins, 1994; Lee et al., 2009). 따라서 본 연구는 스탠다드 절화 국화 ‘백마’와 ‘백선’을 동일한 환경조건에서 시계열에 따른 생육 과정을 요약 및 설명하기 위해 생육 함수 Gompertz 생장모델과 지수선형(expolinear) 생장모델을 이용하여 생육량을 추정하고, 각 모델의 생물학적 의미를 가지는 파라미터를 이용하여 품종별 생육 특성을 구명하며, 작물 생장 모형과 빅데이터 분석에 필요한 동적 생육 자료를 확보하고자 수행되었다.
재료 및 방법
작물의 재배관리
작물 생장 모형 분석과 빅데이터 분석에 필요한 스탠다드 절화 국화(Chrysanthemum morifolium) 의 동적 생육자료를 확보하기 위하여, ‘백마’와 ‘백선’을 연동형 플라스틱 온실에 상시 장일처리(16시간 일장)한 모주에서 삽수를 채취하여 단동 플라스틱 온실(7.0m × 12.5m) 내 설치되어 있는 6개의 재배 베드(베드규격: 높이 20cm × 폭 70cm × 길이 900cm)에 펄라이트(1–2mm, KYUNGDONG ONE, Korea)를 충진하고 포습한 후 2019년 8월 27일 64.6주/m2로 삽목하여 발근될 때까지 간헐적으로 두상살수 하였다. 재배기간 중 온실 내부의 평균 광 투과율은 65%였으며, 온실 내 설치된 주광색(6,500K)과 전구색(3,000K) LED(10W, ELBA, China)를 이용하여 식물체 정단부의 광도를 6.5 ± 0.7µmol·m-2·s-1 이상으로 유지하여 삽목일로부터 50일 간 일장을 16시간(4시–20시)동안 장일처리 하였고, 이후 재배 종료일까지 12시간 30분(19시–7시 30분)동안 암막 스크린(다겹부직포, 9m × 12m)을 이용하여 단일처리 하였다. 단일처리에 따른 국화의 화아분화 이후 꽃목의 신장을 억제하고자 삽목 후 80일 Daminozide을 400배로 희석하여 처리하였다. 양액공급은 삽수가 발근된 이후인 삽목 후 7일부터 일출 30분 후부터 15시 30분까지 외부 광량이 100J·cm-2마다 점적테이프(드리퍼 간격 10cm, 점적테이프 4줄/베드)를 이용하여 EC 2.0dS·m-1와 pH 6.0의 양액을 드리퍼당 35cc(656cc·m-2)의 양으로 양액기(Magma-1000, Green CS, Korea)를 설정하여 공급하였다.
온실 내부의 환경조절은 온실의 양측창과 배기팬을 이용하여 온실 내부의 높은 온도와 습도를 관리하였고, 시설 내부의 낮은 온도는 전기 온수보일러 시스템의 온수(20°C–50°C)를 FCU(10,000kcal·h-1, 5대)로 보내 난방하여 조절하였다. 온실 내부의 CO2 농도는 액화 탄산가스를 이용하여 조절하였으며, 재배 기간 동안 주간 평균 338 ± 24ppm 이었다. 각 시스템의 작동은 외부 환경과 내부 환경을 고려하여 복합환경조절 시스템(Magma, Green CS, Korea)으로 조절하였으며, 일적산광량과 외부온도, 온실 내부의 온도와 습도 및 CO2 농도는 Magma에 저장된 값을 이용하였다(Fig. 1).

생육 조사
실험구의 배치는 난괴법으로 3반복 하여 온실 내부의 6베드 중 3베드씩 ‘백마’와 ‘백선’을 삽목하였다. 삽목일을 정식일로 보아 각 베드 당 국화의 발근이 이루어진 정식 후 14일 부터 수확 종료일까지 7일 간격으로 6주의 식물체를 샘플링하여 초장, 절간 수, 엽 수, 엽면적, 엽, 엽병, 줄기, 화기 등 개별 측정된 기관들의 무게의 합인 총생체중과 총건물중을 측정하였다. 각 처리구의 마지막 생육 조사는 개화 과정까지의 국화의 생육 특성을 파악하고자 설상화의 바깥 꽃잎이 수평적으로 펼쳐진 만개 상태인 2019년 12월 5일(‘백선’)과 2019년 12월 12일(‘백마’)에 조사하였다.
엽면적은 엽면적계(LI-3100C, LI-COR Inc., USA)를 이용하였고, 기관별 건물중은 농산물건조기(KED-132A, Kiturami, Korea)를 이용하여 70°C에서 72시간 이상 완전 건조한 후 실온에서 측정하였다.
생육량 비선형 및 선형 회귀분석
스탠다드 절화국화 ‘백마’와 ‘백선’의 동적 생육 특성을 분석하기 위해 정식 후 일수에 따른 초장, 엽면적, 절간 수, 총생체중을 비선형 회귀식 Gompertz 생장모델과 expolinear 생장모델을 이용하였다. Gompertz 생장모델(Gompertz, 1825; Park and Lee 2021; Eqns. 1 and 2)을 이용하여 정식 후 일 수(x)에 따른 초장, 식물체 엽면적과 절간 수의 최대 값(ymax; yph, yla, yi), 상대생장률(k; kph, kla, ki, d-1), 최대생장률에 도달한 정식 후 일 수(tinf; tph, tla, ti, d), x가 -∞일 때 y값(intercept; y0; y0_ph, y0_la, y0_i)을 추정하여 두 품종을 분석하였다.
Gompertz 생장함수
‘백마’와 ‘백선’의 절대 생장률을 분석하기 위해, 정식 후 일수에 따른 초장, 엽면적과 절간 수를 이용하여 추정한 Gompertz 생장 모델의 파라미터들을 추정하였고, 도함수(Eqn. 2)를 통해 줄기 신장률(stem elongation rate, cm·d-1), 엽면적 확장률(leaf area expansion rate, cm2·d-1), 절간 출현율(internode appearance rate, nodes·d-1)을 분석하였다.
정식 후 일수(x)에 따른 총 생체중은 지수선형 생장함수(Goudriaan and van Laar, 1994; Eqn.3)의 매개 변수인 최대 절대 생장률(cm, g·plant-1·d-1), 최대 상대 생장률(rm, d-1), 선형적 생장으로 전환되기 위해 필요한 일수(tb, d)를 분석하였다.
지수선형 생장함수(expolinear growth function)
정식 후 일수에 따른 생체중의 절대 생장률의 변화는 추정된 지수선형 생장함수의 파라미터들을 도함수(Eqn. 4)를 통해 정식 후 일수에 따른 품종별 총생체중 생장률(plant growth rate, g·plant·d-1)을 분석하였다(Goudriaan and Monteith, 1994). 초장의 신장량에 따른 총생체중 증가량의 생장패턴 분석을 위해 선형모형을 이용하여 분석하였다.
통계분석
최종 생육 조사를 한 ‘백마’와 ‘백선’ 두 품종의 생육 특성 분석은 화기를 제외한 영양기관의 생장량(vegetative organs dry weight, g), 측정된 초장을 줄기 생체중으로 나눈 비경장(specific stem length, cm·g-1), 비경장의 역수인 비경중(specific stem weight, g·cm-1), 개별 엽면적을 엽건물중으로 나눈 비엽면적(specific leaf area, cm2·g-1)을 분석하였다. 동적 생육 특성을 분석하기 위해 정식 후 일수에 따른 초장, 엽면적, 절간 수, 총 생체중을 비선형 회귀식 Gompertz 생장모델과 expolinear 생장모델을 이용하여 각 함수의 파라미터를 통계처리 프로그램(SPSS Statistics, IBM, USA)의 비선형 회귀분석으로 추정하였고. 초장의 신장량에 따른 총 생체중의 생장량 증가는 선형 회귀식으로 분석하였다.
결과 및 고찰
정식 이후 장일처리기간 동안 일적산광량은 12.33 ± 6.16MJ·m-2·d-1, CO2 농도 365.7 ± 15.6ppm, 일평균 온도 22.2 ± 3.2°C, 일평균 RH 86.6 ± 5.54%로 일적산광량은 높은 표준편차로 변화가 심하였으나, 시설 내부의 환경은 일정하게 유지되었다. 단일처리 기간 동안 일적산광량은 10.55 ± 3.67MJ·m-2·d-1, CO2 농도 402.7 ± 25.74ppm, 일평균온도 19.54 ± 1.08°C, 일평균 RH 87.1 ± 4.33%로 온실 내부의 환경의 변화폭이 안정적으로 관리되었다(Fig. 1).
동일한 환경조건에서 재배된 ‘백마’, ‘백선’의 최종 생육 특성은 서로 상이하였다(Table 1). 단일처리 후 출뢰소요일과 개화소요일은 ‘백마’는 ‘백선’ 보다 각각 5일, 7일 지연되었다. 총생체중과 총건물중은 ‘백마’가 ‘백선’보다 각각 72, 45% 무거웠고, 단위 줄기 무게 당 줄기의 길이를 나타낸 비경장(SSL)의 역수는 줄기의 강도를 간접적으로 나타낼 수 있는 지표로서(Poorter et al., 2012; Wyka et al., 2019), ‘백마’는 ‘백선’보다 46% 더 높은 강도를 나타냈다. 이는 화성유도와 화아분화가 지연된 ‘백마’가 ‘백선’ 품종보다 영양생장기간이 길어져 만개까지의 정식 후 소요일수가 7일 지연되었으며, 최종생육량에 미치는 결과로 판단된다. 비엽면적(SLA)은 엽의 두께를 간접적으로 나타낼 수 있고(Johkan et al., 2010; Lee and Kim, 2014; Song et al., 2014), 품종에 따른 차이는 없었다(Table 1).
Table 1.
Comparison of budding and flowering by days after the short-day treatment and final characteristics of the plant height, number of internodes, average internode length, leaf area (LA), total fresh weight (TFW), total dry weight (TDW), vegetative organs dry weight (VDW), specific fresh stem length (SSL), specific fresh stem weight (SSW), and specific leaf area (SLA) for the standard chrysanthemums ‘Baekma’ and ‘Iwanohakusen’ from 27 Aug 2019 to 12 Dec 2019
| Cultivar |
Days to visible flower budz |
Days to floweringy |
Plant height (cm) |
Number of internodes |
Average internode length (cm) |
LA (cm2·plant-1) |
TFW (g·plant-1) |
TDW (g·plant-1) |
VDW (g·plant-1) |
SSL (cm·g-1) |
SSW (g·cm-1) |
SLA (cm2·g-1) |
| ‘Baekma’ | 23x | 47 | 93.6 | 47 | 2.0 | 770.5 | 74.7 | 10.5 | 9.1 | 4.5 | 0.23 | 252.5 |
| ‘Iwanohakusen’ | 18 | 40 | 79.5 | 45 | 1.8 | 550.3 | 43.5 | 7.2 | 6.2 | 6.6 | 0.16 | 246.8 |
스탠다드 절화 국화는 단일조건에서 화기형성이 되는 대표적인 유한생장형 작물로서 ‘백마’, ‘백선’ 두 품종 모두 장일처리기간에서 줄기의 신장과, 엽면적의 확장 그리고 절간의 출현은 정식 이후 낮은 생장률을 나타낸 후 생장률이 급속히 증가한 뒤 단일처리로 인한 화성유도 및 화아분화 이후 완만하게 증가하는 sigmoid형의 신장 패턴을 보였다(Figs. 2 and 3). 절화 국화의 초장과 생체중은 시장의 주요 등급 지표이며(Karlsson and Heins, 1994; Kim et al., 2012) 이는 생육 과정에서 큰 영향을 받는다. 국화의 작물 생장 모형과 동적 생육 특성 분석을 위하여 품종 별 국화의 초장 신장량, 엽면적 확장량, 절간 출현수 그리고 생체중 증가량의 해석이 필요하여, S 생장모델인 Gompertz 생장 함수와 지수생장과 선형생장을 나타내는 expolinear 생장 함수를 이용하여 각 함수의 매개변수를 추정하고 시계열에 따른 생장률을 분석하였다(Goudriaan and van Laar, 1994; Lee, 2002; Park and Lee, 2021). ‘백마’의 초장의 신장, 엽면적의 확장특성은 Gompertz 생장 모델의 매개변수 최대 생장량(ymax), 상대 생장률(k)과 최대 절대 생장률에 도달한 정식 후 일수(tinf), intercept(y0)와 함께 측정값을 98% 이상 유의적으로 설명하였다(Fig. 2 and Table 2). Gompertz 생장 모델은 시그모이드형 생장을 잘 표현할 수 있는 모델로서, 유한생장 식물의 초장 신장패턴(Cao et al., 2019; Park and Lee, 2021)과 강낭콩과 감자의 엽면적 확장량을 묘사했던 연구 결과와 유사하였다(Fleisher and Timlin, 2006; Singh et al., 2017).

Fig. 2.
Dynamics of the plant height (A and B), plant leaf area (C and D), and total fresh weight (E and F) as a function of the number of days after planting the standard chrysanthemums ‘Baekma’ (A, C, and E) and ‘Iwanohakusen’ (B, D, and F) from 27 Aug 2019 to 12 Dec 2019. Lines are the values estimated by the Gompertz growth model (A, B, C, and D) or the expolinear growth model (E and F) with four or three parameters respectively from Table 2. Vertical bars larger than the symbols indicate the standard error of the means of three replications with six samples (n = 18).
‘백마’와 ‘백선’의 초장 intercept(y0_ph)에서는 차이가 없었으나, 재배기간 동안 신장률은 품종 간 상이하였다. ‘백마’의 최대 신장률(MRph)은 단위 일자당 ‘백선’ 보다 11% 더 신장하였으나, 단위일수에 따른 단위 초장당 1cm 신장에 필요한 생장률(kph)은 8% 낮았다. Yph의 차이는 장일처리기간 동안 ‘백마’의 절대 신장률이 높았고, MRph에 도달하는 일수가 지연됨에 따라 yph 가 상이하였다(Figs. 2A, 2B, 3A, and 3B; Tables 1 and 2). 또한 ‘백마’의 평균 절간장이 길었으며, 이는 봄, 여름, 가을 작형 모두 채화 시 ‘백마’가 ‘백선’보다 초장이 더 길었던 결과와 일치하며, ‘백마’는 ‘백선’을 모본으로 하고 부본을 ‘신마’로 한 교배 육종된 품종이며, 초장의 신장은 ‘백선’과 유전적 특성 차이의 영향이 미쳤을 것으로 판단된다(Table 1; Karlsson and Heins, 1994; Shin et al., 2005).
스탠다드 절화 국화 ‘백마’의 시계열에 따른 엽면적 확장량에서 매개변수 intercept(y0_la)는 ‘백선’ 보다 15% 더 높게 추정되었다. 최대 엽면적 확장률에 도달하는 정식 후 일수(tla)는 ‘백선’ 보다 ‘백마’가 5.7일 지연되었고, 최대 엽면적 확장률(MRla)은 29% 더 높았다(Table 2). 두 품종의 절간 수는 차이가 없었으나 식물체 당 평균 개별 엽면적(cm2·leaf-1)은 ‘백마’에서 34% 더 넓었다. 스탠다드 절화 국화는 화성유도 및 화아분화 이후 출엽이 되지 않고, ‘백마’ 품종의 엽당 평균 엽면적이 넓어 ‘백선’보다 높은 최대 엽면적 확장량(yla)을 나타내었다(Figs. 2C, 2D, 3C, and 3D; Tables 1 and 2). Zandonadi et al.(2018)과 Kim et al.(2020)은 국화의 개별 엽면적의 차이는 품종 특성인 엽의 형태, 출엽된 엽수에 따라 다르다는 결과와 유사하였다.

Fig. 3.
Dynamics of the stem elongation rate (A and B), leaf expansion rate (C and D), and plant growth rate (E and F) as a function of the number of days after planting the standard chrysanthemums ‘Baekma’ (A, C, and E) and ‘Iwanohakusen’ (B, D, and F) from 27 Aug 2019 to 12 Dec 2019. Values were calculated by the Gompertz or expolinear growth model derivatives (Eqn. 2 or 4).
절화 국화의 생체중은 외형적 품질 지표 등급으로 경제적 측면에서 재배 환경과의 관계를 표현하는 중요한 요인이다(Kim et al., 2012). 정식 후 절화 총생체중 증가량은 ‘백마’와 ‘백선’에서 정식 후 생육 초기에 지수적 생장을 보인 후 선형적으로 증가하였다(Fig. 2E and 2F). 시계열에 따른 총생체중의 생장량 변화를 지수선형함수의 파라미터 최대 절대 생장률(cm; g·plant-1·d-1)과 최대 상대 생장률(rm; d-1) 그리고 지수생장에서 선형 생장으로 변화되는 정식 후 일수(tb; d)의 세 개의 파라미터로 측정값을 98% 이상 유의적으로 설명할 수 있었다(Table 2).
총생체중의 최대 절대 생장률(cm)은 ‘백마’가 ‘백선’보다 47% 높았고, 지수생장에서 선형 생장으로 변화되는 일 수(tb)는 두 품종이 유사하였다(Fig. 3E and 3F; Table 2). Lee et al.(2003)은 절화 국화의 생장량을 지수선형함수를 이용하여 광량에 따른 최대 절대 생장률(cm)과 최대 상대 생장률(rm)을 예측할 수 있었다. 본 실험에서도 지수선형함수를 이용하여 시계열에 따른 품종 별 총생체중의 생장량을 예측 및 요약할 수 있었고, 국화의 재배환경 정보와 장일처리기간과 정식 시기에 따른 생장량의 분석을 통해 생장 파라미터들을 일반화하여 국화의 생장을 예측할 수 있을 것으로 판단된다.
Table 2.
The growth parameters of the Gompertz or expolinear growth model were estimated with a measurement dataset of the plant height, plant leaf area, and total fresh weight for the standard chrysanthemums ‘Baekma’ and ‘Iwanohakusen’ from 27 Aug 2019 to 12 Dec 2019. The parameters yph and yla indicate the maximum plant height and maximum plant leaf area, respectively. In addition, kph and kla (d-1) denote the relative stem elongation rate and the relative leaf area expansion rate; tph and tla (d) represent the number of days to the reach maximum growth rate (MRph or MRla) for the maximum stem elongation rate and the maximum leaf area expansion rate, respectively. y0_ph and y0_la are the corresponding values of y when x is minus infinity. MR is calculated by the derivative of Eqn. 2 at tinf; where cm denotes the maximum growth rate, rm is the maximum relative growth rate, and tb denotes the lost time when reaching the linear phase
|
Plant height | Cultivar | yph (cm) | kph (d-1) | tph (d) | y0_ph (cm) | R2 |
MRphz (cm·d-1) |
| ‘Baekma’ | 95.1 ± 6.6 | 0.048 ± 0.005 | 46.0 ± 1.4 | 7.0 ± 3.6 | 0.997 | 1.68 | |
| ‘Iwanohakusen’ | 79.4 ± 6.4 | 0.052 ± 0.007 | 42.0 ± 1.4 | 7.0 ± 3.4 | 0.996 | 1.52 | |
|
Plant leaf area | Cultivar | yla (cm2·plant-1) | kla (d-1) | tla (d) | y0_la (cm2·plant-1) | R2 |
MRlay (cm2·d-1) |
| ‘Baekma’ | 765.6 ± 35.5 | 0.055 ± 0.005 | 47.1 ± 1.2 | 43.1 ± 21.9 | 0.995 | 15.49 | |
| ‘Iwanohakusen’ | 583.9 ± 79.8 | 0.056 ± 0.012 | 45.0 ± 2.2 | 37.4 ± 23.6 | 0.983 | 12.03 | |
|
Total fresh weight | Cultivar | cm (g·plant-1·d-1) | rm (d-1) | tb (d) | - | R2 | - |
| ‘Baekma’ | 0.871 ± 0.032 | 0.252 ± 0.750 | 21.3 ± 1.4 | - | 0.984 | - | |
| ‘Iwanohakusen’ | 0.591 ± 0.039 | 0.226 ± 0.496 | 20.7 ± 2.0 | - | 0.983 | - |
초장의 생장 패턴과 유사하게 두 품종의 절간 수의 변화는 sigmoid의 형태를 나타냈다(Fig. 4A and 4B; Karlsson and Heins, 1994). 절화 국화의 초장은 평균 절간 수와 평균 절간장에 의해 결정되고(Carvalho et al., 2002), ‘백마’와 ‘백선’의 최대 절간 출현율은(MRi) 차이를 보이지 않았다. 스탠다드 절화 국화는 단일처리 후 마디의 출현이 이루어지지 않으며, ‘백마’는 단일처리 이후 출뢰가 지연됨에 따라 ‘백선’은 하국 및 하추국으로 절간 출현율이 하추국인 ‘백마’ 보다 상대 절간 출현율(ki)이 6% 높았으나, 최대 절간 출현율에 도달하는 정식 후 일수(ti)에서 ‘백마’가 ‘백선’ 보다 4.2일 지연되었고, 품종 간 최대 절간 수(yi)는 큰 차이를 나타내지 않았다(Fig. 5A and 5B; Table 3).

Fig. 4.
Dynamics of the number of internodes as a function of days after planting for the standard chrysanthemums ‘Baekma’ (A) and ‘Iwanohakusen’ (B) from 27 Aug 2019 to 12 Dec 2019. Lines represent values estimated by the Gompertz growth model with the four parameters in Table 3. Vertical bars larger than the symbols indicate the standard error of the means of three replications with six samples (n = 18).

Table 3.
The growth parameters of the Gompertz growth model were estimated with a measurement dataset of the number of internodes on the standard chrysanthemums ‘Baekma’ and ‘Iwanohakusen’ from 27 Aug 2019 to 12 Dec 2019. The parameter yi indicates the maximum number of internodes, ki (d-1) denotes the relative internode appearance rate, and ti (d) represents the number of days to reach the maximum growth rate (MRi) for the internode appearance rate. y0_i is the value of y when x is minus infinity. MRi is calculated from the derivative of Eqn. 2 at ti
| Cultivar | yi (nodes·plant-1) | ki (d-1) | ti (d) | y0_i (nodes·plant-1) | R2 |
MRiz (nodes·d-1) |
| ‘Baekma’ | 43.8 ± 7.5 | 0.049 ± 0.059 | 47.5 ± 3.4 | 6.9 ± 2.6 | 0.985 | 0.79 |
| ‘Iwanohakusen’ | 40.6 ± 3.2 | 0.052 ± 0.009 | 43.3 ± 1.7 | 7.3 ± 1.9 | 0.989 | 0.78 |
비선형 모델의 주요 장점은 간결함, 해석 가능성 및 예측이다(Bates and Watts, 2007). Archontoulis et al.(2014)에 의하면 비선형 모델에서 매개변수 수를 감소시키고 매개변수의 해석 가능한 생물학적 의미가 있다면 선형모델보다 더 간결하게 사용할 수 있다고 보고하였다. 본 연구에서도 비선형 모델을 이용하여 스탠다드 절화 국화 ‘백마’와 ‘백선’ 두 품종의 시계열에 따른 초장, 엽면적, 총 생체중, 절간 수의 생육량을 유의적으로 설명할 수 있었다.
‘백마’와 ‘백선’의 초장에 따른 총 생체중이 유의적인 선형적인 관계(R2 > 0.96; p < 0.001)로 나타났고, Kim et al.(2015)의 고추의 생체중과 초장의 선형적 관계와 유사하였다(Fig. 6; Kim et al., 2015). ‘백마’는 1cm의 단위 초장당 총생체중은 ‘백선’보다 32% 더 무거웠으며, 줄기 1cm당 줄기 생체중(SSW)은 46% 더 무거웠다. 동일한 환경조건에서도 스탠다드 절화 국화의 단위 초장에 따른 줄기 및 총생체중의 차이를 확인할 수 있었으며, 초장에 따른 국화의 절화중을 예측할 수 있었다(Fig. 6 and Table 1).
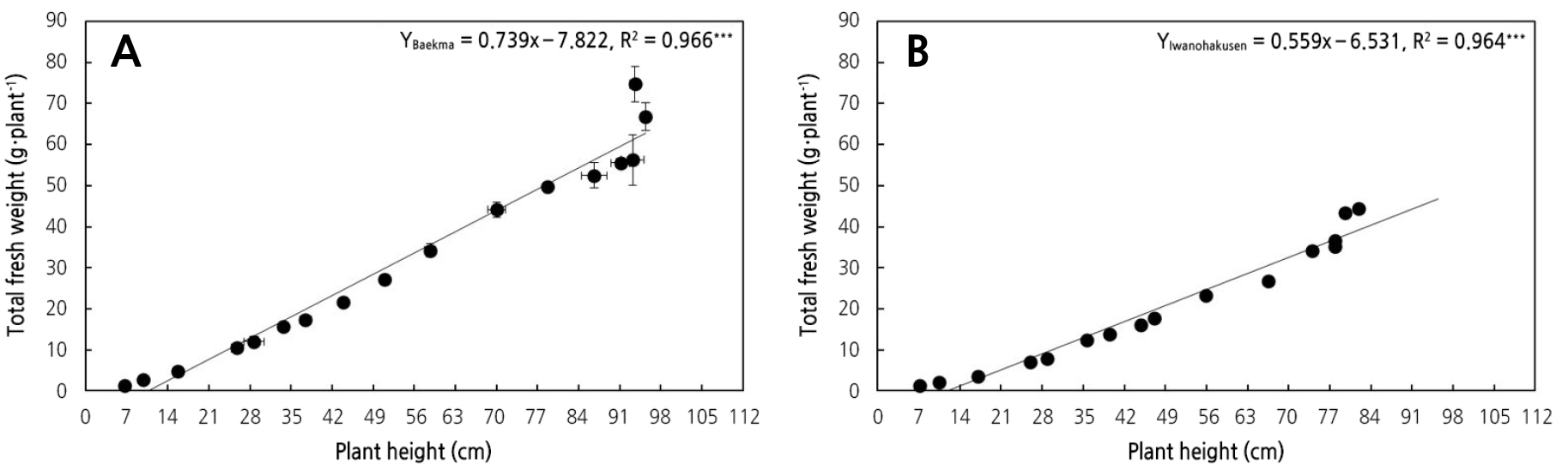
Fig. 6.
Relationship between the plant height (cm) and total fresh weight (g·plant-1) of the standard chrysanthemums ‘Baekma’ (A) and ‘Iwanohakusen’ (B) from 27 Aug 2019 to 12 Dec 2019. Vertical bars larger than the symbols indicate the standard errors (SE) of the mean of three replications with six samples (n = 18).
본 연구에서는 스탠다드 절화 국화 ‘백마’와 ‘백선’의 동적 생육자료를 빅데이터 분석 및 작물 생장 모델의 기초자료로 활용하고자 시계열에 따른 생육 특성을 분석하였다. 동일한 환경조건에서 재배된 두 품종의 최종 생육량은 상이한 결과를 나타냈다. 하지만 품종 간 단순 생육 비교가 아닌 묘사적 생장모델인 Gompertz 생장 모델을 이용하여 시계열에 따른 시그모이드 생육을 나타낸 초장, 엽면적과 절간 수의 측정값을 파라미터와 함께 98% 이상 요약 및 설명하였다. 측정된 총생체중은 최대 상대 생장률, 최대 절대 생장률과 지수생장에서 선형생장으로 전환되는 정식 후 일 수의 3개 매개변수와 함께 지수선형함수는 측정된 값의 98% 이상 설명 및 요약할 수 있었다. 정식 후 일수에 따른 국화의 동적 생육 반응을 이용하는 것은 재배과정에서의 생육조절에 대한 의사결정에 중요한 요소이다. 특히 스탠다드 절화 국화에 대한 초장, 생체중의 동적 생육에 대한 정보는 생산현장에서 정식 일 수와 품종에 따른 중요한 생육 지표로 이용될 수 있을 것으로 판단된다. 또한 정식 시기별 온도와 일장에 따른 Gompertz 생장함수와 지수선형함수의 파라미터 값 변화 정도를 알기 위해서는 추후 재배작형과 관련된 정식시기 및 장일처리 및 단일처리기간과 환경요인 중 광과 온도환경에 따른 파라미터의 변화에 대한 연구가 필요할 것으로 판단된다.
