서 언
재료 및 방법
식물재료 및 재배조건
시설 내 가온처리
광합성 특성 조사
식물계절 및 생육특성 조사
과실특성 조사
통계분석
결과 및 고찰
시설 내 일평균 온도
식물계절학적 특성
광합성 특성
신초의 생육특성
개화 및 착과 특성
과실의 품질
서 언
감귤은 상록과수로 추위에 약하기 때문에 세계적으로 최저기온이 -7°C 이상인 지역에서 주로 재배되고 있으며(Davies and Albrigo, 1994), 우리나라에서는 기후가 온난한 제주지역과 남해안 일부에서 재배되고 있다. 제주도의 연평균 기온은 15.4-16.7°C로 만감류를 포함한 감귤의 재배지 중 추운 지역에 해당되지만(JRMO, 2012), 하우스 재배가 일반화되면서 부지화[不知火, Shiranuhi; (Citrus unshiu × C. sinensis) × C. reticulata], 청견(淸見, Kiyomi; C. unshiu × C. sinensis), 진지향[津之香, Tsunogaori; (C. unshiu × C. sinensis) × C. unshiu Marc.] 등 추위에 약한 품종들도 도입되어 재배되고 있다. 이 중 부지화(상품명: 한라봉)는 1990년대 초반부터 재배되기 시작하여 매년 재배면적과 생산량이 증가되고 있으며(CMSA, 2012), 2013년에는 전체 감귤생산면적의 약 5% 정도인 1,362ha에서 41,400톤이 생산되었다. 부지화의 재배면적과 생산량이 매년 증가하는 것은 다른 감귤류에 비하여 당산비가 높아 맛이 좋고, 오렌지 수입 및 다른 경쟁 과실의 공급 증가로 인해 경쟁력이 약화되는 제주지역 주요 과수인 온주밀감(C. unshiu)의 대체 품종으로서의 가치가 있기 때문인 것으로 보고되었다(BOK, 2003).
부지화는 일본 과수연구소에서 1972년 청견에 중야 3호 폰칸(C. reticulata)을 교배해서 육성되었으며 과실의 가용성고형물 함량은 보통 12.5-15.8°Brix로 다른 감귤 품종보다 높은 편이고 과실에는 종자가 거의 없으며 껍질을 벗기기가 용이할 뿐만 아니라 limonene 등의 강한 향기 성분을 가지고 있다(Matsumoto, 2001; Song et al., 2005). 그리고, 부지화는 온주밀감보다 내한성이 약하여 연평균 기온 16.5°C 이상, 최저기온 -3°C 이상인 지역에서 주로 재배되고 있으며, 재배지역의 온도나 수분 상태 등 환경에 따라 수체 생육 및 과실 품질의 차이가 크게 나타난다(Han and Kang, 2011; Moon et al., 2008; Song et al., 2005). 일본에서는 노지재배가 이루어지고 있는데 비해(Kawase, 1999), 우리나라에서는 대부분이 하우스에서 가온 재배하거나 비가림 시설을 이용하여 재배되고 있다(CMSA, 2012).
부지화는 하우스에서 재배하면 수세가 안정될 뿐 아니라 과실발달이 양호하여 수량이 증가하지만(Kawase, 1999), 하우스 내에서도 봄철 생육기의 온도 및 대목의 종류 등 여러 가지 요인에 의해 다른 만감류의 품종보다도 수량 변동의 폭이 크고 품질에 있어서도 차이가 크게 나타난다(Lee et al., 2006; Moon et al., 2010). 더군다나, 부지화 재배 시 봄철 발아 및 개화시기의 온도가 낮아 새순과 새잎의 신장이 불량하면, 뿌리의 생장이 순조롭지 못하고 과실발달이 제대로 이루어지지 않아 작은 과실이 증가하게 되고 산 함량의 감소도 지연되는 경향이 있다(Matsumoto et al., 1972).
따라서 본 연구에서는 봄철 생육기(발아 및 개화시기)의 가온처리가 부지화의 생장 및 과실품질에 미치는 영향을 규명하여 금후 고품질의 과실을 생산하기 위한 기초적인 자료를 얻기 위해 수행하였다.
재료 및 방법
식물재료 및 재배조건
본 실험의 식물재료는 탱자(Poncirus trifoliate Rafin.) 대목에 접목된 6년생 부지화[不知火, Shiranuhi, (Citrus unshiu × C. sinensis) × C. reticulata)]이며, 외관상 균일한 상태의 나무를 선별하여 사용하였다. 부지화의 육묘는 2년생 묘목을 탱자 대목에 접목시킨 후 제주도 서귀포시 남원읍에 소재한 감귤연구소의 하우스 내에 2.0 × 2.7m로 재식하여 4년간 균일하게 키웠으며, 육묘기간 동안의 하우스 내 환경은 무가온 상태에서 자연광의 80%가 투과되도록 하였다.
시설 내 가온처리
시설 내의 온도처리를 위해 2월 중순에 하우스 바깥쪽 천정 및 측면 외부에 직경 0.1mm 폴리에칠렌 필름을 사용하여 피복하고, 내부에는 0.05mm 폴리에칠렌 필름으로 2개의 층이 되도록 피복하였다. 가온처리는 2월 중순부터 5월 말까지 약 100일 동안 가온처리I(25/15°C, 주간/야간)과 가온처리II(28/18°C, 주간/야간)의 조건이 되도록 하였으며, 대조구는 가온하지 않았다. 그 이후부터 과실의 수확시까지는 하우스 내부 2층의 필름을 완전히 제거하고 바깥쪽 필름의 측면 및 천정 부분의 비닐을 개방하여 대조구와 가온처리구가 동일한 조건이 되도록 조성하고, 자연 온도에 가깝게 유지하였다. 가온처리는 실시간으로 수집된 기상정보를 바탕으로 복합환경제어시스템(TGC-Soldan, Soldan Corp., Korea)을 이용하여 조절하였다. 시험기간 동안의 온도는 지상부로부터 2.5m 높이에 설치한 온도센서(1400-101, LI-COR Inc., Lincoln, USA)를 이용하여 측정하고, 데이터로거(CR1000, Campbell Scientific Inc., Logan, USA)에 연결하여 1시간 간격으로 기록하였다. 하우스 내 수분 관리는 가온 전날에 50mm에 상당하는 양의 물을 시험수 전체에 고르게 관수를 하고, 이후에는 농촌진흥청에서 고시한 표준영농교본에 준하였다.
광합성 특성 조사
광합성 특성은 3월 중순의 오전 9시부터 11시 사이에 처리구별로 수세가 비슷한 나무를 5개체씩 선정하여 수관의 동서남북 4방향에서 나무의 중간 상부에 위치한 15cm 정도의 전년도 봄순의 끝에서 세 번째의 잎을 대상으로 조사하였다. 광합성은 휴대용 광합성측정기(Li-6400, Li-Cor, USA)를 사용하여 측정하였으며 챔버에 잎을 물리고 5분간 안정화시키고 1,000μmol・m-2・s-1의 포화광을 비추어 CO2 고정률(photosynthetic rate)을 측정하였다. 아울러 기공전도도(stomatal conductance), 증산율(transpiration rate) 등을 측정하고 증산율에 대한 CO2 고정률로 수분이용효율(water use efficiency)을 산출하였다.
식물계절 및 생육특성 조사
식물계절(phenological events) 및 생육특성은 눈의 발아와 신장, 개화, 개엽, 과실의 발달과 생장 등을 중심으로 조사하였다. 발아기는 육안으로 관찰하여 봄철에 발생하는 정아 및 액아가 3mm 이상 발생한 시기로 나타내었으며, 만개기는 꽃이 70-80% 이상 개화되었을 때를 기준으로 하였고, 수확기는 과피의 색이 주황색으로 변하고 당산비가 10 범위에 해당하는 시점을 전후로 하였다. 봄순의 길이와 어린 잎은 순이 녹화된 6월 초순에 각 처리구당 20개의 봄순을 무작위로 선정하여 조사하였다. 엽면적은 봄순의 끝으로부터 3번째 위치에 달린 잎 10매를 채취하여 엽면적계(LI-3100, Li-Cor, USA)로 조사하였다. 그리고 신초의 형성률은 개체당 신초의 수와 구엽의 수를 이용하여 나타내었다. 착화수는 만개 직전에 수관의 동서남북 4방향에서 착화수가 중간 정도이고 200매 내외의 구엽을 가진 측지를 나무당 4개씩 표지하고 꽃, 구엽, 신초의 수를 세어 구엽과 신초에 대한 꽃의 수를 제시하였다. 착과량은 생리적 낙과가 완료된 6월 중순에 착화수 조사시 표지해 둔 가지의 과실수를 세어 앞서 조사한 꽃과 신초의 수를 기준으로 나타내었다.
과실특성 조사
과실의 크기는 6월 하순부터 12월 상순까지 수관의 동서남북 4방향에서 나무의 중간 상부에 착과된 유엽과 과실을 각 처리구당 10과씩을 선정하고 1개월 간격으로 디지털캘리퍼스(WA 100, Mitutoyo, Japan)를 사용하여 과실의 종경과 횡경을 측정하였으며, 과형지수는 종경과 횡경의 비로 나타내었다. 그 밖의 과실특성은 과피의 색이 변하기 시작한 9월 하순부터 완전히 착색이 이루어져서 수확할 때까지 1개월 간격으로 나무의 중간 상부에 착과된 과실 9-10개를 채취하여 분석하였다. 과중과 과육중은 디지털 저울을 사용하여 측정하였으며, 과피의 두께는 과육이 손상되지 않게 껍질을 벗긴 후 디지털캘리퍼스로 측정하였다. 과피의 착색 정도는 색차계(Color-Eye 2145, Macbeth, USA)를 이용하여 과실 중간부위의 3지점에서 Hunter L, a, b값을 측정하고, 칼라지수(a/b)로 표시하였다. 그리고, 과피를 벗겨낸 과육은 여과지가 들어있는 착즙기로 착즙한 후 당산분석장치(NH-1000, Horiba, Japan)를 이용하여 가용성고형물과 산 함량을 측정하였고, 당산비는 가용성고형물과 산 함량의 비율로 나타내었다.
통계분석
통계분석은 SPSS 통계 패키지 Release 7.5(SPSS Inc., Chicago, USA)를 이용하여 수행하였으며, 각 처리간의 유의성은 단일변량 분산분석(ANOVA)으로, 평균간의 유의성은 Duncan 다중검정으로 분석하였다.
결과 및 고찰
시설 내 일평균 온도
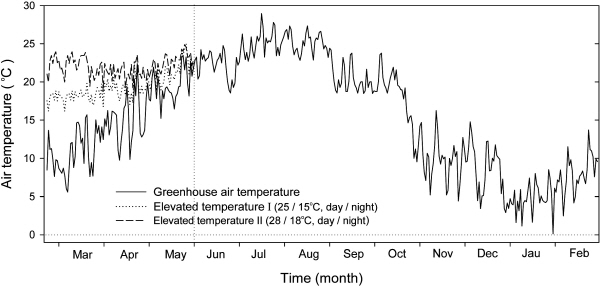
조사기간 동안 시설 내의 년간 일평균 온도 변화는 Fig. 1과 같다. 대조구(무가온 시설재배)에서는 이른 봄철(2-4월)에 일평균 온도가 15°C 이하를 나타내는 일수가 많아 비교적 차갑고, 5월부터 점차 상승하여 6월 이후부터 10월까지는 20°C 이상의 온도를 나타내었다. 그리고 10월 하순부터 일평균 기온이 15°C 이하로 점차 낮아지면서 1월에는 5°C 이하로 현저히 낮아졌다. 그러나, 봄철에 시설 내 주야간의 온도를 조절하여 가온하였을 때, 가온처리I(25/15°C, 주간/야간)에서는 이른 봄철(2-3월)에는 일평균 온도가 대략 18°C 전후였으나 4월부터 외부 기온이 높아짐에 따라 서서히 높아져 20°C 이상의 온도를 나타내었다. 그리고 가온처리II (28/18°C, 주간/야간)에서는 일평균 온도가 2월부터 5월까지 22°C를 전후하여 큰 변화를 나타내지 않았다. 6월부터는 가온처리한 하우스의 내부 2층의 필름을 완전히 제거하고 바깥쪽 필름의 측면 및 천정 부분의 비닐을 개방하였으므로, 가온처리I과 가온처리II의 온도는 대조구의 온도와 같았다(Fig. 1).
식물계절학적 특성
식물계절학적 특성에 미치는 봄철 가온처리의 영향을 살펴보면, 가온처리I과 가온처리II에서 이른 봄철 눈의 발아는 대조구에 비해 각각 11일, 15일 정도, 만개기는 각각 22일, 45일 빨라져, 온도가 높을수록 눈의 발아와 개화 소요일수가 줄어들었다(Table 1). 이와 유사하게 온주밀감에서도 온도가 높을수록 개화가 앞당겨진다는 사실이 보고되었다(Inoue, 1990b; Moon and Kim, 2001). 식물의 개화나 개엽은 온도, 토양수분, 일장 등 여러 환경요소에 의해 결정되며, 특히, 봄철 생육기의 기온이 가장 지배적으로 작용한다(Brown, 1953; Lindsey and Netman, 1956). 온주밀감에서도 겨울철 저온 후 기온이 올라가게 되면 휴면이 타파되어 눈이 발아하고 개화하게 되는데(Reuther et al., 1973), 봄철에 적절한 가온은 눈의 발아와 개화에 필요한 소요일수를 줄이고 과실의 수확시기를 앞당기게 된다(Yamanish, 1994). 더군다나 개화시기는 봄철의 상해, 수분 및 결실에 의한 생산성의 변화와 관련이 깊어, 과실의 안정적인 생산 및 출하시기 조절 등 경제적인 측면에서 중요할 뿐만 아니라, 변화하는 기상 또는 기후에 대한 대처 측면에서도 중요하다고 할 수 있다. 그리고, 봄철의 가온처리가 과실의 발달과 생장에도 우호적으로 작용하여, 가온처리I의 조건으로 재배하였을 때에는 12월 하순부터, 그리고 가온처리II의 조건에서는 11월 하순부터 수확이 가능하여, 무가온 시설재배인 대조구에 비하여 과실의 수확시기를 대략 1-2개월 앞당길 수 있었다(Table 1).
광합성 특성
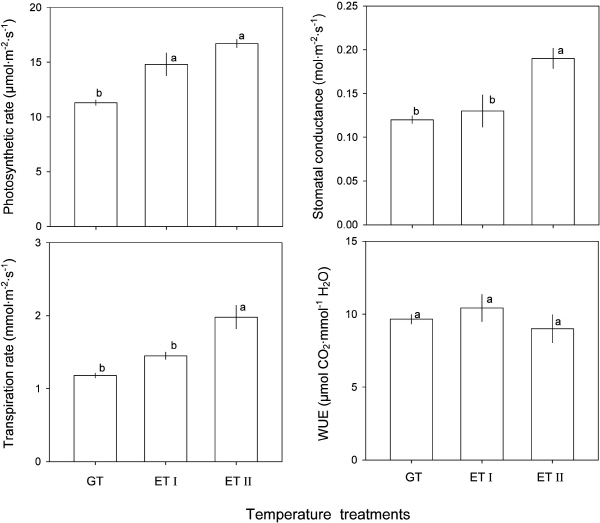
감귤속 식물들의 생육을 위한 최적온도는 22-30°C이고(Sun and Ma, 1999), 대부분의 감귤속 식물들은 덥고 습한 적도에서부터 따뜻한 아열대 지역에 걸쳐 다양한 기후조건에서 재배가 가능한 것으로 알려져 있다(Guo et al., 2000). 부지화의 CO2 고정률 역시 대조구에서 보다 가온조건에서 높은 것으로 나타났다(Fig. 2A). 즉, 가온처리I과 가온처리II 조건에서는 부지화 잎의 광합성 속도가 각각 14.8μmol CO2・m-2・s-1와 16.7μmol CO2・m-2・s-1로 대조구에서 자란 부지화 잎의 광합성 속도 11.3μmol CO2・m-2・s-1와 비교하였을 때 훨씬 높았다. 이와 같은 결과로부터 제주지역의 이른 봄철의 무가온 시설 내 온도가 Fig. 1에서 보는 바와 같이 비교적 낮아서, 봄철 부지화의 발달과 생장에 있어 가온이 우호적으로 작용하고 있음을 알 수 있다(Ge et al., 2012). 이와 유사하게 청도온주(C. unshiu cv. Aosima)와 흥진조생(C. unshiu cv. Okitsu Wase)에서는 25°C 전후, 태전폰칸(C. reticulate cv. Ota ponkan)과 천야하귤(C. natsudaidai cv. Kawano natsu)에서는 30°C 전후의 온도에서 광합성이 가장 왕성하게 일어나는 것으로 보고되었다(Iwasaki and Oogaki, 1985). 그리고 온주밀감(C. unshiu)에서도 광합성이 왕성하게 일어나는 시기인 7-9월의 온도는 대략 29°C, 10월의 온도는 22°C로 높다(Ono and Daito, 1982). 그러나, 지속적인 가온은 식물의 후기 생장 단계에 있어서 엽록소 함량 및 광합성 능력의 감소로 이어지고, 급기야는 식물의 노화를 야기할 수 있기 때문에(Ge et al., 2011; Zhou et al., 2011), 부지화 재배에 있어 적정한 온도의 규명과 더불어 가온 시기 및 기간을 좀더 체계적으로 살펴볼 필요가 있다.
한편, 봄철 부지화 잎의 기공전도도와 증산율은 대조구나 가온처리I에서 보다 가온처리II 조건에서 크게 높았다(Fig. 2B, 2C). 이는 종에 따라 일정 온도가 될 때까지는 온도가 증가함에 따라 광합성은 물론 기공전도도와 증산율이 증가한다는 사실로 미루어 보았을 때(Oh et al., 2015), 봄철 가온처리II 조건에서의 재배는 부지화의 생육을 위해 유용한 것으로 보인다. 이와 유사하게 발렌시아오렌지(C. sinensis cv. Valencia)를 저온(15.6/4.4°C, 주간/야간)과 고온(32.2/21.1°C, 주간/야간) 조건에서 30일간 재배하였을 때에도 CO2 고정률뿐만 아니라, 기공전도도와 증산율이 고온에서 증가하는 것으로 보고된 바 있다(Vu and Yelenosky, 1987). 하지만, 수분이용효율은 통계적으로 유의한 차이를 보이지는 않았지만 대조구나 가온처리I에서 보다 가온처리II 조건에서 다소 낮았다(Fig. 2D). 이러한 결과는 봄철 가온처리II 조건이 부지화의 광합성을 촉진하지만 수분수지에는 크게 영향을 미치지 않는 조건임을 나타내 주고 있다.
신초의 생육특성
봄철 발아기 및 개화기에 온도를 달리하여 부지화를 재배하였을 때 봄순의 발생빈도(구엽 기준)는 온도가 높을수록 감소하는데 반하여, 봄순의 평균 길이는 대조구에서 14.3cm인데 반해 가온처리I과 가온처리II에서는 각각 25.9cm와 27.7cm로 온도가 높아짐에 따라 유의하게 길었다(Table 2). 그리고 신초의 잎 수는 대조구에서 다소 많았으며, 엽면적은 대조구에 비해 가온처리구에서 더 넓었다. 부지화 재배시 문제점 중의 하나가 봄순이 액아 마다 발생할 뿐 아니라 1개의 액아에서 2-3개의 새순이 발생하여 그대로 두면 가지 길이도 짧고 잎이 아주 작은 빈약한 가지가 되어 수세가 쇠약해지므로(Matsumoto, 2001) 농가들이 순을 솎는데 많은 시간과 노동력이 소모된다는 점이다. 본 연구에서도 대조구에서 재배하였을 때에는 주당 발생하는 봄순의 수가 많고 길이는 짧아졌으나 가온하였을 때에는 봄순의 수가 적어지고 길이가 길어지는 양상을 보여 부지화 재배시 봄철의 발아기 및 개화시기에 적절한 가온이 봄순 생장에 유리하게 작용하고 있음을 알 수 있었다. 부지화의 엽면적은 품종 육성시 15.3cm2 정도로 보고되었는데(Matsumoto, 2001), 본 연구에서 봄철 일반 시설에서 재배된 부지화의 잎은 이보다 좁은 반면 가온하여 재배된 잎은 더 넓어진 것으로 나타났다. 이러한 결과는 부지화 재배 시험에서 온도가 낮은 노지보다 무가온 하우스에서 봄순의 길이가 길고 엽면적도 넓어졌다는 보고와도 유사하다(Kitazono, 2001). 일반적으로 엽면적이 증가하면 과실생장이 촉진되는데(Fishler et al., 1983), 부지화에서는 가온으로 엽면적을 충분히 확보하여 과실품질을 향상시킬 수 있을 것으로 보인다.
개화 및 착과 특성
착화수는 온도가 높을수록 점차 감소하였는데, 구엽당 착화수의 차이는 통계적으로 유의하였으나 봄순당 착화수의 차이는 유의성이 인정되지 않았다(Table 3). 가을과 겨울철의 저온으로 화아분화가 잘 되면 착화수가 많아진다(Inoue and Harada, 1988; Inoue, 1990a, 1990b; Okada, 1985). 그러나 저온 이후에 봄철의 온도가 높아지면 개화시기가 앞당겨지고 착화수가 감소하게 된다(Inoue, 1990a; Poerwanto and Inoue, 1990; Yamanishi, 1994). 온주밀감에서 봄철에 온도가 높을수록 착화수가 감소하였고(Inoue, 1990a; Poerwanto and Inoue, 1990), 포멜로(C. grandis (L.) Osbeck)에서는 봄철 가온에 의해 착화수가 감소하는데 반하여 슈트와 잎의 발생이 촉진되는 것으로 밝혀졌다(Yamanishi, 1994). 따라서 본 연구에서 온도가 높을수록 착화수가 감소하는 것은 고온에 의해 영양생장이 더 활발하게 일어나기 때문인 것으로 보인다. 착과율은 꽃 수를 기준으로 표시했을 때에는 가온처리의 영향을 받지 않아 꽃이 열매로의 발달은 온도의 영향에 둔감함을 나타내주었다. 반면에 봄순을 기준으로 표시했을 때에는 꽃의 열매로의 발달이 가온처리구에서 유의하게 낮았다. 이는 봄철 가온조건에서 재배하였을 때에는 대조구에서 재배하였을 때보다 봄순의 감소가 착화수의 감소 보다 크기 때문이다. 그러나, 가온처리시 착과율이 낮은 것은 과실생장에 긍정적으로 작용할 가능성이 있다. 부지화는 온주밀감과는 달리 과실이 클수록 상품성이 높기 때문이다. 따라서, 부지화 재배시 봄철에 적절한 가온처리는 과실의 크기를 조절하는데 유용할 것이다. 온주밀감이나 네블오렌지의 경우 착화량이나 착과량이 너무 많아 착과부담이 커지면 과실의 크기는 작아지는 것으로 보고되었다(Morioka, 1988; Stover, 2000).
과실의 품질
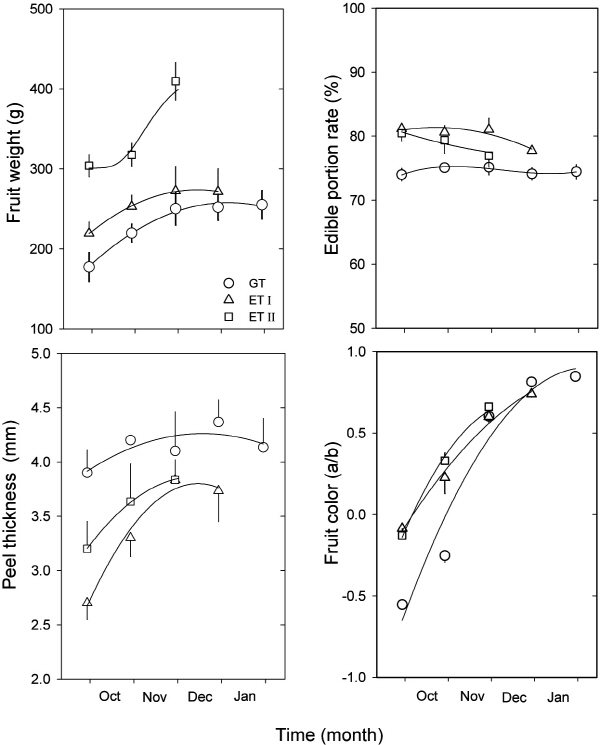
부지화의 과실 종경은 조사 개시일인 6월 하순부터 과실 비대가 거의 멈춘 12월 초순까지 대조구에 비해 봄철 생육기(발아 및 개화시기)에 온도를 높여서 재배하였을 때에 훨씬 크게 발달하였다(Fig. 3A, Table 4). 과실 횡경도 종경과 마찬가지로 측정시기에 관계없이 가온처리구에서 크게 생장이 이루어졌다(Fig. 3B, Table 4). 조생 온주밀감에서도 고온일수록 종경 비대가 왕성하다고 하였고(Nii et al., 1970), 개화기의 고온 조건에 의해 과경부의 생장이 왕성하여 기형과가 발생하였다고 한다(Takagi et al., 1982). 그러나, 부지화에서 과실의 종경 비대는 꼭지깃의 발달을 의미하기 때문에 품종 고유의 과실특성이 나타나는 바람직한 결과라고 할 수 있다. 과형지수는 가온조건에서는 측정시기에 관계없이 1.0 이상으로 온도가 높을수록 횡경보다는 종경이 커서 과실이 도란형에 가까운 모양을 나타내었다(Fig. 3C, Table 4). 일반적으로 부지화는 꼭지깃이 발달한 도란형의 과실이 상품성이 높으므로 품종 특성을 발현시키기 위해서는 개화시기나 과실비대 초기에 온도를 높여주는 것이 바람직한 것으로 보인다.
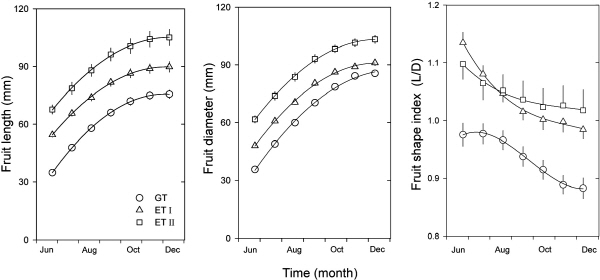
과중은 과실이 발달하는 동안 대조구나 가온처리I에서 보다 가온처리II에서 훨씬 더 높았으며(Fig. 4A), 수확 후 온도처리간의 비교시에는 가온처리II에서 가장 높았다(Table 4). 상품과로서 부지화의 과중은 200-300g이 적당한데(JCGAC, 2000), 봄철 가온처리I에서 재배하였을 때에는 이 범위를 유지하였으나, 가온처리II에서는 300g 이상으로 나타났다. 부지화는 과중이 클수록 국내 시장에서 좋은 평가를 받지만 너무 크면 당 함량이 낮아져 상품성이 저하될 수 있다. 본 연구에서도 가온처리II에서 과중은 높지만 가용성고형물 함량은 낮았다. 과육의 비율은 과실이 발달하는 동안에는 가온처리구에서 다소 높았으나(Fig. 4B), 수확 후 온도처리간 비교시에는 통계적 유의성이 관찰되지 않았다(Table 4). 과피는 과실 발달과정이나 수확 후 모두 대조구에서 보다 가온조건에서 다소 얇아지나 통계적으로 유의한 차이가 없었다(Fig. 4C, Table 4). 과피의 착색은 11월 하순까지 대조구보다 가온조건에서 빨리 이루어져 칼라지수(a/b)가 높았으나, 수확 후 비교시에는 대조구에서 오히려 가장 높았다(Fig. 4D, Table 4).
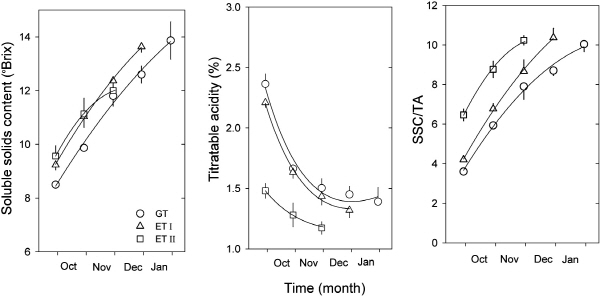
과즙의 가용성고형물 함량(SSC)은 과실이 성숙하는 동안 대조구보다 가온조건에서 높았다(Fig. 5A). 그러나 수확 후의 당도는 가온처리II보다 대조구와 가온처리I에서 더 높았으며 13°Brix 이상을 나타내었다(Table 5). 산 함량(TA)은 과실의 발달함에 따라 서서히 감소하였으며, 가온처리II 조건에서는 그 값이 대조구와 가온처리I에서 보다 크게 낮았다(Fig. 5B). 그러나, 수확 후에는 처리구 간에 통계적으로 유의한 차이는 없었다(Table 5). 당산비(SSC/TA)는 초기에는 가온처리II 조건에서 더 높았으나 수확 후에는 유의한 차이를 보이지 않았다(Fig. 5C, Table 5). 이러한 결과는 문단[C. grandis (L.) Osb.]에서 지나친 고온이 오히려 당과 산 함량을 모두 떨어뜨린다는 보고와 유사하다(Susanto and Nakajima, 1990). 부지화의 가온재배에 있어서 낮은 가용성고형물 함량이 문제가 되는 것을 감안했을 때(Song et al., 2005), 부지화 재배시 당도를 높이기 위해서는 대조구 또는 가온처리I 조건에서 재배하는 것이 더 적합한 것으로 판단된다. 최근 노지 및 시설에서 재배된 한라봉의 관능평가 결과를 보면, 시설에서 재배된 한라봉이 신맛이 높은 데 반해, 노지에서 재배된 한라봉은 당도가 높고 상대적으로 강한 향기특성을 지니고 있어 향과 맛을 고려한 전체적 선호도에서 노지 한라봉이 높게 평가된 바 있다(Song et al., 2005). 그러나, 과수재배의 주 목적이 상품성이 높은 고품질의 과실을 균일하게 생산하는 것과 더불어 조기 수확 및 출하시기 조절 등을 통해 농가소득에 도움이 되어야 한다. 더군다나 부지화는 내한성이 약하기 때문에 제주도의 기후조건에서 연중 지속적으로 품질이 좋은 과실을 생산하기 위해서는 가온이나 보온이 불가피하다. 이러한 측면에서 가온처리I의 조건으로 재배하였을 때에는 부지화의 조기 수확 및 출하가 가능하며, 과중 또한 무가온 재배에서와 차이가 없을 뿐 아니라 과형지수가 1.0 이상이고 가용성고형물 함량이 13.6°Brix로 높아 봄철 생육기에 가온처리I 조건 즉, 25/15°C(주간/야간) 조건으로 유지하는 것이 과실품질 향상을 위해서 바람직한 것으로 판단된다.