

서 언
개불알풀속(Veronica L.)은 현삼과(Scrophulariaceae)에 속하는 분류군으로(APG IV, 2016), 세계적으로 500여 종이 대다수 북반구와 일부 남반구에 거쳐 분포하며 수생지역에서 고산지대까지 서식지역이 다양하다(Albach et al., 2005). 우리나라에 자생하는 Veronica속 식물은 봉래꼬리풀[Veronica kiusiana var. diamantiaca (Nakai) T.Yamaz.], 부산꼬리풀(Veronica pusanensis Y.N.Lee) 등 23종이 보고되어 있다(KPNI, 2018). 특히 봉래꼬리풀, 섬꼬리풀, 부산꼬리풀은 우리나라에서만 자생하는 특산식물(endemic plant)이며(Chung et al., 2017), 봉래꼬리풀은 산림청 지정 멸종위기종[critically endangered (CR) species]으로 지정되어 있다(KNA, 2012). 부산꼬리풀은 우리나라에서도 부산지역에서만 자생하는 분포가 한정적인 희귀식물로 현재 산림청 지정 자료부족종[data deficient (DD) species]으로 분류되어(Son, 2013) 종 보존과 활용을 위한 기초자료 연구가 시급한 실정이다.
종자의 휴면은 식물이 불리한 환경조건에서 생존하기 위해 발아를 억제하는 수단으로 식물의 생활환, 서식지의 환경 및 지리적 분포에 따라 같은 속 사이의 식물간에도 휴면의 깊이가 다르게 나타날 수 있다(Karlsson and Milberg, 2008). 종자휴면의 종류는 생리적휴면(physiological dormancy, PD), 형태적휴면(morphological dormancy, MD), 형태생리적휴면(morphophysiological dormancy, MPD), 물리적휴면(physical dormancy, PY), 그리고 조합휴면(combinational dormancy, PY+PD)으로 크게 다섯가지로 나뉘어져 있다(Baskin and Baskin, 1998; Baskin and Baskin, 2004). 이러한 종자휴면은 특히 온대 북반구에 자생하는 식물들이 가지는 전형적인 특징인데, 같은 속(genus)이라고 하더라도 개체군 수준에서 휴면의 깊이가 다르거나 전혀 다른 휴면유형을 보이는 경우도 다수 보고되고 있다(Baskin and Baskin, 2014).
Corylopsis속 식물 중 C. sinensis var. calvescens의 종자는 20°C 또는 25°C 온도에서 세 달간 층적처리를 받은 뒤 5°C에서 세 달간의 저온처리를 하면 28%이상 발아하였다. 그러나 C. coreana의 종자는 10, 15, 20, 25°C 온도범위에서 한 달간 층적처리를 받은 뒤 5°C 온도에서 두 달간의 층적처리 시 90% 이상의 발아율을 보였다고 하여, 종간 차이를 보고한 바 있다(Roh et al., 2008). Heloniopsis속 식물 중 처녀치마(H. koreana)와 숙근처녀치마(H. tubiflora)의 종자는 각각 22%와 40%의 형태적휴면(MD)과 78%와 60%의 형태생리적휴면(MPD)이 있다고 보고되었는데(Lee et al., 2014), 일본 자생종인 H. orientalis의 종자는 30일 이내에 대부분 발아하여 형태적휴면(MD)을 보인다고 보고된 바 있다(Takahashi, 1984; Baskin and Baskin, 2014). 뿐만 아니라, Hosta plantaginea는 형태생리적휴면(MPD)을, H. minor는 형태적휴면(MD)를 갖는다고 하여, 같은 속(genus) 내에서도 종간에 전혀 다른 휴면유형을 보이는 분류군이 다수 보고되고 있다. 형태적휴면(MD)과 형태생리적휴면(MPD)은 종자가 모식물체에서 탈리되는 시점에 미숙배를 가지고 있기 때문에 나타나는 생태생리적 특성인데(Adams et al., 2005a, 2005b), 미숙배를 판단하는 기준은 단순히 배의 크기가 아니라 초기에 비하여 발아직전(또는 종피가 갈라지는 시점)까지 얼마만큼의 추가적인 배의 길이신장이 일어나는지가 가장 중요한 지표이다(Baskin and Baskin, 2005; Chen et al., 2015).
자생 Veronica속의 경우 종자의 내부 형태와 휴면유형, 휴면타파를 위한 구체적인 연구결과가 보고된 바 없다. Whittington et al.(1988)의 문헌에 따르면 미국 Alabama주에 자생하는 V. spicata L. 종자의 내부형태는 배유가 유근을 감싸고 있는 형태로서, 이로 인해 발아가 지연될 수 있다고 보고하였다. Guerin et al.(2013)은 South Australia주에 자생하는 V. parnkalliana 종자가 미숙배이며 형태생리적휴면(MPD)이라고 분류하였지만 발아하기 직전까지 종자 내부의 배가 신장하는지 ratio of embryo length to seed length(E:S ratio)를 제시하지 않았다. Mennan and Zandstra(2006)은 터키, Samsun주에 자생하는 V. hederifolia 종자가 모체에서 수확 직후인 9 - 10월에 깊은 휴면을 가지고 있다고 밝히고, 야외에 종자가 층적되어지는 기간 동안 종자의 휴면이 감소된다고 보고하였다. V. arvensis 종자는 10 - 15°C 온도 조건에서 58 - 99% 발아하지만 25°C에서는 전혀 발아하지 않았다고 보고된 반면(Baskin and Baskin, 1983), V. hederifolia L. 종자는 10 - 30°C 범위에서 발아가 촉진되고 저온(4°C)에서는 오히려 생리적 휴면이 없던 종자에서 2차 휴면이 유도되었다고 보고한 바 있다(Roberts and Neilson, 1982).
따라서 본 연구는 한국 희귀·특산식물인 봉래꼬리풀을 포함한 Veronica속 8종 종자의 발아특성을 조사하고 휴면 유형을 분류하여, 종 보존 및 효율적인 활용을 위한 생태생리적인 자료를 제공하고자 하였다.
재료 및 방법
실험 재료
본 연구는 2017년 9월 15일부터 2018년 7월 6일까지 국립수목원 유용식물증식센터에서 실시되었다. 연구에 사용된 공시재료는 Veronica속 8종으로 구와꼬리풀, 둥근산꼬리풀, 봉래꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰구와꼬리풀, 큰산꼬리풀이었다(Table 1). 구와꼬리풀 종자는 경상북도 의성군 춘산면 방계리의 자생지로부터, 섬꼬리풀 종자는 자생지인 울릉도 남양계곡에서 채종하였다. 부산꼬리풀과 큰구와꼬리풀 종자는 경기도 포천시 소재의 국립수목원에서 관리중인 개체에서 채종하였다. 둥근산꼬리풀과 큰산꼬리풀은 경기도 양평군 소재의 국립수목원 유용식물증식센터에서 관리중인 개체에서 채종하였다. 봉래꼬리풀과 산꼬리풀은 강원도 양구군 소재의 국립수목원 DMZ 자생식물원에서 관리중인 개체에서 채종하였다. 채종한 종자는 통풍이 잘 되는 곳에서 상온 건조 후 정선하여 실험에 사용하기 전까지 냉장(4°C) 보관하였다.
Table 1. Common name, scientific name, source, and date of seed gathering of eight Veronica species used in this study 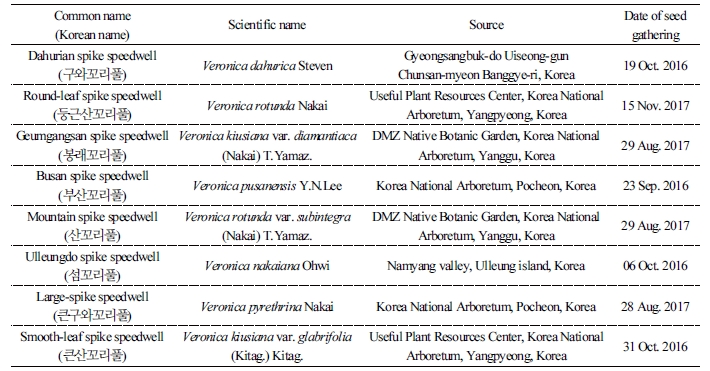 |
종자의 내·외부 형태 관찰
종자의 내·외부 형태를 관찰하기 위해 USB전자현미경(AM 3111 Dino-Lite premier, AnMo Electronics Co., Taiwan)을 이용하여 종자와 배의 길이를 측정하였다. 또한 종별로 20개 종자를 자른 후 미숙배인지 여부를 판단하기 위해서 초기의 배 길이, 종피가 갈라지는 시점에 배의 길이를 측정하였다. 같은 종이더라도 종자의 크기가 다양하고 배의 길이도 차이가 있어서, 미숙배를 좀 더 명확하게 표현할 수 있는 배(embryo)와 종자(seed)의 비율인 E:S ratio를 계산하였다(Vandelook et al., 2009; Chen et al., 2013).
발아 온도
종자는 발아상에 치상하기 전 500mg·L-1의 소독제(Benomyl, FarmHannong, Seoul, Korea)에 1시간 침지 소독한 뒤 증류수로 5회 이상 수세한 후 치상하였다. 모든 처리구는 90×15mm petridish에 여과지 2매를 깔고 그 위에 종자를 20립씩 파종한 뒤 처리별 완전임의배치로 3반복으로 치상하였다. 온도에 따른 발아반응을 알아보기 위해 소독된 종자를 4, 15, 20, 25, 30°C 항온으로 조절된 생장상(WCC-1000, Daihan Scientific Co., Wonju, Korea)에 배양하였다. 광도는 약 10µmol·m-2·s-1 조건에서 16시간 명조건을 유지하였으며, 유근이 1mm 이상 출현한 것을 발아된 것으로 간주하였다. 조사는 치상 후 24시간마다 발아 개체수를 조사하여 발아율(germination percentage, GP), 평균발아일수(mean germination time, MGT)를 아래의 식을 이용하여 계산하였다(Ellis and Roberts, 1981). 조사 기간 중 발아된 종자들은 제거하였고, 건조되지 않도록 증류수를 보충해주었다. 조사 시 부패한 종자들은 발아율 계산에서 제외하였다.
• GP = (N / S) × 100
• MGT = ∑ (ti × ni)/ ∑ ni (N; 총 발아수, S; 총 공시 종자수, ti; 치상 후 조사일수, ni; 조사 당일의 발아수)
저온 층적처리
휴면타파 효과를 알아보기 위하여 봉래꼬리풀 종자를 대상으로 저온 층적처리를 실시하였다. 소독한 종자는 90×15mm petridish에 여과지 2매를 깔고 여과지가 마르지 않을 정도의 증류수를 주입한 후 치상하였다. Petridish 내 수분의 손실을 막기 위해 parafilm으로 밀봉한 뒤 aluminum foil을 두 겹 감싸 암처리 하였다(Kırmızı et al., 2018). 4°C의 항온 생장상(WIM-RL4, Daihan Scientific Co., Wonju, Korea)에서 각각 0(대조구), 3, 6주간 처리한 후 25°C의 항온 생장상에서 배양하며 발아율을 조사하였다. 25°C 항온 생장상 내의 광도는 약 10µmol·m-2·s-1 조건으로 16시간 명조건을 유지하였다.
GA3 처리
온도별 발아실험에서 최종 발아율이 낮았던 봉래꼬리풀 종자를 대상으로 GA3 처리에 의한 휴면 타파 효과를 알아보고자 실험을 실시하였다. 소독한 종자는 0(대조구), 50, 100, 1,000mg·L-1의 GA3 용액에 24시간 동안 암조건에서 침지 처리한 뒤 증류수로 5회 이상 수세하였다. 처리된 종자는 90×15mm petridish에 여과지 2매를 깔고 여과지가 마르지 않을 정도의 증류수를 주입하였다. 25°C의 항온 생장상 내의 10µmol·m-2·s-1 광조건에서 16시간 명조건을 유지하며 배양 후 발아율을 조사하였다.
통계분석
실험은 20립 3반복으로 수행하였고, 온도 처리, 저온 층적처리, GA3 처리에 대하여 수집된 자료는 Sigma Plot 10.0(SPSS Inc., Chicago, IL, USA)을 이용하여 그래프로 제시하였다. 실험의 결과는 SAS 9.4(SAS Inst. Inc., Cary, NC, USA)를 이용하여 분산분석(ANOVA)을 하였고, 처리의 평균간 통계적인 유의성은 Tukey’s honestly significant difference test(p < 0.05)를 사용하여 검정하였다.
결 과
Veronica속 8종의 종자 외부형태를 관찰하였을 때 약 1mm 정도의 크기로 종피는 옅은 갈색을 띄는 것을 관찰하였다(Table 2 and 3). 종자의 길이와 배의 길이를 조사한 결과 같은 종이더라도 길이의 차이가 있기 때문에, 종자 길이에 대한 배의 상대적인 크기를 명확하게 표현할 수 있는 E:S ratio(Vandelook et al., 2009; Chen et al., 2013)를 제시하였다(Table 3). 큰산꼬리풀 종자의 길이는 1.41mm로 Veronica속 8종의 종자 길이 중 가장 길게 측정되었으며, 큰구와꼬리풀 종자의 길이는 1.11mm로 가장 작게 측정되었다. 봉래꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰산꼬리풀의 초기 E:S ratio는 약 0.52로 확인되었다. 둥근산꼬리풀 종자의 초기 E:S ratio가 약 0.61인 반면 구와꼬리풀, 큰구와꼬리풀 종자가 각각 0.46와 0.47이였기 때문에 동일 속 내에서도 배의 비중이 다름을 알 수 있었다. 종피가 갈라지는 시점에 E:S ratio를 조사한 결과, Veronica속 8종 모두 E:S ratio가 18.8 - 58.0% 정도 신장한 후 유근이 돌출되었다(Table 2 and 3).
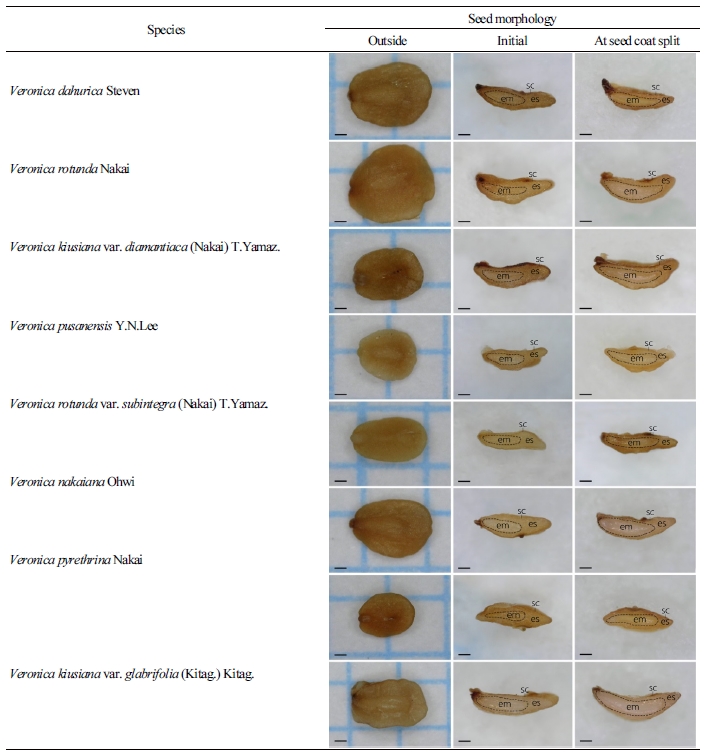
Table 3. Seed length, embryo length, and ratio of embryo length to seed length (E:S ratio) in seed of eight Veronica species 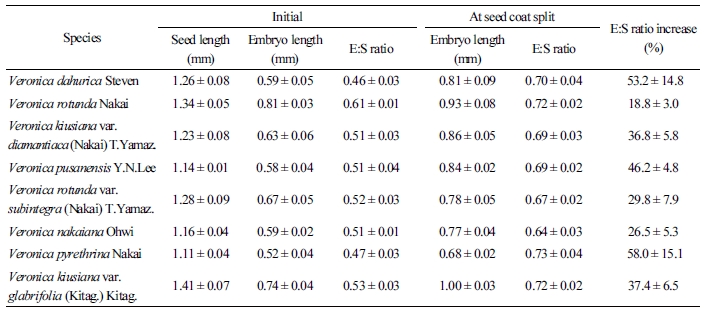 | |
Data are mean ± SE of five replications. | |
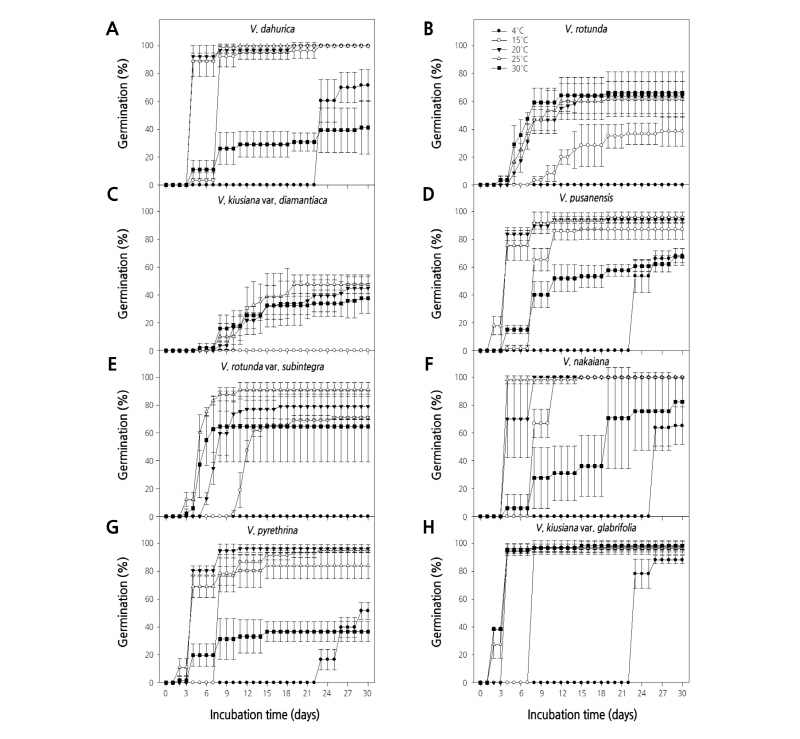
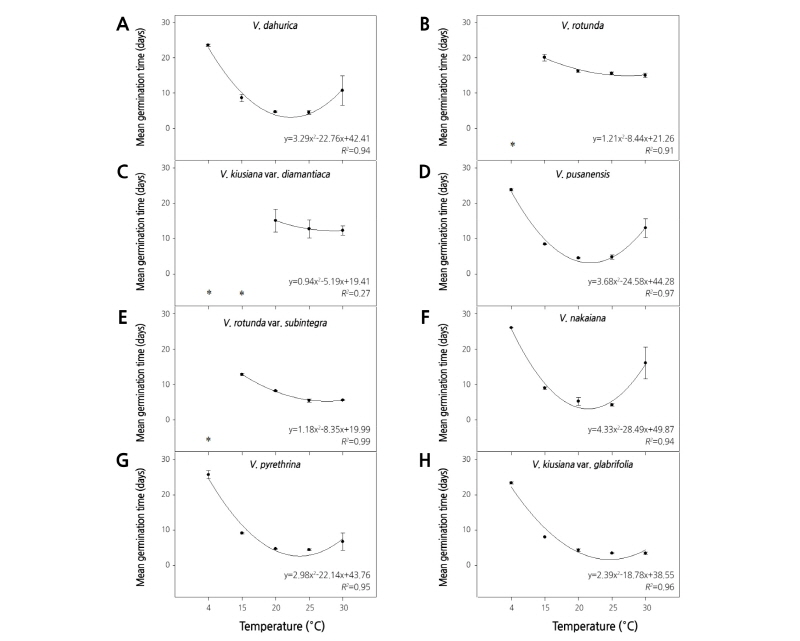
온도별 발아실험 결과, 구와꼬리풀 종자는 20, 25°C 온도처리에서 평균발아소요일수 4.5일을 보이며 100%의 최종발아율을 보인 반면, 30°C 온도처리에선 평균발아소요일수 23.6일을 보이며 41.2%의 최종발아율을 보였다(Figs. 1A and 2A). 둥근산꼬리풀 종자는 20, 25, 30°C 온도처리에서 약 4일만에 발아하기 시작하여 약 66.1%의 최종발아율을 보였으나, 4°C 온도처리에서는 전혀 발아하지 않았다(Fig. 1B). 둥근산꼬리풀 종자의 평균발아소요일수는 20, 25, 30°C 온도처리에서 각각 8.5, 7.6, 6.6일로 조사되어 20 - 30°C 온도범위에서 열흘내에 발아하는 것을 확인하였다(Fig. 2B). 봉래꼬리풀 종자는 20, 25, 30°C 온도처리에서 평균발아소요일수가 각각 15.2, 12.8, 12.3일을 보이고 최종발아율은 각각 44.9, 47.6, 37.6%을 보였다. 4, 15°C 온도처리에서는 전혀 발아하지 않았다(Figs. 1C and 2C). 부산꼬리풀 종자는 4 - 30°C 모든 온도처리에서 최종발아율이 70.0% 이상이었으며, 20, 25°C 온도처리에서 94.0% 이상 발아하였다(Fig. 1D). 부산꼬리풀 종자의 평균 발아소요일수는 4°C 온도처리에서 23.9일을 보인 반면 20, 25°C 온도처리에서 각각 4.6, 4.8일을 보여 평균발아소요일수가 4°C보다 약 19.2일 단축되었다(Fig. 2D). 산꼬리풀 종자는 20, 25°C 온도처리에서 각각 6, 3일차에 발아하기 시작하여 79.1, 91.3%의 최종발아율을 보인 반면, 4°C 온도처리에서 전혀 발아하지 않았다(Fig. 1E). 산꼬리풀 종자의 평균발아소요일수는 20, 25, 30°C 온도처리에서 각각 8.1, 5.4, 5.5일을 보여 20 - 30°C 온도범위에서 열흘 이내에 발아하는 것으로 조사되었다(Fig. 2E). 섬꼬리풀 종자는 20, 25°C 온도처리에서 평균발아소요일수 각각 5.2, 4.2일로 100%의 최종발아율을 보인 반면, 4°C 온도처리에서 26일차에 발아하기 시작하여 65.3%의 최종발아율을 보였다(Figs. 1F and 2F). 큰구와꼬리풀 종자는 15, 20, 25°C 온도처리에서 각각 95.0, 96.5, 82.3%의 최종발아율을 보였으나 30°C 온도처리에서 36.6%의 최종발아율을 보였다(Fig. 1G). 큰구와꼬리풀 종자는 4°C 온도처리에서 평균발아소요일수 25.6일을 보인 반면 15, 20, 25°C 온도처리에서 각각 9.1, 4.7, 4.4일로 평균발아소요일수가 단축되었다(Fig. 2G). 큰산꼬리풀 종자의 평균발아소요일수는 4, 15, 20, 25, 30°C 온도처리에서 각각 23.3, 8.0, 4.3, 3.4, 3.4일이었으며 최종발아율은 모든 온도처리에서 88.0% 이상으로 나타났다(Figs. 1H and 2H). 큰산꼬리풀 종자는 4, 15, 20, 25, 30°C 온도처리에서 각각 23, 8, 4, 2, 2일차에 발아하기 시작하여 온도가 높을수록 발아시작이 빨라졌다. 따라서 위의 결과를 통해, Veronica속 8종의 종자발아를 위한 최적의 온도조건은 20 - 25°C 범위인 것으로 확인하였다.
온도별 발아실험에서 최종발아율이 낮았던 봉래꼬리풀의 휴면타파를 위해 저온 층적처리 기간 및 농도별 GA3 처리를 실시하였다. 봉래꼬리풀 종자에 기간별로 저온 층적처리한 결과, 3주 및 6주간 처리를 하였을 때 최종발아율은 각각 75.3, 80.3%를 보여 대조구에서 조사된 45.0%의 최종발아율보다 각각 30.3, 35.3% 향상되었다(Fig. 3A). GA3를 처리한 결과 50, 100, 1,000mg·L-1의 농도에서 5일차부터 발아하기 시작하여 각각 71.3, 87.2, 91.7%의 최종발아율을 보여 GA3 0mg·L-1의 대조구에서 조사된 44.7%의 최종발아율보다 각각 26.6, 42.5, 47.0% 향상되었다(Fig. 3B).
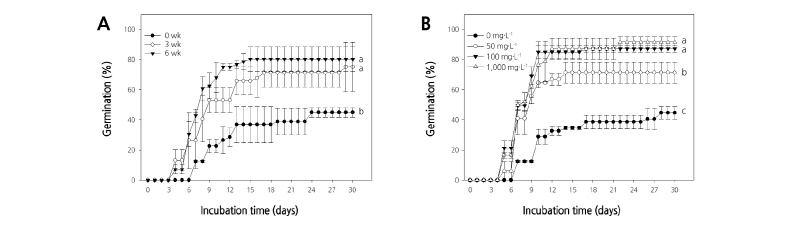
Fig. 3. Germination response according to cold stratification treatments (0, 3, and 6 weeks at 4°C) (A) and GA3 soaking treatments (0, 50, 100, or 1,000 mg·L-1) (B) of Veronica kiusiana var. diamantiaca seeds at 25°C for 30 days. Error bars indicate the mean ± SE from three replications. The different letters represent statistically significant differences, as determined by Tukey’s HSD tests (p < 0.05).
고 찰
종자는 내부 배의 형태에 따라 크게 기저형(basal), 지협형(peripheral) 및 중축형(axile)의 세 가지로 나뉜다(Martin, 1946). 기저형은 배가 상대적으로 작고 종자 크기의 1/2을 넘지 않으며, 종자의 하단부에 위치한다. 지협형은 배가 길고 크며, 종피를 따라 휘어 있는 형태이다. 중축형은 배가 작은 것부터 큰 것까지 다양하지만 배가 종자 중앙부에 길게 위치한 선형(linear type)이 대표적인 예로 보고하였다. 본 연구 결과, Veronica속 8종의 배의 형태는 종자 크기의 약 52% 정도 되는 선형(linear type)으로 판단되었다(Table 2 and 3). Martin(1946)은 중축형(axile)의 종자들 중에서 크기가 작은 것을 miniature subdivision으로 나누고 이를 다시 dwarf type(0.3-2mm)과 micro type(0.2mm 이하)의 두 가지로 분류하였다. 본 실험에서 종자의 길이는 1mm 내외로 조사되었기 때문에, dwarf type으로 분류할 수 있었다.
종자가 미숙배를 가지는지는 채종 시점에 육안으로 확인되는 상대적 크기가 아니라 발아직전까지의 추가적인 신장이 일어나는지가 가장 중요한 지표이다(Baskin and Baskin, 2014). 예를 들면, Drosera anglica의 경우, 초기에 약 0.24mm의 상당히 작은 배를 가지고 있었으나, 추가적인 배의 신장 없이 곧바로 발아하였기 때문에 완전히 발달된 배로 판단하였다(Baskin and Baskin, 2005). 반면에, Nothapodytes nimmoniana 종자의 경우, 초기 배의 길이가 종자길이의 약 74%로 상당히 컸지만, 발아 직전까지 약 17% 추가적인 증가를 확인하여 미숙배를 가지는 형태적휴면(MD)으로 분류하였다(Chen et al., 2015). 뿐만 아니라, Dianella sandwicensis(Xanthorrhoeaceae)의 경우도 초기 배의 크기가 종자의 약 78% 정도로 상당히 컸지만, 발아직전까지 약 16% 추가로 증가하여 미숙배로 판단하였다(Wolkis et al., 2018). 이는 생태생리적인 관점에서, 발아단계로 전이되기 이전의 일종의 지연시간으로 판단된다(Baskin and Baskin, 2014). 본 연구에서, Veronica속 8종간에 차이는 있었지만, 초기 배의 크기에 비해서 18.8 - 58.0% 정도의 추가적인 신장이 일어난 다음에 발아단계로 전이되었기 때문에, 대상종 모두 미숙배를 가진다고 판단할 수 있다. Baskin and Baskin(2007)은 Martin의 ‘seed classification system’을 재검토하면서, dwarf-seed type(0.3-2mm)의 경우 초기 E:S ratio가 0.5 정도 또는 그 이하일 경우, ‘linear underdeveloped’로 분류하여 본 연구 결과를 뒷받침해 준다.
미숙배로 분류된 종자는 발아하기 전에 배가 일정 크기 이상 자라야 하는데, 적절한 조건에서 30일 이내에 배의 신장과 발아가 이루어질 경우 이를 형태적휴면(MD)이라 한다. 반면 발아기간이 더 오래 걸리거나 저온습윤, 고온습윤, 또는 복합처리를 해야만 배가 신장하고 발아되는 경우를 형태생리적휴면(MPD)이라 분류한다(Baskin and Baskin, 2004). 따라서 한국 자생 Veronica속 중 구와꼬리풀, 둥근산꼬리풀, 봉래꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰구와꼬리풀, 큰산꼬리풀은 미성숙 배를 가지고 있지만 적절한 조건에서 30일 이내에 각각 100%, 66.1%, 47.6%, 94.0%, 91.3%, 100%, 96.5%, 98.4%가 발아하였기 때문에(Fig. 1), 구와꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰구와꼬리풀, 큰산꼬리풀 종자는 형태적휴면임을 확인하였다. 둥근산꼬리풀 종자는 약 66% 정도가 형태적휴면임을 알 수 있었다. 봉래꼬리풀 종자는 약 48% 정도가 형태적휴면(MD)이며 GA3 1,000 mg·L-1 처리시 발아율이 92%이므로 약 44% 정도가 형태생리적휴면(MPD)임을 확인하였다.
형태생리적휴면은 배발달 온도가 상대적 고온 범위(15 - 20°C)인 simple-type MPD와 상대적 저온 범위(0 - 10°C)인 complex-type MPD로 나눌 수 있다(Geneve 2003; Baskin and Baskin 1998, 2004). 특히, 봉래꼬리풀은 상대적 고온인 20 - 25°C 범위에서 배가 발달하여 E:S ratio가 초기에 비해 증가하였고 발아가 촉진되었기 때문에 형태생리적휴면 중 simple-type MPD로 판단되었다. 형태생리적휴면(MPD)은 형태적휴면(MD)과 생리적휴면(PD)이 복합적으로 작용하는 특성인데, 특히 생리적휴면의 깊이에 따라 nondeep, intermediate, deep type의 세 가지로 나눌 수 있다(Baskin and Baskin 1998, 2004; Geneve 2003). Nondeep PD는 짧은 기간(몇 일 - 2개월 정도)의 저온 층적처리, GA 처리로 휴면이 쉽게 타파되며, 후숙(dry after-ripening) 효과가 있는 것이 많다. Intermediate PD는 최소 2 - 3개월의 저온 층적처리가 필요하고, GA 처리로 모든 종의 휴면을 타파시키지는 못한다. 뿐만 아니라, 후숙 처리는 저온 층적처리 소요 기간을 단축시키는 특징을 가지고 있다. Deep PD는 GA 처리 효과가 없으며, 최소 3 - 4개월의 저온 층적처리 기간이 필요하다. 본 실험에서 봉래꼬리풀 종자는 6주간의 저온 층적처리와 GA 처리로 효과적으로 휴면이 타파되었기 때문에, nondeep simple MPD로 분류할 수 있었다. Guerin et al.(2013)은 Veronica속 식물 중 V. parnkalliana 종자가 형태생리적휴면을 가진다고 보고하여 본 연구의 둥근산꼬리풀과 봉래꼬리풀의 결과와 유사하였다. 그러나 Baskin and Baskin(2014)은 V. americana, V. anagallis-aquatica, V. ciliate, V. eriogyne, V. szechuanica, V. rockii, V. alpina 등이 생리적휴면을 보인다고 정리하였기 때문에, 한국 자생종들과는 다름을 알 수 있었다.
GA/ABA 비율, 민감도는 종자의 휴면과 발아를 조절하는 중요한 요소로 알려져 있다(Chien et al., 1998; Finch-Savage and Leubner-Metzger, 2006; Cho et al., 2018). 생합성 된 ABA 함량이 증가하거나 민감도가 증가하면 휴면이 유도되고, 반대로 GA 함량이 증가하거나 ABA에 대한 민감도가 떨어지면 휴면이 타파되어 발아가 촉진되는 경향이 있다. 본 연구에서 봉래꼬리풀 종자에 저온 층적처리 시 처리기간이 길어질수록 발아율과 발아속도가 향상되었다(Fig. 3). Taxus mairei종자는 채종 직후 종자당 ABA 함량이 8,888pg이였지만 5°C에서 저온 층적처리 시 종자당 ABA 함량이 536pg까지 낮아졌다고 보고되었다(Chien et al., 1998). 저온 층적처리는 종자의 내생 ABA 함량을 감소시킬 뿐 아니라 GA의 합성을 촉진시키고 배의 정단분열조직의 세포분열을 촉진시켜 발아를 촉진시킨다. Veronica parnkalliana 종자는 5 - 40°C의 온도 범위에서 종자가 전혀 발아하지 않았지만, 250mg·L-1 GA3 처리 시 80 - 90% 발아율이 향상된 바 있다(Guerin et al., 2013). 봉래꼬리풀 종자에 GA3 100mg·L-1과 GA3 1,000mg·L-1 처리 시 최종발아율은 각각 87.2%와 91.7%로 무처리와 GA3 50mg·L-1 처리에 비해 유의하게 높게 발아하였다(Fig. 3). 따라서 저온 층적처리와 GA 처리는 휴면 타파에 효과적이었다. 효율 측면을 고려하면, 봉래꼬리풀 종자의 휴면타파를 위한 최적의 조건은 GA3 100mg·L-1 처리로 판단된다.
본 실험 결과, 같은 속의 식물이지만 일부 종에서 다소 차이가 나는 휴면유형이 관찰되었다. Baskin and Baskin(1998)은 Veronica arvensis종자와 V. peregrine종자가 휴면을 하지 않거나(non-dormancy type) 환경에 따라 휴면을 하는(conditional dormancy type) 종이라 분류하여 같은 속에서도 휴면유형이 완전히 다를 수 있다고 보고하였다. Lamium속 중 스웨덴(Sweden)에 자생하는 L. amplexicaule 종자는 10, 15, 20, 25°C 온도조건에서 거의 발아하지 않았기 때문에 휴면이 깊었으나, 미국 켄터키주(Kentucky)에 자생하는 L. amplexicaule 종자는 휴면이 얕았는데, 이는 각 지역의 환경에 식물이 적응했기 때문으로 해석하였다(Karlsson and Milberg, 2008). 이처럼 종자 휴면 깊이의 차이는 종들의 지역 적응을 돕고 같은 종 사이에서도 휴면 양상이 다를 수 있다고 보고되고 있다. Aristolochia 속의 A. macrophylla, A. tomentosa, A. manshuriensis, A. californica 식물의 종자는 미숙배를 지녀 형태적휴면이나 형태생리적휴면을 모두 지닌다고 보고된 바 있다(Baskin and Baskin 2004; Adams 2005b). 미국 캘리포니아주의 특산식물인 A. californica는 미숙배의 성장과 종자 발아를 위해서 상대적 저온인 10°C에서 16주 후 74% 발아율을 보여 complex-type MPD로 분류되었다(Adams et al., 2005a). 반면 A. macrophylla, A. tomentosa, A. manshuriensis 종은 상대적 고온에서 발아하여 형태적휴면(MD)과 nondeep simple-type MPD로 분류된 바 있다. 이는 종마다 생존하는 과정에서 서로 다른 휴면유형을 발달시켜왔으며, 각각의 휴면유형 내에서도 깊이를 달리하는 전략을 선택적으로 활용해왔음을 의미한다(Baskin and Baskin, 2004; Adams et al., 2005a). 본 연구 대상종인 봉래꼬리풀의 경우, 남한의 강원도 설악산 지역에 한정적으로 분포하고 있는 것으로 알려져 있는데, 휴면타파를 위한 저온 요구도와 휴면유형이 지리적 분포에 영향을 받은 것으로 판단된다. Carta et al.(2014)은 자생 Crocus속의 종자휴면 연구에서, 서로 다른 고온, 저온 요구도가 지리적 분포와 밀접한 연관이 있다고 보고한 바 있다.
결론적으로, Veronica속 8종(구와꼬리풀, 둥근산꼬리풀, 봉래꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰구와꼬리풀, 큰산꼬리풀)의 발아 적온은 20 - 25°C 범위로 판단되었다. 특히 봉래꼬리풀은 모든 온도조건에서 50% 미만의 발아율을 나타냈으나 저온층적 3 - 6주 처리, GA3 100 - 1,000mg·L-1 처리 시 휴면이 타파되었다. 구와꼬리풀, 둥근산꼬리풀, 부산꼬리풀, 산꼬리풀, 섬꼬리풀, 큰구와꼬리풀, 큰산꼬리풀 종자는 형태적휴면을 지니며 봉래꼬리풀 종자는 형태적휴면과 형태생리적휴면을 동시에 지니고 있었다. Veronica속 내에서도 휴면의 유형이 다를 수 있음을 확인하였고, 다른 대륙 및 나라에 자생하는 Veronica속 식물의 휴면유형과도 차이 나는 것을 확인하였다.
