

Introduction
Materials and Methods
Plant Materials
Carotenoid Extraction
HPLC Analysis of Carotenoids
Statistical Analysis
Results
Diversity of Carotenoid Profiles Among Tea Cultivars
Profile of Carotenoid Diversity Based on the Concentration of Each Compound
Discussion
Introduction
Tea is a perennial woody plant belonging to the Camelliagenus and the Theaceae family. It is used to produce a range of beverages from green tea, which is non-fermented, to black tea, which is fully fermented by a variety of processes (Takeo and Hampton, 1992a, 1992b). Tea consumer preference is largely dependent on the flavor of prepared tea beverages (Ravichandran and Parthiban, 1998). The flavor of tea can be divided into two groups, namely aroma (volatile compounds) and taste (non-volatile compounds) (Ho et al., 2015). Tea aroma, which is composed of volatile flavor compounds (VFCs) generated during tea processing, is an important quality parameter and determines the market of prepared tea beverages. These VFCs can be classified into two categories, specifically the products of lipid breakdown and the derivatives of terpenoids, carotenoids, and amino acids. TheVFC profile has considerable impact on the flavor of prepared tea (Ravichandran and Parthiban, 2000; Ravichandran, 2002; Lee et al., 2013) and depends on many factors, including plant cultivar, environmental conditions, agronomic practices, harvest time, and processing methods (Yang et al., 2013). Many studies have been conducted to characterize this complex picture from different aspects; however, most studies have focused on the metabolic pathways of VFCs and their detection and identification methods. It is worth noting that different tea cultivars differ greatly in VFC profile, but this remains relatively less characterized (Schuh and Schieberle, 2006; Rawat and Gulati, 2008).
Carotenoids are a large group of pigments produced in plants that display colors ranging from yellow to red. Carotenoids synthesized in the fresh tea leaf are important precursors of tea volatile flavor compounds, of which some are amongst the most potent flavor components and significantly contribute to tea quality (Kumazawa and Masuda, 2002; Yang et al., 2009). In particular, β-ionone and damascenone are important C13-carotenoid-derived compounds that have low human odor perception thresholds. Significantly contribute to aroma in black tea (Ravichandran, 2002). Previous studies have identified carotenoids, including β-carotene, lutein, zeaxanthin, and others, as aroma precursors in the tea leaves (Chen and Zhou, 2005; Ho et al., 2015). Quantitative and qualitative analyses of carotenoids are of great importance for both scientific and commercial interests because of their contribution to the quality of prepared tea.
As the center of origin for the tea plant, China has accumulated abundant and diverse tea germplasms. The identification and evaluation of these germplasms, especially elite cultivars, is fundamentally important for cultivar development and processing and for improving extraction of bioactive compounds such as carotenoids (Felfe et al., 2011). A previous study reported the differences in β-carotene and lutein contents among 119 tea germplasms from four countries (Wang et al., 2010). However, the carotenoid compositions of a number of elite Chinese tea cultivars are not fully elucidated. This study was performed to better understand the carotenoid profile of 31 select elite tea cultivars so they could be used effectively for breeding and production. The main objectives of this study are: (1) to understand the variability of carotenoid contents in tea plants originating from different tea-producing regions of China; (2) to reveal the profile and the variation in contents of the major carotenoids produced by the 31 elite Chinese tea cultivars under the same growth environment; (3) to identify the key steps in the carotenoid biosynthetic pathway involved in differential carotenoid accumulation among the 31 tea cultivars.
Materials and Methods
Plant Materials
A total of 31 elite tea cultivars originated from China were selected for analyses (Table 1). These cultivars were cultivated in the same environment with same agronomic practices at the Tea Research Institute, Chongqing Academy of Agricultural Sciences at Yongchuan, Chongqing, China. Previous research results indicated that carotenoids were at high levels in autumn (Yang et al., 2013). To analyze the main carotenoids, 20 pieces of “one bud and two leaves” of the first flush in autumn (September) were harvested from 3 individual plants of each cultivar in 2014. Thus, 3 samples of 20 “one bud and two leaves” were individually collected for each cultivar. All samples were freeze dried at -50°C, ground to a fine powder, and stored at -20°C until analysis.
Carotenoid Extraction
Carotenoid extraction followed the protocol described by Lee and Castle (2001) with slight modifications. In brief, the tea powder (0.5 g) was homogenized in 25 mL of extracting solvent (hexane/acetone/ethanol, 50/25/25, v/v) in screw-top tubes. Samples were then centrifuged for 5 min at 6,500 rpm at 5°C (AllegraTM64R Centrifuge, Beckman Coulter TM, USA). The top colored layer of hexane was recovered and transferred to a 25 mL volumetric flask, then dried under nitrogen gas with a bath-type nitrogen blowing instrument (Ferren Technology Co., Ltd).
For saponification, the residue was re-dissolved with 2 mL of methyl tert-butyl ether (MTBE), and placed in a 15 mL culture tube to which 2 mL of 10% methanolic KOH solution was added. The tubes were wrapped with aluminum foil to protect from light, and were shaken overnight at room temperature. Samples were transferred to a separator funnel to which 5 mL of water and 2 mL of 0.1% BHT (butylated hydroxytoluene)/MTBE were added. The colored layer was collected and dried under nitrogen. The residue was dissolved in 2 mL methanol/acetone (2:1, v/v). Samples were filtered through a Millipore 0.5 µm filter before injection into the high-performance liquid chromatography (HPLC) apparatus. Analyses were carried out under red light to avoid carotenoid degradation during extraction and saponification.
HPLC Analysis of Carotenoids
Carotenoids were analyzed by HPLC using a Waters High Performance Liquid Chromatographic Acquity system (Milford, MA, USA) and separated along a C30 column (250 mm×4.6 mm i.d., 5 µm) (YMC, Wilmington, NC, USA). The system was operated by the Waters Empower software (Waters Corporation). HPLC conditions were conducted according to the method of Mouly et al.(1999). The gradient profile and the mobile phase composition were given in Table 2. The carotenoids were identified by comparing their retention time and the UV-visible spectra of their peaks with the standards. The standards of lutein, β-carotene, zeaxanthin, β-cryptoxanthin, and α-carotene were obtained from Sigma (St. Louis, MO, USA). The carotenoids with standards were quantified by a calibration curve; all-E-violaxanthin and Z-β-carotene detected were quantified by calibration curve of β-carotene. Three biological replicates were used for all samples.
Table 2. Gradient profile used in the liquid chromatographic separation of carotenoids
 | |
zLinear gradient yMethyl tert.-butyl ether | |
Statistical Analysis
The samples from individual plant were analyzed in triplicates and averaged. All data were expressed as mean ± standard deviation. Statistical analysis was performed using the SPSS v19.0 software (SPSS Inc. Chicago, IL, USA). Significant differences among the samples were calculated using one-way ANOVA followed by Duncan’s multiple-range test at the 5% level (p≤0.05). PCA was performed using SPSS software. Hierarchical cluster analysis was performed using Ward’s method with JMP software (http://www.jmp.com).
Results
Diversity of Carotenoid Profiles Among Tea Cultivars
For each of the 31 cultivars, three leaf samples (each consisting of 20 pieces of first flush material) collected from three plants were analyzed by HPLC. Seven carotenoids were detected, and their chromatographic and spectral characteristics are presented in Table 3 and Fig. 1. The mean, range, and coefficient of variation (CV) for carotenoid contents in new shoots
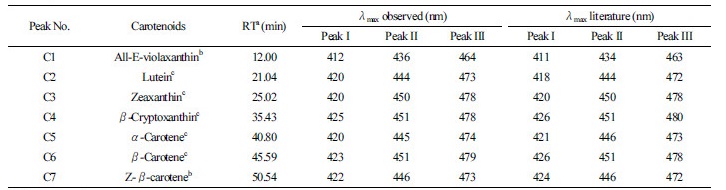
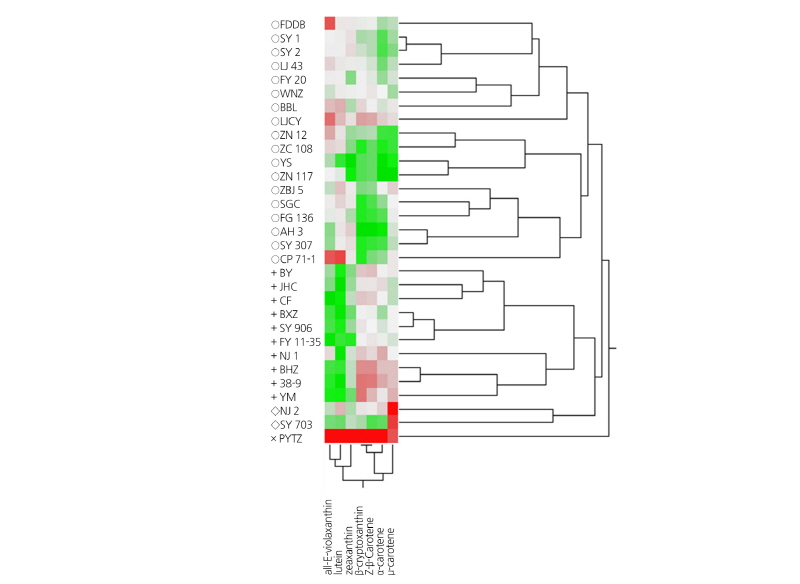
Fig. 1.
Clusteranalysisofcarotenoidcontentsof31elite Chinese tea cultivars.Thecarotenoidcontentsindifferentcultivars were sorted using JMP software (http://www.jmp.com/) on the basis of the 7 measured carotenoids shown across the bottom. Coloring of green to white to red indicates increasing levels of carotenoid content.
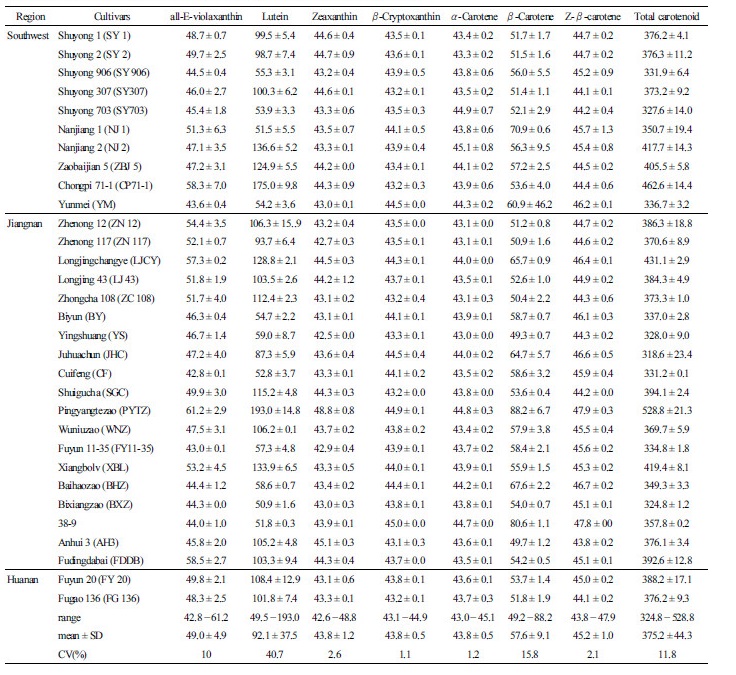
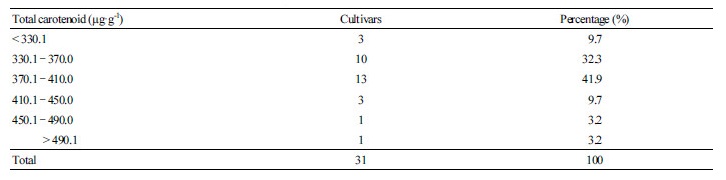
among the 31 cultivars are summarized in Table 4. The average total carotenoid content was 375.2 ± 44.3 µg·g-1, varying from
324.8 to 528.8 µg·g-1, and that of the 26 cultivars ranged from 330.0 to 450.0 µg·g-1 (Tables 4 and 5). The highest average contents of all-E-violaxanthin (61.2 µg·g-1), lutein (193.0 µg·g-1), zeaxanthin (48.8 µg·g-1), β-cryptoxanthin (44.9 µg·g-1), β-carotene (88.2 µg·g-1), Z-β-carotene (47.9 µg·g-1) were measured in Pingyangtezao (PYTZ) cultivar (Table 4). Consequently, a very strong effect of genotype was attributed to the variability of each component.
Profile of Carotenoid Diversity Based on the Concentration of Each Compound
All quantitative data for carotenoids detected in the 31 cultivars were used for cluster analysis (CA) and principal component analysis (PCA). As shown in Fig. 1, three main clusters were observed. Specifically, cluster 1(‘o’ annotation) displayed higher concentrations of lutein than β-carotene, cluster 2 (‘+’ annotation) displayed higher concentrations of β-carotene than lutein, and cluster 3 (‘x’ and ‘◊’ annotations) displayed high concentrations of both lutein and β-carotene. Lutein and β-carotene appeared to broadly contribute to the total amount of carotenoids. Thus, differences between the 31 genotypes were qualitative and quantitative.
PCA was performed using the same data as used in CA. The loading scatter plot showed the differences/similarities between various tea cultivars, which was based on comprehensive carotenoid contents (Fig. 2). The first two principal components described 99.23% of the total variance. Those carotenoids that accounted for maximum variance in the data set were given more weight or loading. Therefore, lutein (on PC1) and β-carotene (on PC2) were the most differentiating carotenoids (Fig. 2). The PCA load plot was overlaid on the PCA score plot, which indicated the relationship between heavily weighted carotenoids and cultivar groups. The PCA model indicated two centers primarily but not exclusively, namely Center I that had “Yunmei” (YM) as the central cultivar with negative values on PC1 and Center II that had “Fudingdabai” (FDDB) as the central cultivar with positive values on PC1. As shown in Table 1, YM was a single line selected from Yunnandaye group, and FDDB was a single line selected from Fuding group. In Center I, all cultivars aside from NJ1 and BHZ were hybrid
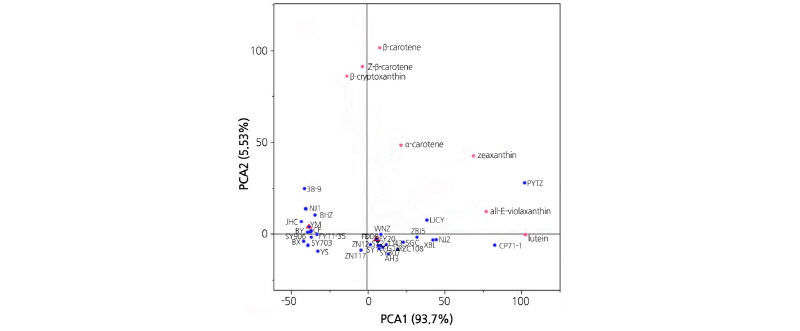
off-springs of Yunnandaye either as paternal or maternal material. In Center II, the cluster was more complicated, but it showed those hybrid off-springs of FDDB, including Fugao 136 (FG136), Fuyun 20 (FY20), Zhenong 12 (ZN12), had score values on PC1 close to FDDB. Interestingly, most of those cultivars belonging to the single line selected from different origins were positively distinguished from those two central cultivars (i.e., YM and FDDB), including Nanjing 1 (NJ 1), Nanjing 2 (NJ 2), Zaobaijian 5 (ZBJ 5), Baihaozao (BHZ), Chongpi 71-1 (CP 71-1), Longjingchangye (LJCY), Pingyangtezao (PYTZ), Xiangbolv (XBL), and Anhui 3 (AH 3). Thus, the analyses revealed more interspecific differences than intraspecific differences.
Discussion
In this study, the carotenoid contents of 31 tea cultivars from three main tea-producing regions in China, including southwest region, Jiangnan region, and Huanan region, were assayed and analyzed (Table 6). There was no significant difference in the detected carotenoids among the three tea-producing regions (p> 0.05), which was consistent with the results of PCA. This result might be due to the limited experimental material; however, a previous investigation found no significant difference in β-carotene and lutein contents among 119 germplasms from four countries (Wang et al., 2010). Nevertheless, cultivars from Huanan region had the highest average lutein content (99.7 µg·g-1), and cultivars from Jiangnan region had the highest average β-carotene content (59.2 µg·g-1). Cultivars from Jiangnan region had the highest CV in all the detected carotenoids. As shown in Table 1, several cultivars from different producing regions had the same parent(s), namely Fudingdabai or Yunnandaye or both. Shared parents may explain why no significant differences were observed in carotenoid contents among the cultivars from different tea-producing regions.
We analyzed carotenoid composition of the 31 genotypes to evaluate the contribution of genotype diversity to carotenoid profiles. Because the large sample amounts were analyzed (60 of “one bud and two leaves” per genotype) and several extraction/saponification/HPLC analyses were performed for each genotype, we can conclude that the contribution of genotype to the variance in carotenoid composition is very high. Therefore, carotenoid profile is highly influenced by genetic factors whereas other sources of variation such as growing conditions, geographical origin, leaf maturity, and method of
Table 6. Carotenoid profiles of tea cultivars from different producing regions
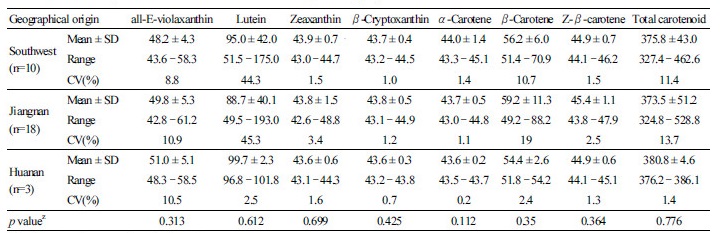 | |
zThe F-test results among 3 producing regions. | |
analysis are minimal. Thus, we can analyze more accurately the relationships between genetic and carotenoid diversities.
In the present study, lutein and β-carotene were found to be the main carotenoids present in the 31 tea cultivars (Table 4, Fig. 1), which is in agreement with previous findings using different cultivars (Shao, 2003). In China, Yunnandaye and Fudingdabai are two excellent commercial cultivars used for producing different kinds of tea products and breeding materials and have thus been introduced to different tea-producing regions. In this study, more than half (n=18) of the samples are derivatives of the two cultivars (Table 1). According to PCA (Fig. 2), results indicated that the hybridization with Yunnandaye or Fudingdabai resulted in indistinguishable carotenoid accumulation in parental and offspring material. One exception was the 38-9, a hybrid offspring of Fudingdabai × Dayechangqing, which was distinguishable from the two centers by a positive high score value on PC2, suggesting that Dayechangqing is be an excellent material for high carotenoid breeding. Those single lines selected from different origins may also be used as high carotenoid breeding parents. Taken together with previous findings (Wang et al., 2010), our results suggest the carotenoid contents of tea leaves are highly influenced by genetic factors rather than cultivation factors.
One purpose of this study was to identify key steps of the carotenoid biosynthetic pathway involved in differential carotenoid accumulation among the31 eliteChinese teacultivars. Based onthe quantitativedataof all carotenoids, threemain clusters were indicated by CA (Fig. 1). The behavior of genotypes of each cluster is illustrated in Fig. 3, where each arrow shows which pigments were synthesized. Genotypes of cluster 1 produced higher lutein than other carotenoid types, genotypes of cluster 2 produced higher β-carotene than other carotenoid types, and genotypes of cluster 3 produced higher total carotenoids.
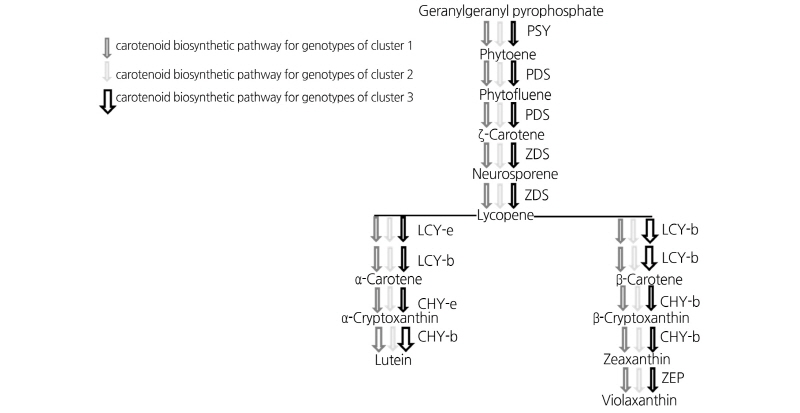
Fig. 3.
Carotenoid biosynthetic pathway obtained through the qualitative carotenoid composition in 31 elite Chinese tea genotypes. Behaviors of genotypes of each cluster are represented by different arrows. PSY, phytoene synthase; PDS, phytoene desaturase; ZDS, ζ-Carotene desaturase; LCY-b, lycopene β-cyclase; LCY-e, lycopene ε-cyclase; CHY-e, ε -carotene hydroxylase; CHY-b, β-carotene hydroxylase; ZEP, zeaxanthin epoxidase.
For cluster 1, our results lead us to hypothesize that key steps are the transformation of α-carotene into α-cryptoxanthin and lutein by hydroxylation catalyzed by ε-carotene hydroxylase (CHY-e) and β-carotene hydroxylase (CHY-b). Concerning the genotypes of cluster 2, qualitative data and quantitative data (Table 4 and Fig. 3), the latter in particular, suggested that the most important step is the formation of β-carotene from lycopene by the action of lycopene ε-cyclase (LCY-e) and lycopene β-cyclase (LCY-b). It should be noted that for all of the genotypes of cluster 2, total carotenoid contents were lower than that for genotypes in cluster 1 and cluster 3. Therefore, from a quantitative point of view, the relatively low amount of total carotenoid suggests that the most important step is the formation of phytoene from geranylgeranyl diphosphate (GGPP) by the action of a phytoene synthase (PSY). For the genotypes of cluster 3, the high accumulation of both lutein and β-carotene suggests there is high enzymatic activity of those enzymes, such as PSY, PDS, ZDS, LCYs, and CHYs, situated upstream in the carotenoid biosynthetic pathway. It is worth noting that the CHYs are active on the left branch of the pathway to synthesize lutein, but are not active on the right branch to synthesize β-cryptoxanthin (Fig. 3). According to previous research, two different types of carotenoid hydroxylases (CHYs) have been found in plants, namely non-heme di-iron β-carotenoid hydroxylases (BCH type) and cytochrome P450 enzymes (CYP97 type). In Arabidopsis, there are at least five genes encoding CHYs, specifically two BCH-type CHYs which encode enzymes that are most active in the hydroxylation of the β-carotene, and three CYP97-type CHYs which encode enzymes that prefer α-carotene as their substrate (Kim et al., 2009; Kim et al., 2010).Thus, it is reasonable to suggest that the active CHYs in the genotypes of cluster 3 are CYP97-type CHYs. To our knowledge, there isno comprehensive study on the carotenoid biosynthetic pathways in tea plants. The present findings suggest there maybe several regulation mechanisms behind carotenoid accumulation in tea plants, indicating that continuing experiments are required.
To concluded, carotenoids area important determinant of tea quality as many of the quality related flavor volatiles are produced through the degradation of carotenoids during tea processing. Here, we present valuable information concerning the carotenoid profiles of 31 elite Chinese tea cultivars, including carotenoid composition and organization. Those cultivars with high carotenoid contents could be used as breeding material for producing fine tea, as well as research material for further understanding of variation in carotenoid production in tea plants.
Moreover, in China, most tea farmers harvest only spring shoots to produce elite teas, and do not harvest in summer and autumn due to low market price of these seasonal teas. Therefore, abundant tea leaves are left on the tea bushes and are not harvested every year. The tea bushes are often pruned after the spring harvesting to prepare a new crown for the next year, leading to high-level wastage of tea leaves. There is potential in developing cultivars with distinguishable high carotenoid contents, especially those with high-level carotenoids in their summer and autumn tea leaves and young stems, for the extraction of carotenoids for use in the food industry. This would lead to an increase in the value of summer and autumn tea leaves and overall tea crop profitability.
