

서 언
휴면은 온대 낙엽수목이 겨울과 같은 불량 환경에서 생존하기 위한 고도의 진화형태로 휴면의 유도는 온도와 광주기를 포함한 다양한 요인에 의해 영향을 받으며(Perez and Lira, 2005), 식물 내한성이나 내동성과 관련된 주요한 생육 단계이다. 자연상태에서 휴면 기작을 통한 눈의 발달과 내재 휴면의 완료를 위해서는 일정기간 저온에 노출되어야 한다(Alburqureque et al., 2008). 특히, 포도나무는 낙엽과수 중 다른 과종과 달리 휴면기간 동안 불충분한 저온 또는 극한의 저온에 노출될 경우, 이듬해 발아하지 않거나 발아기가 늦어지는 등 불규칙한 발아를 초래하고 이는 신초 생장량과 과실 생산량의 감소뿐만 아니라 균일하지 못한 과실의 발달로 이어지는 경우가 많다(Lavee et al., 1984; McColl, 1986). 일반적으로 포도나무의 발아율은 적당한 저온에 대한 노출 증가로 향상되는 경향을 보이고, 지금까지 알려진 바에 의하면 발아에 요구되는 저온 노출시간은 7°C 이하의 온도에서 50-400시간이나, 이는 유전자형에 따라 다르게 나타날 수 있다(Dokoozlian, 1999; Terzo-Martinez et al., 2009). 이러한 저온에 노출된 정도는 휴면 심도를 결정할 수 있는 내생 마커 부재에 대해 발아라는 가시적인 변화로 실제 눈이 가지고 있는 휴면 심도를 결정할 수 있는 이점이 있다. 그러나 지금까지 저온의 축적이나 발아를 추정하는데 사용되는 방법은 제한된 환경조건을 기반으로 하며(Dokoozlian, 1999; Mohamed et al., 2010; Sagredo et al., 2005), 실제 재배 포장에서 눈이 가지고 있는 휴면의 심도나 휴면 기간 중 눈의 발달 정도를 고려하고 있지 않다. 따라서 본 연구는 우리나라 포도의 대표 품종인 ‘Campbell Early’의 발아 특성 관찰을 통해 눈의 휴면 심도 및 내재휴면 타파를 위한 저온 축적 사이의 관계를 알아보고, 저온에 노출되는 동안의 눈 발달을 관찰하여 발아와 휴면과의 관계에 대한 기초 자료를 축적하고자 수행하였다.
재료 및 방법
경기도 안성시 소재 포도원(N36.99o E127.28’)에서 수세가 비슷한 4년생 ‘Campbell Early’ 포도나무를 대상으로 2012년 9월부터 2013년 2월까지 15일 간격으로 1년생 가지를 채취하였다. 시기별 무작위로 10개의 가지를 채취하였으며, 눈이 충실하게 발달된 것으로 생각되는 3-9번째 마디 부위를 사용하였다. 수집된 가지는 남아있는 잎을 제거한 뒤, 70% 에탄올로 1차 소독한 다음 증류수에 수삽하였다. 증류수는 매일 교환해 주었고, agar 등에 의해 물에 잠긴 기부의 물관부가 손상되는 것을 방지하기 위해 7일에 한번씩 기부의 1-1.5mm 정도를 잘라내었다.
발아 조건을 온도 23 ± 2°C, 상대습도 30% 및 광주기 16/8 (day/night)로 일정하게 유지시킨 후 지속적인 육안관찰을 통해 최초 발아까지 소요된 일수와 시기별 발아율을 계산하고, 발아하지 않는 눈의 발아율은 0으로 간주한 뒤 발아의 평균속도 및 최종발아율을 조사하였다. 또한 부푼 비늘눈(bud scale) 사이로 연둣빛 인편이 3mm 정도 밀려나온 눈을 발아한 것으로 간주하였다. 같은 기간 눈의 내부 발달 양상을 알아보기 위하여 먼저 가지 및 액아의 외적 특성을 조사하였다. 가지의 길이는 결과모지로부터 10번째 마디까지의 평균 길이를 산출하였고, 가지의 굵기는 신초의 3번과 4번 마디 사이의 직경을 측정하였으며, 눈의 크기는 액아의 종경과 횡경을 각각 측정하였다. 그 후 액아를 종축으로 절단하여 실체현미경(SZX10, Olympus, Japan) 관찰을 통해 시기별 눈의 생존율과 생존한 눈에서의 화아형성율을 관찰하였으며, 이때 눈의 생존 여부 조사는 Kim et al.(2011)의 방법을 응용하였다.
가용성 당의 분석을 위해 동결 건조한 액아에 80% 에탄올을 가해 중탕으로 가용성 당을 추출하였다. 다시 추출된 당을 원심분리하고 상등액을 0.45μm membrane filter로 여과한 후, sugar-pak I(waters, MA, USA)을 이용하여 물질을 분리하였으며 RI detector를 장착한 HPLC(Agilent 1200 series, USA)로 분석하였다.
통계분석은 PASW Statistics 18(SPSS Inc., USA) 프로그램을 이용하여 수집된 데이터의 Duncan 다중검정(p = 0.05) 및 상관분석을 실시하였다.
결과 및 고찰
발아율 조사를 통한 내재휴면 타파시기 설정
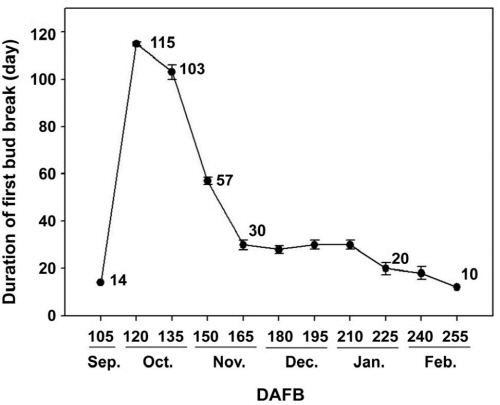
‘Campbell Early’ 품종의 가지를 포장상태에서 채취한 뒤 발아에 적당한 환경 하에 두었을 때, Balandier et al.(1993)의 방법에 따라 chilling unit(CU) 축적이 충분히 이루어져 휴면 타파일에 가까운 것일수록 먼저 발아한다는 가정하에 실험의 결과를 정리하였다. 수삽 후 발아 특성을 조사한 결과, 만개 후 105일에 채취한 가지는 수삽 후 14일부터 액아의 일부가 발아하기 시작하였으나 이내 고사하였다. Lavee and May(1997) 및 Shulman et al.(1983)에 의하면 수삽 후 발아 소요 일수가 약 20-30일을 초과하고, 발아율이 50% 미만일 경우 포도나무의 내재 휴면 기간으로 정의하고 있는데, 만개 후 105일에 채취한 가지는 20일 이내 발아하였으나, 금방 고사하여 발아율이 50% 이상인지 확인하기 어려웠다. 이 시기의 가지는 과실 수확 후 얼마 지나지 않은 시기에 채취된 것으로 내부에 축적된 상대적으로 적은 양의 양분을 발아에 모두 소진하고 고사하는 것으로 추정되어 아직 내재 휴면이 시작되지 않은 것으로 판단되었다.
만개 후 120일에는 최초 발아에 소요되는 기간이 115일로 증가하여 만개 후 105일에서 120일 사이에 내재 휴면이 시작된 것으로 판단되었으며, 만개 후 120일부터 165일까지 채취된 가지의 최초 발아에 소요되는 기간이 115일에서 30일까지 급격히 감소하였고, 만개 후 165일 이후에도 지속적으로 감소하지만 이전에 비해 완만하였다(Fig. 1).
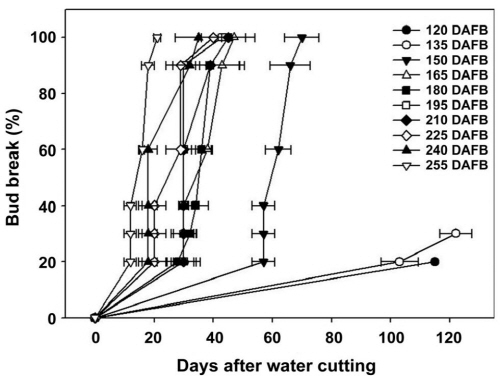
발아율은 만개 후 150일에 채취된 가지부터 시간이 경과됨에 따라 100% 발아하여 내재 휴면이 타파된 것으로 추정 가능하였으나, 만개 후 150일에 채취한 가지는 최초 발아까지의 기간이 57일로 만개 후 165일에 비해 약 2배 정도 길게 소요되어 발아 속도가 현저히 떨어졌다(Fig. 2). 따라서 수삽 후 약 20-30일 정도의 일정 기간 내 발아하고, 발아율이 50% 이상 되는 것을 휴면이 타파된 것으로 보고한 기존의 결과(Lavee and May, 1997; Shulman et al., 1983)와 비교하였을 때, 만개 후 165일 이후에는 100% 발아할 뿐만 아니라 발아 소요 일수가 짧아 내재 휴면이 완전히 타파된 것으로 판단 가능하였다. 위의 결과를 종합하여 최종적으로 ‘Campbell Early’ 포도나무의 내재 휴면은 9월 말에서 11월말까지 약 2달간 지속됨을 알 수 있었다. 또한 만개 후 120일(낙엽기)과 135일에 수집된 가지는 이후 수집된 가지에 비해 발아 소요 일수가 길고 발아율이 낮아 내재 휴면의 심도가 최대에 가까운 것이라 생각할 수 있으며, 이는 상대적으로 축적된 저온의 양이 적기 때문인 것으로 판단되었다(Mohamed et al., 2010). 따라서 발아율은 축적된 저온의 양과 밀접한 관계를 가지며, 어떠한 임계점에 도달한 후에는 큰 차이가 없는 것으로 생각된다.
저온요구도 설정
자연상태에서 휴면의 타파는 유전자에 의해 제어되는 CU 축적을 필요로 하며, 저온이 이를 위한 주요 인자로 알려져 있다(Balandier et al., 1993). 이러한 정도를 매 시간 경과 기온에 따른 가중적 산값으로 표현한 것이 저온요구도로 사실상 내재 휴면 타파에 걸리는 기간을 시간과 기온의 조합인 온도시간으로 표현한 것이라 할 수 있으며, 휴면 타파 시기 예측을 위해 여러 과종에서 이용되어 왔다(Jung et al., 2006). 또한 CU의 축적은 발아 날짜를 계산하고, 화학적인 휴면타파제의 사용을 결정하기 위한 지표로 사용되고 있다(Sagredo et al., 2005; Sabry et al., 2011).
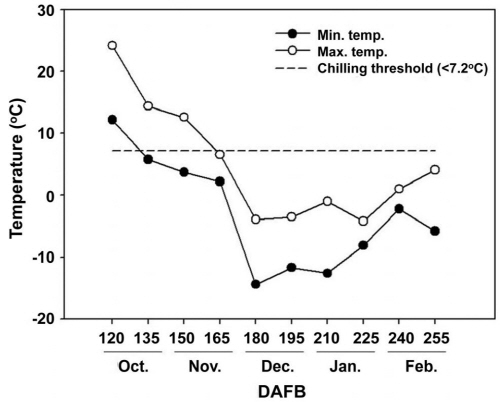
일반적으로 포도나무를 포함한 온대과수의 휴면 타파에 필요한 온도는 7.2oC로 알려져 있어 이를 기준으로 수집된 온도 자료를 분석하였을 때, CU가 처음으로 축적되는 시기는 최저온도가 7.2°C 이하로 내려간 10월 12일로 조사되었다(Fig. 3). 이후 지속적으로 온도가 낮아져 최고온도가 기준 이하로 내려간 시점은 만개 후 165일 즉, 11월 말경이었고, 12월 이후에는 최저, 최고 온도 모두 0°C 이하로 내려가 생리적으로 무효한 온도로 판단하였다.
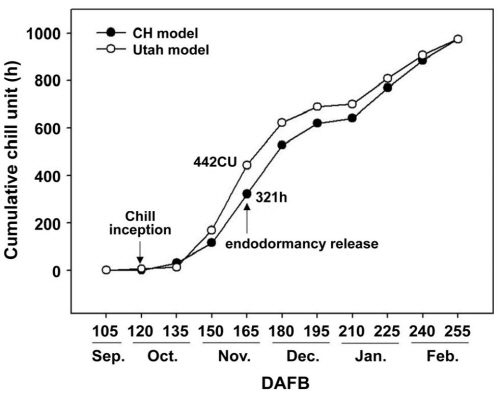
수집된 기온을 0-7.2°C 사이의 온도에서 유지되는 모든 시간을 저온 축적에 유효한 범위로 보며, 기준 범위 안에서 온도에 따른 차이 없이 모두 동일한 효과를 갖고 있음을 고려하는 CH 모델(Chandler, 1942)과 1.4-18.0°C 구간에서 서로 다른 온도 범위에 따라 상이한 저온 축적 효율을 나타내는 가중치를 포함하고 있으며, 저온이 상쇄되는 부정적인 효과도 함께 계산하는 Utah 모델(Richardson et al., 1974)의 두 모델에 적용시켜 요구되는 저온을 계산하였다(Fig. 4). 두 모델 모두에서 10월 초 CU 축적이 시작되었고, 발아 소요 일수 및 발아율 조사에서 내재 휴면이 타파되었다고 추정된 만개 후 165일의 CU은 CH 모델 적용 시 321h, Utah 모델 적용 시 442CU으로 조사되었다. Utah 모델의 경우 1.5-12.4°C 사이의 온도를 CU 축적에 유효한 온도로 판단하고 있어(Rea and Eccel, 2006; Richardson et al., 1974), 본 실험에서도 Utah 모델 이용 시는 1.5-12.4°C 사이의 누적값을 환산 적용하였다. 또한 ‘Perlette’나 ‘Zinfandel’ 품종의 가지는 0-10°C 사이 온도에서 구간별로 저온 처리한 후 발아 조건을 주었을 때, 0-10°C 사이의 구간에서는 최종 발아율에 차이가 없었다는(Dokoozlian, 1999; Weaver and Iwasaki, 1977) 보고 등, 과수의 휴면타파에 7.2°C 이상의 온도도 저온 축적 효과가 있다고 보고되고 있으며, 향후 지구 온난화의 진행은 미래의 겨울철 CU감소를 전제하고 있으므로(Baldocchi and Wong, 2008; Luedeling and Brown, 2011) CH 모델과 Utah 모델을 비교하였을 때, 18.0°C를 기준으로 넓은 온도를 다각적으로 평가한 Utah 모델이 저온요구도 평가에 보다 효과적일 것으로 판단된다.
휴면기의 액아 발달 및 생존율과의 관계
눈의 휴면 유지와 타파는 눈 자체가 가지는 내부 구조 및 물질대사의 변화 수준에서 밀접한 관계를 갖고 있다(Ramina et al., 1995). 또한 눈은 이듬해 결실에 필요한 화원기를 가지고 있으며, 잘 발달된 화원기를 가진 충실한 눈의 확보는 안정된 발아와 고품질의 과실을 생산하는 중요한 요소로 작용한다. 따라서 휴면기와 휴면 타파기 동안의 ‘Campbell Early’ 포도나무의 액아 발달 양상과 생존율을 조사하였다.
9월에 관찰된 눈은 1개의 주아와 2개의 부아를 가지고 있어 포도 액아의 일반적인 형태를 보였으며, 마디, 엽원기(leaf primordium)를 뚜렷하게 구분할 수 있었다. 또한 아직까지 꽃차례(inflorescence)를 보이지 않는 화원기(flower primordium)가 관찰되었다(Fig. 5A). 10월에는 미약하게나마 주아의 꽃차례가 형성되고 있음을 구분할 수 있었으며, 조금 더 두껍고 강하게 발달한 인편이 관찰되었다(Fig. 5B). 또한 모든 시기에서 부아보다 주아의 발달 단계가 더 앞서 있었고, 12월부터 눈의 끝부분에서 인편 밖으로 밀려나온 하얀 솜털(white wool tip)이 관찰되었는데(Fig. 5D, F), 이는 휴면이 끝난 눈이 부풀어 오르고 있다는 사실을 가시적으로 확인할 수 있는 첫 번째 형태학적인 변화로(Andreini et al., 2009), 12월 이후의 눈은 생장 개시를 준비하고 있음을 추정할 수 있었다.
외적 특성 중 시기에 따른 액아의 종경은 10월에 4.84mm에서 이듬해 2월에 5.72mm까지 증가한 반면, 액아의 횡경은 시간이 지날수록 감소하는 경향이었다(Table 1). 10월, 11월 관찰된 주아는 각각 85.4, 91.7%의 높은 생존율을 보이다가 11월 이후 생존율이 급격히 감소하여 2월에는 40%만이 살아있는 것으로 관찰되어(Table 2), 내재 휴면이 타파된 경우 눈이 가지는 내한성이 감소하여 저온에 취약해져 괴사율이 높아질 수 있다는 추정이 가능하였다. 또한 시기가 진전될수록 나타나는 액아의 횡경 감소는 11월 이후 발생하는 주아 괴사에 의한 눈의 외형적 수축현상으로 판단되며, 이러한 괴사와 수반되는 액아의 외형적 수축 현상은 ‘홍이슬’ 및 ‘흑구슬’에서도 이미 보고된 바 있다(Kim et al., 2011; Kwon et al., 2011).
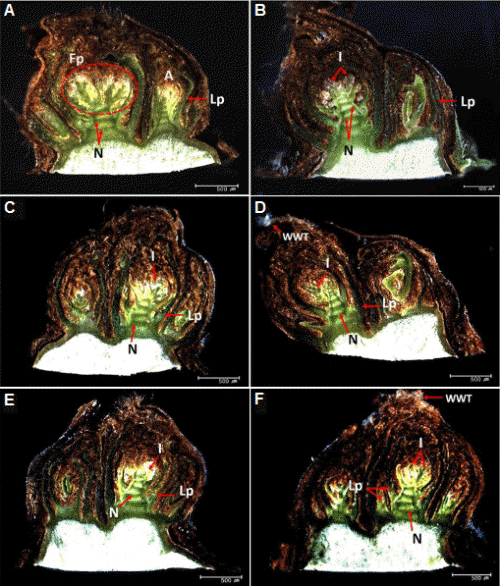
반면에 제 1 부아의 생존율은 시기적인 차이 없이 90% 이상의 수준으로 높게 유지되었다. Choi et al.(2007)의 보고에 의하면 ‘Campbell Early’의 주아 괴사는 만개 후 30일부터 발생하여 50일에서 60일 사이에 급증하고 이후 휴면기까지 완만하게 증가한다고 하였는데, 본 연구에서는 내재 휴면의 타파가 이뤄지는 11월까지도 주아가 건전하게 생존해 있다가 이후 괴사율이 높아지는 것으로 조사되었다. 이러한 결과는 비교적 괴사가 많이 발생하지 않는 직경 8.0mm 이하의 적정 세력의 가지를 수집(Table 1)하여 관찰했기 때문인지(Choi et al., 2007; Kim et al., 2011; Lavee et al., 1981), 또는 생육 초반 이미 괴사한 주아를 대신해 부아가 빠르게 발달하여 휴면기 이후에는 주아처럼 관찰되는 것인지 좀 더 구체적인 연구가 필요할 것으로 생각된다.
살아 있는 주아와 부아를 대상으로 실시한 화아형성율 조사에서도 주아는 생존율 감소와 더불어 화아형성율 역시 감소하였으며, 부아의 경우 11월 15.7%에 불과했던 화아형성율이 2월 조사 시에는 약 3.6배 증가하여 56.7%로 관찰되었다. 화아형성이 이루어진 눈을 대상으로 주아 대비 부아의 비율을 조사한 결과, 내재 휴면이 타파되기 이전인 10월(만개 후 135일)에는 1:0.23으로 주아의 비율이 월등히 높았으나, 내재 휴면이 타파된 11월(만개 후 165일) 이후 꾸준히 증가하여 이듬해 2월(만개 후 255일)에 이르러서는 1:1.54로 주아에 비해 부아의 비율이 높게 나타나 내재 휴면 타파 이후의 꽃눈 형성은 오히려 부아에서 더 충실한 것으로 관찰되었다. 일반적으로 주아가 괴사하거나 세력이 약한 경우, 부아가 대체하여 발달함으로써 1-2개의 화원기를 형성하고, 제 2 부아는 화원기가 거의 없거나 존재하지 않기 때문에(Choi et al., 2007; Lavee et al., 1981; Pratt, 1979), 이듬해 생장하는 신초의 2/3 정도는 제 1 부아가 발달하여 생장한 것으로 추정되었다.
한편, ‘홍이슬’, ‘흑구슬’, ‘Riesling’, ‘Thomson Seedless’ 및 ‘Queen of the vineyard’의 액아는 가지의 세력뿐만 아니라 차광, 가지 내 액아의 위치 또는 수체 내 동화산물의 감소에 의해 눈의 괴사가 발생한다고 하여(Kim et al., 2011; Kwon et al., 2011; Lavee et al., 1981; Vasudevan et al., 1998), 본 실험에서는 가지 채취 시기, 액아의 크기 및 당 함량 중 주아와 부아의 생존율에 어떠한 요소가 영향을 미치는지 각 항목별 상관분석을 실시하였다(Table 3). 그 결과, 부아의 생존율 및 화아형성률은 모든 요소와 상관관계가 없었으며(자료 미제시), 주아만이 위의 요인들과 상관관계를 보이는 것으로 조사되었다. 주아의 생존율은 시기(r = -0.713**), 총 당함량(r = -0.532*)과 음의 상관관계를 나타내었으며 주아의 화아 분화 정도 또한 시기(r = -0.728**), 총 당함량(r = -0.584*)과는 음의 상관관계를 나타내었으나 액아의 직경(r = 0.511*)과는 정의 상관관계를 나타내었다. 그 중에서 특히 주아의 생존율 및 화아형성은 시기와 가장 밀접한 관련을 나타냈다. 또한 여기에서 주목할만한 사실은 주아의 생존율과 화아형성율 사이에 높은 정의 상관관계(r = 0.970**)를 보인 것으로, ‘Queen of the vineyard’에서의 주아 괴사 발생 양상(Lavee et al., 1981)과도 일치하는 결과이다.
따라서 국내에서의 ‘Campbell Early’의 휴면타파 시기는 기존에 알려진 1월 초, 중순보다(Kwon et al., 2005) 한달 이상 빠른 11월 말경으로 조사되었으며, 특히 휴면타파 이후 주아 괴사율이 계속 높아지는 것으로 조사되어 12월 이후의 추위가 이듬해 발아에 크게 영향을 미치는 것으로 판단되었다.
