

Introduction
Materials and Methods
Experimental sites
Plant materials and experimental design
Leaf anatomical observations
Ecophysiological trait measurements
Statistical analysis
Results
Leaf anatomical plasticity across highland and lowland environments
Altitude-induced variations in ecophysiological traits
Trait divergence between highland and lowland conditions
Relationships among apple leaf traits
Cluster-based differentiation of cultivar adaptation strategies
Heatmap and principal component analysis of trait-based adaptation
Discussion
Altitude as a driver of trait shifts
Stomatal and photosynthetic adjustments
Leaf trait responses across altitude gradients
Anatomical trait coordination and multivariate relationships
Conclusion
Introduction
Climate change is increasingly influencing genetic and phenotypic variation, significantly impacting crop physiology, productivity, and sustainability across ecosystems. The link between genetic variation and climate suggests that crops, including apple (Malus domestica Borkh.) have evolved strategies to cope with environmental changes. The trees of this domesticated apple are typically robust and vigorous, having been selected for high fruit production, and they bear fruits that are generally large, variable in shape, smooth-skinned in diverse colors, and palatable for consumption (Ahl et al. 2021). However, as a temperate species native to Europe and Asia, apple is particularly sensitive to temperature and photoperiod changes, typically requiring chilling periods to break dormancy and ensure flowering and fruiting (Chen et al. 2023; Khan et al. 2023).
Despite these constraints, apple cultivation has expanded into tropical highlands, including parts of Indonesia, through adaptive horticultural practices such as branch bending and defoliation to regulate flowering. Indonesia’s apple germplasm includes 72 diverse varieties. Five cultivars—‘Rome Beauty’, ‘Manalagi’, ‘Huanglin’, ‘Royal Red’, and ‘Anna’—have shown promise due to their favorable agronomic and fruit quality traits and adaptability to warmer climates. These cultivars are grown successfully at 750–1700 m in highland regions that simulate temperate conditions through cooler temperatures and higher humidity (Verheij 1990; Anggara et al. 2017; Liu et al. 2020).
Environmental gradients between Indonesia’s highlands and lowlands—differences in temperature, humidity, light intensity, and rainfall—impose selection pressures on plant anatomy and physiology. In highlands, plants face cooler temperatures, lower oxygen levels, stronger UV radiation, and more fluctuations, requiring specialized adaptations for growth and reproduction. Conversely, lowland environments with warmer temperatures and higher humidity challenge plants with heat stress, water scarcity, and disrupted seasonal cues (Fischer et al. 2016; Ferrante and Mariani 2018; Saleem et al. 2025).
Ecophysiological traits provide insights into how plants manage these forms of stress (Bhusal et al. 2018). Anatomical and ecophysiological traits, especially leaf-based characteristics, are crucial for adaptation along altitudinal gradients (Liu et al. 2019; Bai et al. 2019; Liu et al. 2021). Leaves play key roles in photosynthesis, respiration, and transpiration, making them highly sensitive to environmental variations in light, temperature, humidity, soil pH, and moisture (Sahu et al. 2020). Relocated apple plants in lowlands are expected to undergo anatomical changes—e.g., altered leaf thickness, cuticle structure, and stomatal traits (Tian et al. 2016; Liu et al. 2021; Wang et al. 2022), alongside physiological adjustments such as changes in water-use efficiency and photosynthetic rates (Rindyastuti and Hapsari 2016; Hu et al. 2020; Yang et al. 2021). Temperature, in particular, is a dominant factor influencing these leaf traits (He et al. 2018).
Although temperate apple cultivation has been studied extensively, integrative anatomical and ecophysiological analyses under tropical conditions—especially lowland environments—remain limited. To ensure long-term sustainability, ex-situ conservation has duplicated elite varieties at lower altitudes, such as the Purwodadi Botanic Garden (±300 m), which acts as a living laboratory to evaluate adaptive potential under future climate conditions. Therefore, this study investigates varietal-specific anatomical and ecophysiological adaptations of five apple cultivars (‘Rome Beauty’, ‘Manalagi’, ‘Huanglin’, ‘Royal Red’, and ‘Anna’) grown under contrasting tropical highland and lowland settings, marking the first evaluation of temperate apple germplasm in Indonesia’s lowland botanic garden. The findings will provide baseline data for ex-situ germplasm conservation and offer insights to support breeding strategies and sustainable apple cultivation in the face of climate change.
Materials and Methods
Experimental sites
This study was conducted at two contrasting altitudinal sites in East Java, Indonesia. The lowland site was at the Purwodadi Botanic Garden, Pasuruan (±300 m), whereas the highland site was located in Junrejo, Batu (±950 m). The lowland site is characterized by warmer temperatures, lower relative humidity, and greater amounts of annual rainfall, but it experiences relatively low light intensity. In contrast, the highland site has cooler temperatures, higher humidity, and lower rainfall, yet it is exposed to stronger sunlight. The detailed environmental parameters are presented in Table 1.
Table 1.
Climatic and micro-climatic conditions of the study areas
Plant materials and experimental design
Five apple (Malus domestica Borkh.) varieties were selected for the study: ‘Anna’, ‘Huanglin’, ‘Manalagi’, ‘Rome Beauty’, and ‘Royal Red’. Seedlings were initially propagated via budding and grown in polybags at the highland site. After reaching one year of age, the plants were transported to the lowland site for acclimatization. To induce uniform stress adaptation, lowland-grown plants underwent three defoliation cycles before data collection.
For each variety and location, three healthy plants were selected. Fully expanded, mature leaves were sampled to take anatomical and ecophysiological measurements. Each parameter was measured in triplicate.
Leaf anatomical observations
Transverse sections of leaf blades were prepared from regions near the midrib using a hand microtome (MT.5503), following the protocol of Metusala (2017) with modifications. An amount of approximately 1 cm2 of leaf segments was soaked in distilled water and subsequently treated with 5.25% sodium hypochlorite for 3–15 minutes for tissue clarification. Samples were rinsed thoroughly and stored in a 70% ethanol:glycerin solution (3:2). Before being subjected to a microscopic examination, the samples were stained with 1% safranin to enhance the tissue contrast.
To observe the stomata, the abaxial leaf surface was replicated using a clear nail polish technique (Auliya et al. 2019). All anatomical traits were examined under a light microscope under 100x and 400x magnification. Images were captured using an Optilab Advance digital camera and analyzed using ImageJ software.
The quantitative leaf anatomical traits observed were the major leaf structural traits of the leaf area (LA), total leaf thickness (LT), upper/lower epidermis thickness (UE, LE), upper/lower cuticle thickness (UC, LC), palisade mesophyll thickness (P), spongy mesophyll thickness (S), and total mesophyll thickness (M). The tissue proportion indices of the P/M and S/M ratios were observed, as were the vein and stomatal characteristics of the vascular bundle area (VBA), stomatal aperture (SA), stomatal density (SD), and trichome density (TD).
Ecophysiological trait measurements
Ecophysiological traits were assessed using standardized protocols developed by Pérez-Harguindeguy et al. (2013). Ten mature, healthy leaves per replicate (excluding petioles) were collected to determine the following variables: chlorophyll content (CC), specific leaf area (SLA), specific leaf weight (SLW), and leaf dry matter content (LDMC). CC was measured using a CCM-200 Plus Chlorophyll Content Meter, SLA was calculated as the ratio of leaf area to the leaf dry weight (cm2/g), SLW was obtained by dividing leaf dry weight by leaf area (mg/cm2), and LDMC was determined as the ratio of the leaf dry weight to the fresh weight (mg/g).
Fresh leaf area was measured by photographing leaves against a black cloth background with a ruler for scale and analyzing the image in ImageJ. Leaves were oven-dried at 80°C for 48 hours to determine the dry weight.
Statistical analysis
Quantitative data for anatomical and ecophysiological traits were analyzed by means of a one-way ANOVA to assess differences among varieties at each altitude. Mean separation was conducted using Duncan’s multiple range test at α = 0.05. To evaluate the effect of altitude on each variety, independent t-tests were applied (α = 0.05). All statistical tests were conducted using SPSS 23.0.
To explore multivariate relationships and trait contributions to varietal differentiation across altitudes, a principal component analysis (PCA) was performed using a correlation matrix in the PAST 4.03 software package. A biplot was generated to visualize trait clustering and altitude-based separation. Heatmaps and hierarchical clustering were produced in R 4.3.2 using the pheatmap package after standardizing variables via Z-scores.
Results
Leaf anatomical plasticity across highland and lowland environments
Significant variations in leaf anatomical traits were observed among the five apple varieties and between the two altitudinal environments (Figs. 1, 2, 3). Across all cultivars, LA and LT were consistently and significantly reduced in the lowland environment. ‘Manalagi’ and ‘Anna’ exhibited the most significant LA values in the highland area, while ‘Royal Red’ showed the lowest values. In contrast, ‘Royal Red’ and ‘Anna’ developed the thickest leaves, with ‘Huanglin’ and ‘Manalagi’ displaying the thinnest lamina. When transferred to the lowland, ‘Rome Beauty’ experienced the most significant reduction in LA (53.91%), while ‘Royal Red’ showed the least reduction (16.89%). For LT, ‘Royal Red’ also exhibited the highest decrease (45.63%), while ‘Anna’ showed the smallest (25.02%) (Fig. 1A and 1B).
Notably, specific protective structures increased in thickness under the lowland conditions. UE thickness significantly increased in ‘Anna’ (40.94%) and ‘Manalagi’ (42.57%). Similarly, UC thickness generally increased across the varieties tested here, although the differences were not statistically significant. In contrast, LE and LC thickness outcomes were significantly reduced in most varieties under lowland stress. ‘Anna’ consistently developed the thickest UC and UE layers across environments (Figs. 1C, 1F, and 2).
P, S, and total M were significantly thinner in all varieties at the lowland site (Figs. 1G–1I, and 2). ‘Royal Red’ developed the thickest palisade layer at the highland site, which decreased by 41.59% in the lowland, while ‘Manalagi’ experienced the most pronounced reductions in S and M (55.01% and 49.73%, respectively). Tissue proportion ratios (P/M and S/M) showed variable responses. While most differences were not statistically significant, ‘Royal Red’ exhibited a significant 30.82% reduction in P/M and a 36.62% significant increase in S/M in the lowland environment (Fig. 1J–1K).
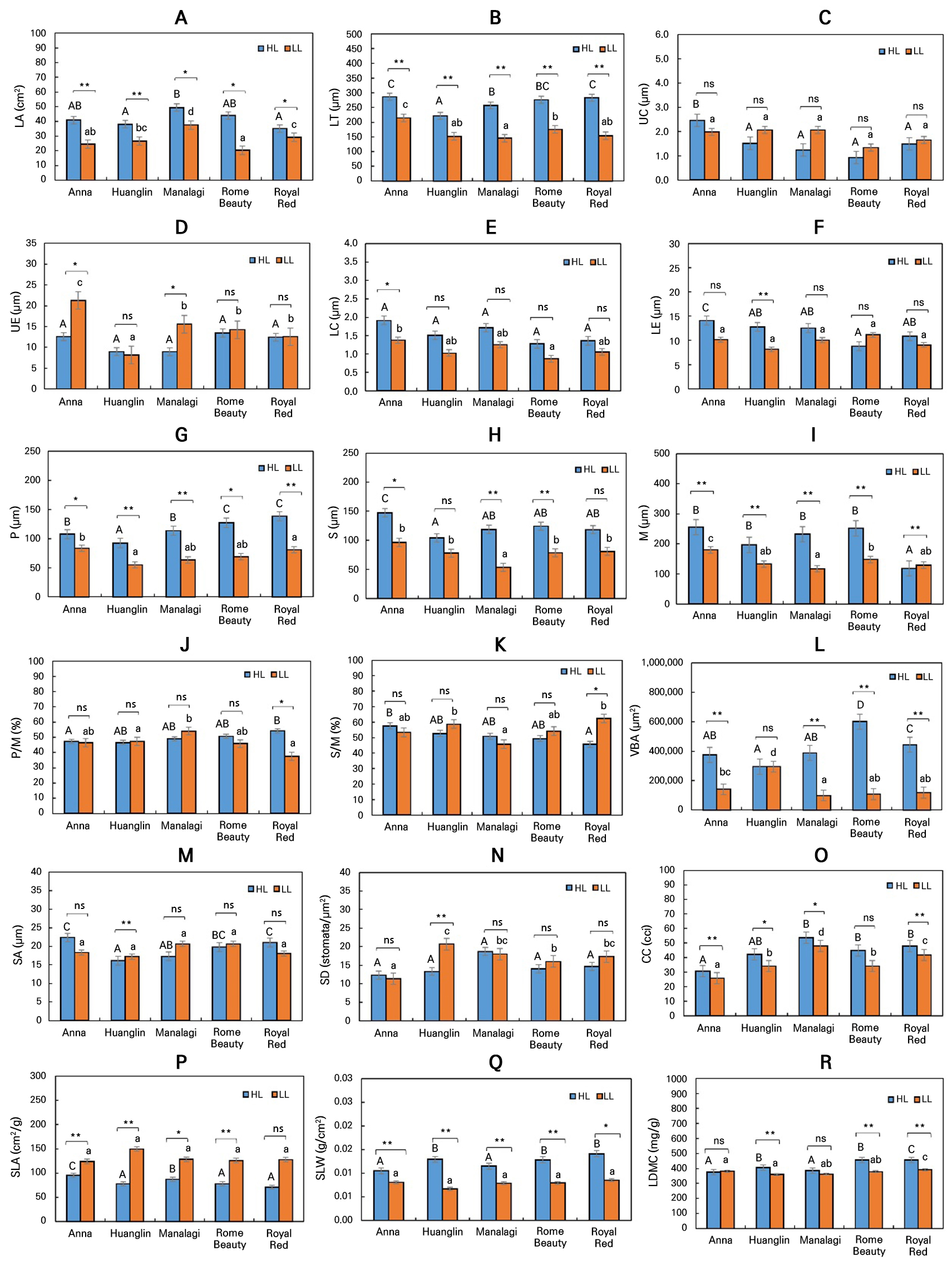
Fig. 1.
Comparative leaf anatomical and ecophysiological traits of five apple varieties under tropical highland (HL) and lowland (LL) conditions.
Note: Measured traits include those that are anatomical (A–N) and ecophysiological (O–R). Bars represent the mean ± SE (n = 3). Different capital letters indicate significant differences among varieties within the highland group; lowercase letters denote significant differences within the lowland group (ANOVA, Duncan’s test, p < 0.05). Asterisks (*) indicate significant differences between highland and lowland conditions within each variety (T-test); ns = not significant.
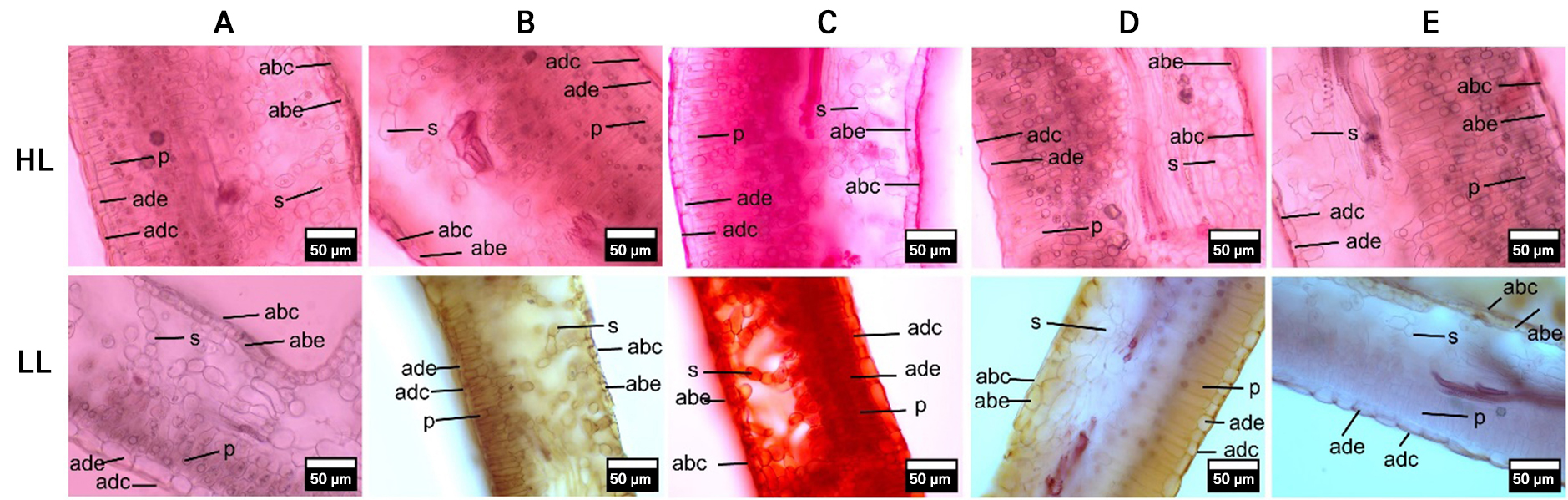
Fig. 2.
Transverse anatomical sections of the leaf blade in five apple varieties under highland (HL) and lowland (LL) conditions.
Note: Representative images illustrate changes in tissue structure across altitudes, in this case adaxial cuticle (adc), adaxial epidermis (ade), palisade (p), spongy mesophyll (s), abaxial epidermis (abe), and abaxial cuticle (abc). Varieties: A = ‘Anna’, B = ‘Huanglin’, C = ‘Manalagi’, D = ‘Rome Beauty’, E = ‘Royal Red’. Magnification: 400x.
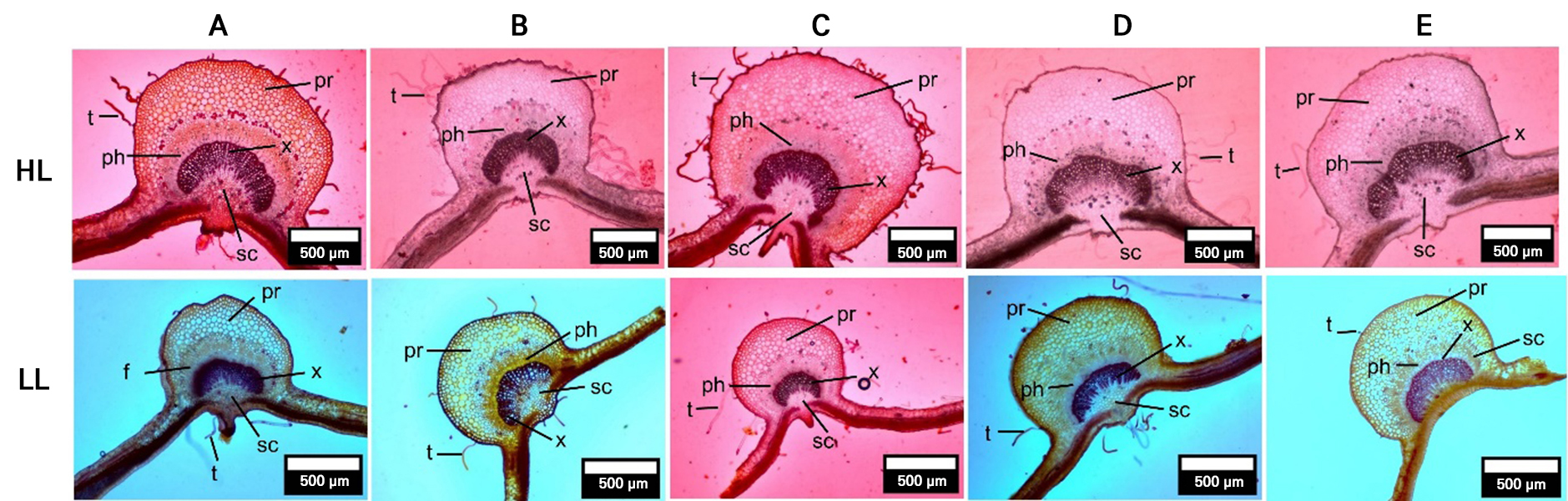
Fig. 3.
Transverse sections of leaf midrib vasculature across five apple varieties grown in highland (HL) and lowland (LL) conditions.
Note: Visual comparisons highlight structural changes in vascular bundle tissues (xylem [x], phloem [ph]), parenchyma (pr), sclerenchyma (sc), and trichomes (t). Anatomical variation suggests that lowland-grown plants have reduced vascular investment. Varieties: A = ‘Anna’, B = ‘Huanglin’, C = ‘Manalagi’, D = ‘Rome Beauty’, E = ‘Royal Red’. Magnification: 100x.
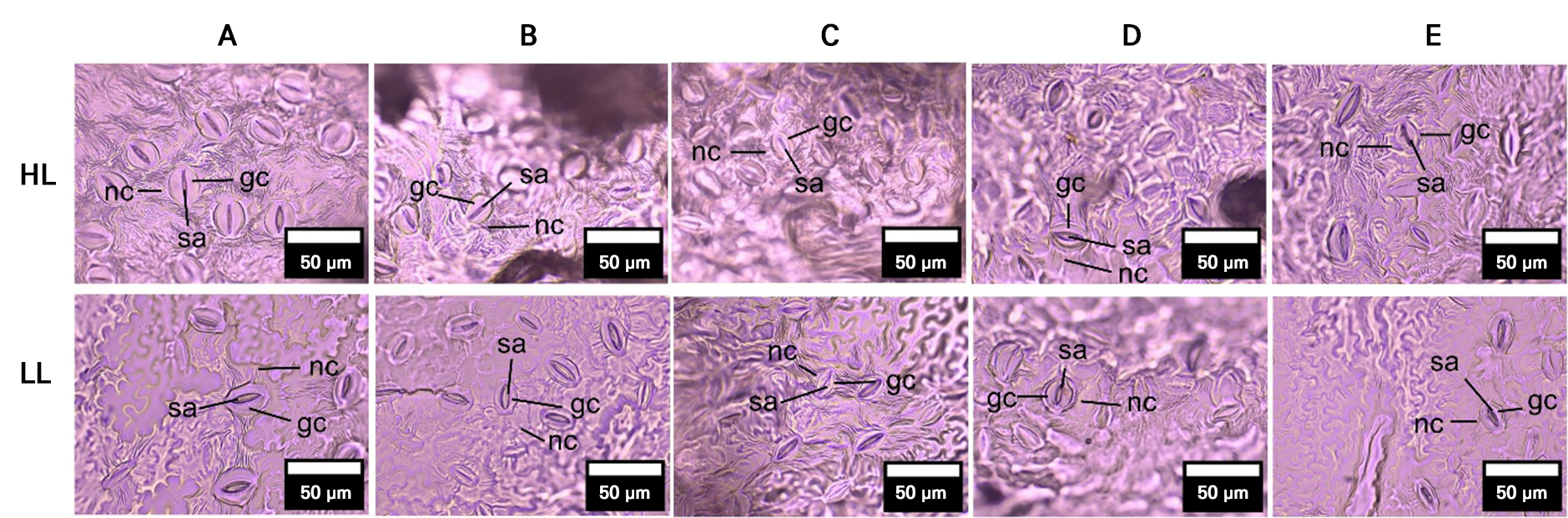
Fig. 4.
Stomatal impressions from the abaxial leaf surface of apple varieties under highland (HL) and lowland (LL) conditions.
Note: Images show variation in stomatal aperture (sa), guard cells (gc), and neighboring cells (nc) among cultivars. Differences in stomatal traits suggest cultivar-specific adjustments to environmental conditions. Varieties: A = ‘Anna’, B = ‘Huanglin’, C = ‘Manalagi’, D = ‘Rome Beauty’, E = ‘Royal Red’. Magnification: 400x.
VBA also declined significantly in most cultivars under lowland conditions (Figs. 1 and 3). ‘Rome Beauty’ had the largest VBA in the highland and the most significant reduction at the lowland site (82.14%), followed by ‘Royal Red’, ‘Manalagi’, and ‘Anna’. Only ‘Huanglin’ maintained relatively stable VBA values, showing the smallest decrease (42.40%).
Stomatal characteristics showed cultivar-specific responses to altitude (Figs. 1M, 1N, and 4). A significant increase in both SA and density SD was observed in ‘Huanglin’ under lowland conditions. Other cultivars exhibited varying but non-significant trends, with SA decreasing in ‘Anna’ and ‘Royal Red’ but increasing in ‘Manalagi’ and ‘Rome Beauty’, and SD increasing in ‘Rome Beauty’ and ‘Royal Red’ but decreasing in ‘Anna’ and ‘Manalagi’.
Altitude-induced variations in ecophysiological traits
CC declined significantly in all varieties under lowland conditions, with the reductions ranging from 10.60% to 23.96%, except in ‘Rome Beauty’, which showed no significant change. ‘Manalagi’ maintained the highest CC overall, followed by ‘Rome Beauty’, ‘Royal Red’, ‘Huanglin’, and ‘Anna’. In contrast, SLA increased significantly for most varieties at the lowland site, indicating the development of thinner and broader leaves. The most pronounced SLA increase was observed in ‘Huanglin’ (48.02%), while ‘Anna’ exhibited the smallest increase (23.12%) (Figs. 1O–1R).
SLW significantly decreased across all cultivars under lowland conditions, consistent with the SLA trends. Varieties with inherently higher SLW values at the highland site (‘Huanglin’, ‘Rome Beauty’, ‘Royal Red’) experienced a notable reduction, suggesting reallocation away from dense tissue formation under heat or water stress. ‘Anna’ and ‘Manalagi’ consistently displayed lower SLW values compared to the other varieties, even at the highland site.
LDMC also declined significantly under lowland conditions in most varieties, suggesting lower tissue density and possibly reduced resource investment in the leaf structural components. ‘Royal Red’ consistently maintained the highest LDMC, whereas ‘Anna’ exhibited the lowest. Interestingly, LDMC outcomes in ‘Anna’ and ‘Manalagi’ did not show significant differences across altitudes.
Trait divergence between highland and lowland conditions
Altitude had a significant effect on most leaf anatomical and ecophysiological traits in the five apple varieties studied here, as confirmed by independent t-tests (Table 2). Among the anatomical traits, LA and total LT were significantly lower in lowland-grown plants (p < 0.01). Other laminar traits, specifically LC, LE, P, S, M, and VBA, were also significantly reduced. In contrast, UE thickness increased significantly (p = 0.042), while UC showed a non-significant increasing trend.
Table 2.
Leaf anatomical and ecophysiological responses of apple varieties under tropical highland and lowland conditions
SA and SD showed no significant difference across altitudes. However, SD tended to be higher, and SA slightly lower, in the lowland samples, suggesting potential shifts in stomatal behavior aimed at regulating gas exchange and minimizing water loss. However, these trends were not statistically significant. With regard to ecophysiological traits, all four measured variables differed significantly between the highland and lowland environments. CC, SLW, and LDMC were all significantly lower in the lowland environment (p < 0.01).
Relationships among apple leaf traits
A comprehensive correlation among apple leaf traits was observed across environments and varieties (Fig. 5A). In particular, mesophyll thickness was strongly correlated with both palisade (r = 0.85) and spongy parenchyma (r = 0.88) layers. Moreover, the thickness of the cuticle and upper epidermis exhibited a strong positive association (r = 0.65). Interestingly, a lower epidermal thickness displayed a moderately negative correlation with stomatal aperture (r = –0.55), while SD was weakly correlated with epidermal or mesophyll traits. SLA displayed a strong negative correlation with mesophyll thickness (r = –0.62; R2 = 0.39) and a moderate negative correlation with the chlorophyll content (r = –0.57; R2 = 0.34). In contrast, the negative correlation between stomatal density and aperture size was weak (R2 = 0.12).
Cluster-based differentiation of cultivar adaptation strategies
To classify cultivar-specific adaptation mechanisms further, hierarchical clustering was utilized using all measured anatomical and ecophysiological traits. The resulting dendrogram (Fig. 5B ) groups the five apple cultivars into two major clusters, revealing clear multivariate differentiation in their trait syndromes. The first cluster consisted primarily of ‘Royal Red’, ‘Huanglin’, and ‘Rome Beauty’, which exhibited high SLA values, a thin cuticle and epidermis, and a thinner mesophyll thickness. The second cluster included ‘Manalagi’ and ‘Anna’, which displayed structurally robust traits such as a thicker cuticle, mesophyll, and epidermis, along with reduced SLA values.
Heatmap and principal component analysis of trait-based adaptation
The findings of the heatmap analysis (Fig. 5C) illustrate cultivar-specific anatomical and ecophysiological syndromes by visualizing standardized (Z-score) values for all measured traits. Notably, ‘Royal Red’ and ‘Rome Beauty’ exhibited higher SLA values, a greater stomatal aperture, and thinner mesophyll layers. In contrast, ‘Manalagi’ displayed high positive values for mesophyll thickness, the epidermal layers, and the chlorophyll content. ‘Anna’ also showed elevated investment in the upper epidermis and cuticle thickness, while ‘Huanglin’ emerged as an intermediate cultivar, balancing moderate SLA values and mesophyll development with a relatively high stomatal density. The heatmap thus provides a clear visual summary of adaptive trait expression, supporting multivariate selection for cultivar suitability across diverse tropical agroclimates.
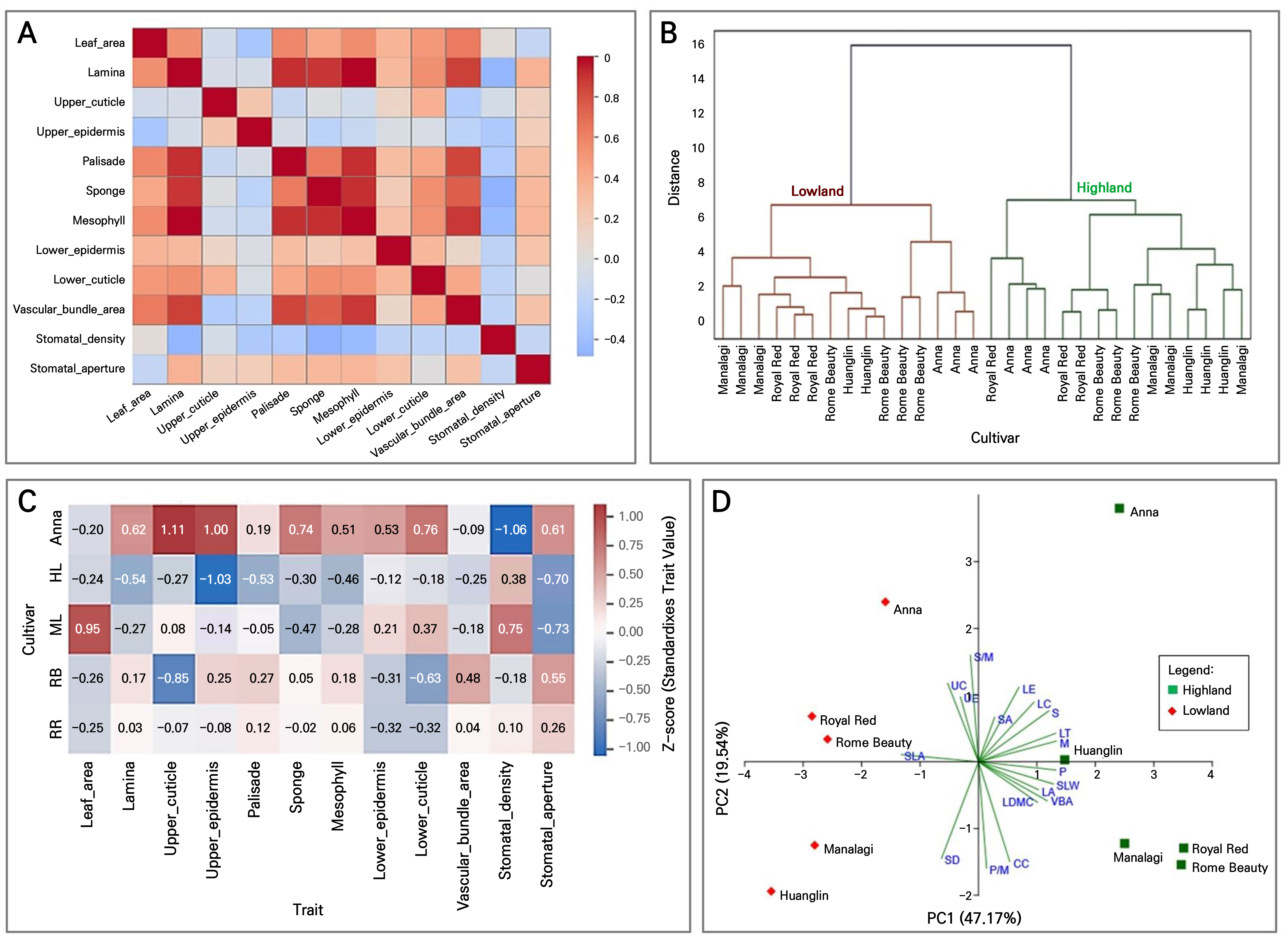
Fig. 5.
Correlogram illustrating Pearson correlations among anatomical traits in apple leaves across all cultivars and environments (A), and dendrogram showing the hierarchical clustering of apple cultivars based on standardized anatomical and ecophysiological traits (B), Heatmap showing standardized (Z-score) trait values across apple cultivars based on anatomical traits (C), and PCA biplot (D). Blue and red tones reflect low and high relative investment, respectively. Color intensity reflects the strength and direction of the correlation: red indicates a positive correlation, and blue indicates a negative correlation.
The PCA biplot (Fig. 5D) revealed distinct clustering of samples according to altitude, confirming that environmental differences between the highland and lowland sites strongly influence the expression patterns of that trait. The first two principal components (PC1 and PC2) accounted for a combined 66.71% of the total variance, with eigenvalues of 8.49 and 3.52, respectively. PC1 primarily captured variations in the leaf size, thickness, and dry matter investment traits, while PC2 was more associated with tissue proportions and stomatal behavior. Samples from the lowland environment clustered toward the positive side of PC1, strongly associated with increased SLA, UC and UE thickness, S/M, and SD outcomes. In contrast, samples from the highland environment clustered on the negative side of PC1, characterized by LT, MV, VBA, and CC traits.
Discussion
Altitude as a driver of trait shifts
Under lowland conditions, most apple varieties showed reductions in the total leaf thickness (LT), mesophyll tissues (P, S, M), and vascular bundle area (VBA), indicating a shift in resource allocation to reduce water loss (Guo et al. 2017; Saputra et al. 2018; Bernado et al. 2021; Wang et al. 2022; Khan et al. 2023). ‘Royal Red’ had the smallest reductions in LA and LT, suggesting better heat tolerance, while ‘Rome Beauty’ showed the most significant declines. The ability to maintain leaf size and thickness suggests greater structural stability and resilience to heat stress in the lowland conditions. Notably, under high-altitude conditions, thicker leaves help maintain the internal temperature and reduce UV damage (Xinrui et al. 2023).
Interestingly, UC and UE thickness measures were found to increase in some lowland samples, particularly in ‘Anna’ and ‘Manalagi’, likely as protective adaptations to reduce transpiration and solar stress (Yang et al. 2021; Ke et al. 2022). ‘Anna’ had the thickest UC and UE, indicating its resilience in warmer climates. Significant thickening of the upper and lower epidermis in the ‘Anna’ variety serves as a protective layer that suppresses water loss through the cuticle while also retaining thermal radiation. Physiologically, this mechanism protects the mesophyll tissue from oxidative stress due to extreme temperatures, but it prolongs the CO2 diffusion pathway, thereby reducing the net photosynthesis potential. Thus, this protective strategy illustrates the trade-off between increased heat resistance and decreased gas exchange efficiency (Harrison et al. 2021; Muñoz-Gálvez et al. 2025).
Conversely, the ‘Royal Red’, ‘Rome Beauty’, and ‘Huanglin’ varieties showed increased SLA and SLW outcomes (approximately 1.6 fold), indicating higher photosynthetic efficiency per unit biomass and improved structural resistance of the leaves. Leaves with high SLA values support intensive photosynthetic capacity but are more vulnerable to overheating and mechanical damage (Jovanović et al. 2023; Muñoz-Gálvez et al. 2025). This emphasizes the presence of an ecological trade-off, where thin-leaved/active metabolic varieties excel in stable agronomic conditions but are less resistant to extreme heat fluctuations.
This trade-off is very important to understand in the context of variety adaptation to heat stress and the development of superior breeding. Conservative strategies such as that displayed by the ‘Anna' variety inspire the breeding of heat-tolerant varieties through structural protection, while efficiency strategies such as those of ‘Royal Red’ and its peers can be developed under relatively more stable agronomic conditions. Further research investigating the relationships among the epidermal structure and SLA/SLW with the photosynthesis rate, water loss, and internal leaf temperature—as well as their integration into hydraulic trade-off models—will strengthen our understanding of cost-benefit trade-off, ultimately enabling the development of superior and climate-adaptive varieties.
Stomatal and photosynthetic adjustments
The lack of significant differences in stomatal traits across environments suggests that apple varieties rely more on anatomical and SLA adjustments than stomatal changes to manage stress. Cultivar-specific patterns emerged: ‘Huanglin’, ‘Rome Beauty’, and ‘Royal Red’ increased SD in lowlands to maintain CO2 uptake, while ‘Anna’ and ‘Royal Red’ reduced SA for water conservation (Jane et al. 2016; Huang et al. 2022; Haworth et al. 2023; Lee et al. 2023). A significant decline in CC under lowland conditions suggests possible heat-induced chloroplast damage (Metusala 2017; Hu et al. 2020; Liu et al. 2020).
Leaf trait responses across altitude gradients
The t-test highlights key adaptive responses of apple plants to highland and lowland environments. In lowlands, they develop smaller, thinner leaves to reduce water loss while increasing LA to optimize light capture. Altered stomatal traits help balance gas exchange and water use. Reduced CC, SLW, and LDMC, alongside higher SLA outcomes, suggest a shift toward maximizing photosynthesis with minimal water loss. These adaptations reflect a trade-off between growth and conservation (Dwyer et al. 2014; Jo et al. 2024), essential for climate-resilient apple cultivation.
Anatomical trait coordination and multivariate relationships
The correlogram and trait-level regressions reveal distinct multivariate strategies among apple cultivars, balancing structural investment, gas exchange, and photosynthetic capacity. These integrated trait patterns across environments support trait-based selection for tropical adaptation under climate stress (Xin’e et al. 2022; Liying et al. 2023; Hai et al. 2023). Clear cluster separation confirms tight coordination of anatomical and functional traits, reinforcing adaptation syndromes. The PCA biplot analysis showed that lowland samples exhibited favorable values for SLA, UC, UE, SD, and S/M—traits that enhance light capture, gas exchange, and heat regulation. Highland samples clustered with LT, M, VBA, and CC in the PCA biplot, reflecting traits associated with biomass retention and photosynthetic resilience. Each cultivar showed unique adaptations: ‘Anna’ emphasized protective traits (thicker cuticle, epidermis) for water loss control; ‘Huanglin’ and ‘Manalagi’ modified stomatal and mesophyll traits for improved gas exchange; ‘Rome Beauty’ and ‘Royal Red’ increased SLA for higher carbon gain but potentially lower water-use efficiency. These genotype-dependent strategies mirror findings in other perennial crops (Tian et al. 2016; Bai et al. 2019; Yang et al. 2021; Rayyan et al. 2023). Cultivar-specific traits offer critical guidance for climate-resilient breeding—e.g., ‘Anna’ for dry zones, ‘Rome Beauty’ for shaded lowlands. Understanding temperate apple adaptation to tropical lowlands is crucial amid shifting climate zones (Lawson and Blatt 2014; Sahu et al. 2020; Erqian et al. 2020; Xuying et al. 2023; Saleem et al. 2025). These insights inform trait-based selection for ex-situ conservation and the development of resilient cultivars for tropical orchards.
Following the findings of this study, future research should investigate in more depth the eco-physiological trade-offs underlying cultivar-specific adaptive strategies across a broader range of apple genotypes. The integration of anatomical and physiological assessments with transcriptomic and metabolomic approaches would provide deeper insights into the regulatory networks underlying these traits. Moreover, long-term field evaluations under projected climate scenarios are required to assess the stability of adaptive responses and to identify cultivars capable of consistent performance. Such studies will strengthen trait-based screening for cultivar selection and support breeding programs aimed at developing climate-resilient apple cultivars suitable for ex situ conservation and lowland orchard systems.
Conclusion
Cultivar-specific responses revealed distinct adaptive syndromes: ‘Anna’ favored structural protection through thicker a cuticle and epidermis, ‘Huanglin’ and ‘Manalagi’ adjusted stomatal traits to enhance gas exchange, while ‘Rome Beauty’ and ‘Royal Red’ exhibited increased SLA and photosynthetic plasticity. These findings highlight the importance of trait-based screening for selecting apple cultivars suitable for non-traditional or climate-challenged environments. For germplasm conservation and management in tropical lowland botanical gardens, these findings underscore the importance of selecting varieties with high SLA levels, moderate chlorophyll contents, and anatomical flexibility to ensure optimal physiological performance in warmer climates. Additionally, these traits could serve as selection criteria in breeding programs targeting climate-resilient apple cultivars suitable for future ex situ and lowland orchard systems.
