서 언
재료 및 방법
실험 재료
춘화처리, 재배 온도 관리 및 토양수분 결핍 처리 방법
엽육조직의 형태적 관찰
생장 해석 및 수량 조사
실험설계 및 통계방법
결과 및 고찰
배추의 엽육조직
배추의 생장
배추의 생장해석
배추의 수량 및 결구 형성
배추의 생장
서 언
배추는 최근에 시설재배가 시도되고 있으나 주로 노지에서 95%가 생산되기에 기상조건의 영향을 많이 받는 전형적인 밭작물이다. 호냉성 채소인 배추는 생육적온 보다 높은 온도에서는 생육저하 및 세포가 손상되어 품질저하와 생산량이 감소 된다(Lee et al., 2009; Lee et al., 2011; Lee et al., 2012; Oh et al., 2014). 여름철 배추는 높은 온도와 습도로 인한 생리장해 및 병해충 발생으로 평지에서 생산이 어려워서 주로 고랭지 지역에서 집중적으로 생산되기 때문에 그 지역의 가뭄이나 집중 호우시 피해에 따라 가을배추의 가격변화에 미치는 영향이 크다(Hwang et al., 2003).
배추를 저온기에 육묘할때 온도관리가 부적절하게 되면 화아분화와 추대를 유발하여 상품성이 저하되며, 여름철 고온은 저온성 작물인 배추의 생리장해를 유발하여 생장과 결구형성 등에 부정적인 영향을 미친다. 장마가 장기간 동안 지속되는 해에는 일조부족으로 배추의 생육을 떨어뜨리고 무름병 및 뿌리혹병 등의 발생을 증가시켜 생산량 감소의 원인이 된다. 반대로 비가 오지않는 해에는 토양 함수량이 낮아 배추 체내의 수분 부족현상으로 위조증상을 초래하고 신엽 발생의 억제 및 tip burn 발생으로 수량이 감소하게 된다. 이러한 생리적 장해 요소들에 대한 많은 연구가 보고되었는데, 배추 추대 특성에 대한 유전적 분석(Kakizaki et al., 2011)과 추대 사이의 여러 생리적 요소들의 관계를 구명하기 위한 연구(Li et al., 2010)가 수행되었다. 그리고 온도와 일장이 봄배추 추대에 미치는 영향(Lee et al., 2011)을 비롯해 봄배추의 추대를 억제하고자 triazole계 화합물 처리(Seong et al., 2003)를 통해 추대 억제에 관한 연구들이 보고되었다. 고온 스트레스에서 배추의 세포 손상과 사멸 가능성(Oh et al., 2014), 생육저하와 결구불량(Lee et al., 2009), heat shock protein의 발현과 석회 흡수 패턴(Hwang et al., 2003), 그리고 갓을 40°C 이상으로 고온처리하였을 때 생육과 엽록소 함량이 20%이상 감소한다는 보고(Hayat et al., 2009) 등 고온에 의한 엽채류 작물의 생리적 및 생화학적 반응에 미치는 영향이 연구되었다. 가뭄에 대한 연구들로는 건조 스트레스시 양배추의 생리적 변화와 내건성(Hajiboland and Amirazad, 2010) 및 유채의 건조에 의한 피해현상(Hashem et al., 1998)이 보고 되었다. 그리고, Xu and Zhou(2005)는 온도와 건조 스트레스 조합 처리가 잔디의 광합성과 질소 대사에 미치는 영향과 지질 과산화(Xu and Zhou, 2006)에 미치는 영향을 보고하였다.
배추 재배시 저온에 감응하여 나타나는 생리장해인 추대, 그리고 고온, 가뭄에 대한 다양한 연구들이 보고되었으나 이들이 결합된 작용으로 인한 생리적 반응이나 장해에 대한 연구는 부족한 실정이며 특히 최근의 지구온난화로 인한 빈번한 이상기상에 대비하기 위해 배추의 생장에 영향을 미치는 주요 생리장해에 대한 연구를 통한 피해평가와 대응기술개발이 절실히 요구된다(Lee et al., 2009). 따라서 본 실험은 여름철 배추재배시 안정적인 생산을 위하여 초기 저온(춘화처리)과 재배 기간의 온도관리(적온과 고온), 토양 수분 결핍 처리(단수 기간)가 배추의 엽육조직, 생장 및 발달 그리고 생산량에 미치는 영향을 구명하고자 실시하였다.
재료 및 방법
실험 재료
배추의 공시 품종은 ‘춘광’(Sakada Korea Ltd., Seoul, Korea)이었다. 2014년 3월 10일 국립원예특작과학원 실험 온실(경기도 수원)에 시판용 경량상토인 바이오상토1호(Hungnong Seed Co., Suwon, Korea)를 105공 플러그 트레이(35mL/공)에 채우고 1셀당 2립씩 파종하였고 발아 후에는 1주씩 남기고 솎아 주었다. 정식은 파종 23일 후 재식간격 70 × 45cm을 기준으로 하여 직경이 45cm이고 깊이가 60cm인 플라스틱 포트에 정식하였다.
춘화처리, 재배 온도 관리 및 토양수분 결핍 처리 방법
정식 활착 후 7일간 10°C로 춘화처리를 한 배추와 춘화처리를 하지 않은 배추로 구분하였고, 시설내 일중 최고 온도를 정상적 온도 관리구(25°C)와 고온 관리구(30°C)를 두었다. 그리고 정식 활착후 8일부터 관수를 중단하여 토양 건조 처리를 각각 2주간(토양수분 평균함량 20% 유지)와 4주간(토양수분 평균함량 10% 유지) 한 것과 정상적으로 충분히 관수한 대조구(토양수분 평균함량 40% 유지)를 두었다. 실험에 대한 처리구들을 표로 정리하였다(Table 1).
엽육조직의 형태적 관찰
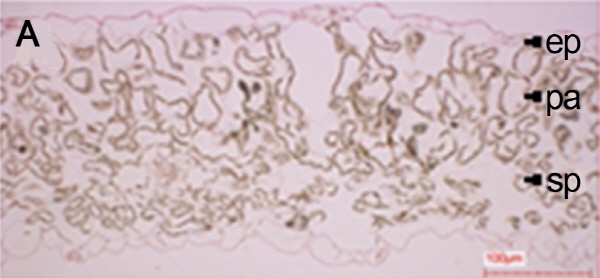
배추의 엽육조직 관찰은 춘화처리를 하지 않고 정상적 온도 관리를 하고 충분히 관수한 처리구(NV25FI), 2주간 토양수분 결핍 처리구(NV25SD2) 및 4주간 토양수분 결핍 처리구(NV25SD4)의 3-5번째 외엽을 채취하여 1차 고정액 2.5% glutarldehyde에서 90분간 고정을 위한 처리, 0.1M phosphate buffer(pH 7.2)로 15분 간격 4-5회 세척, 2차 고정 1% osmium tetroxide 90분간 처리, 위와 동일한 세척 과정 후 하룻밤을 침지시켰다. 탈수는 상온에서 40, 60, 80, 90, 95% ethanol로 각각 5분씩, 100% ethanol로 5, 10, 15, 30분간 처리로 이루어졌으며 propylene oxide로 치환 후 최종적으로 epon에 포매(embedding)하여 60°C의 항온기에서 4일간 중합시켰다. 중합된 epon block을 초미세절편기(Ultracut R, Leica Co., North Ryde, Australia)를 이용하여 1,500nm의 두께로 시료를 절단하여 P.A.S. 염색법으로 염색한 후 광학현미경(Axioskop 2, Carl Zeiss Co., Oberkochen, Germany) 20μm로 검경하였다(Chang, 1973; Dorange and Pennec, 1989).
생장 해석 및 수량 조사
정식후 60일에 처리구의 반복별로 3주씩 최대 엽장, 엽수, 엽면적(LI-3100, Area meter, LI-COR Inc., Nebraska, USA), 생체중 및 건물중을 조사하였다. 배추의 생장해석은 상대생장률(relative growth rate; RGR), 단위 엽건물중 증가율(unit leaf rate; ULR), 엽면적비율(leaf area ratio; LAR) 및 비엽중(specific leaf area; SLA), 엽중률(leaf weight ratio; LWR)로 하였다. 생장해석의 요소(RGR과 ULR)들은 아래와 같이 계산되었고, 정식일의 엽면적(73.15cm2/plant), 엽중(3.26g/plant), 건물중(0.27g/plant) 값을 초기값으로 넣어 계산하였다. 그리고 LAR은 RGR에 ULR로 나눈 값으로 하였고, SLA는 엽면적을 엽중으로 나눈 값으로 하였으며, LWR은 엽중을 총건물중으로 나눈 값으로 하였다.
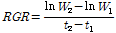
W1: t1 시기(정식일)에서의 작물의 건물량, W2: t2 시기(정식 후 60일)에서의 작물의 건물량.
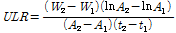
W1: t1 시기(정식일)에서의 엽면적의 건물중, W2: t2 시기(정식 후 60일)에서의 엽면적의 건물중, A1: t1 시기(정식일)에서의 단위 엽면적, A2: t2 시기(정식 후 60일)에서의 단위 엽면적.
정식후 60일에 수확한 배추의 지상부 생체중을 처리구 당 10주씩 조사하여, 평균 생체중에 재식밀도(3,000주/10a)를 곱하여 수량으로 산출하였다. 배추의 결구력은 결구 형성 및 단단한 정도를 조사하여 지수화(상, 중, 하) 하였다.
실험설계 및 통계방법
실험 처리구는 완전임의배치 3반복으로 하였다. 춘화처리, 재배 환경 온도처리 그리고 토양수분 결핍 처리의 3요인 분산분석을 하였다. 각 처리구별 생장 요인들과 수량에 대해 Duncan의 다중 검정을 실시하였다. 통계처리는 SAS 프로그램(SAS 9.2, SAS Institute Inc., NC, USA)을 이용하였다.
결과 및 고찰
배추의 엽육조직
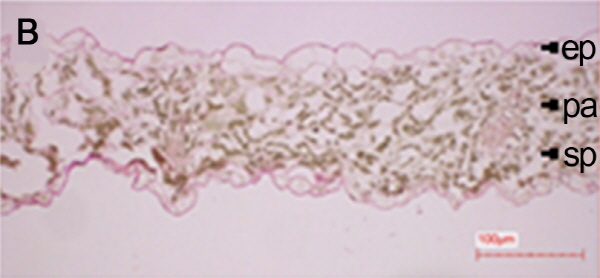
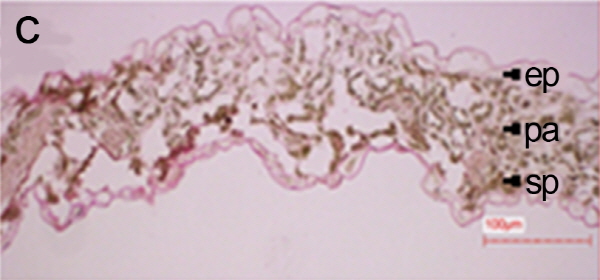
처리별 엽육조직을 광학현미경으로 관찰한 결과, 2주 토양수분 결핍 처리구(NV25SD2)에서는 울타리조직과 해면조직의 세포구조를 확인할 수 있었으나 4주 토양수분 결핍 처리구(NV25SD4)에서는 세포조직이 완전히 붕괴되었다(Fig. 1). 본 연구결과와 유사하게 건조 스트레스에 의해 시금치 잎 조직내의 전분이 소실되고 thylakoid에 대한 stroma의 부피비 증가로 인하여 잎의 세포형태가 변화되는 것으로 보고 되었다(Zellnig et al., 2004). 그리고 유관상으로도 NV25SD2 처리구에서는 잎에 미미한 위조증상을 보였으나, NV25SD4 처리구에서는 위조증상이 매우 심하였다. 따라서 배추재배시 2주 동안 가뭄이 지속되면 엽육조직의 세포가 붕괴되고 신선도가 떨어지며 수확량이 줄어들기 때문에 2주간 가뭄이 지속된다면 관수를 해야 할 것으로 사료된다.
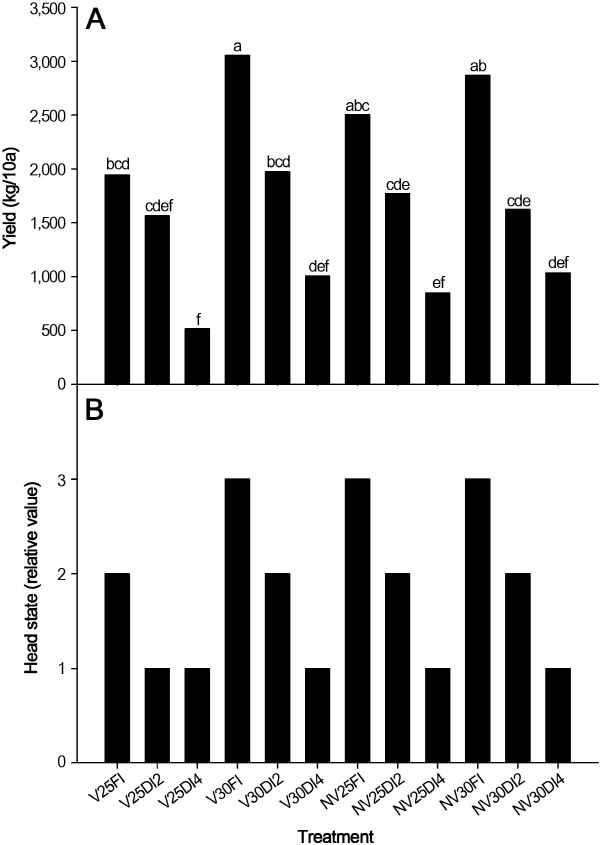
배추의 생장
토양수분 결핍처리 요인이 정식 후 60일 배추의 모든 조사 생장 특성에 통계적으로 유의하게 효과가 인정되었다. 그리고 재배기간의 온도처리 요인도 최대 엽장, 엽수, 엽면적 및 지상부의 생체중에 유의하게 영향을 주었다. 그러나 춘화처리의 효과는 모든 조사항목에서 인정되지 않았고, 3요인의 모든 조합 처리들에서 통계적으로 유의하게 영향을 미치지 못하였다(Table 2). 최대 엽장은 토양수분 결핍 4주 처리구(SD4)에서 작았다. 춘화처리를 하고 토양수분 결핍 처리를 4주 한 상온 및 고온 처리구들의(V25SD4와 V30SD4) 최대 엽장은 각각 30.2cm와 30.3cm로 모든 처리구 중에서 가장 작았다. 반면에 춘화처리를 하지 않고, 재배 관리 온도 30°C로 처리하고 충분히 관수된 조합 처리구(NV30FI)의 최대 엽장은 38.7cm로 가장 길었다. 엽수는 춘화처리를 한 여부와는 무관하게 재배 관리 온도를 25°C로 하고 토양수분 결핍 4주 처리구(V25SD4와 NV25SD4)에서 각각 46.7, 49.3매/주로 다른 처리구들에 비하여 유의하게 작았다(Table 2). 그러나 재배관리 온도를 30°C로 하고 관수를 충분히 하면 춘화처리에 무관하게 엽수를 많이 확보할 수 있었다(V30FI; 68.7매/주, NV30FI; 71.0매/주). 엽면적, 지상부의 생체중 및 건물중의 결과는 엽수의 영향을 받기 때문에 그 결과의 경향이 매우 유사하였다. 배추는 호냉성 채소로 생육기에 고온이 되면 결구가 되지 않고 병해에 약할 수 있으나(Opeña et al., 1988), 재배 기간 중 대기온도보다 4°C 또는 7°C를 높인 처리구에서 배추 엽수 발생 속도나 엽장이 긴 결과가 보고되었다(Oh et al., 2014). 일반적으로 온도가 증가할수록 작물의 세포 발생 및 활성도가 높아지는 것이 엽수 발생 속도와 잎의 신장이 촉진되는 결과로 이해할 수 있다. 그러나 극심한 온도 스트레스는 회복될 수 없는 세포손상을 유발하고 세포 사멸을 유도하기때문에 최적의 온도 관리가 배추의 생장을 촉진하는데 중요하다(Yang et al., 2006).
배추의 생장해석
토양수분 결핍처리는 배추의 상대생장률(relative growth rate; RGR), 단위 엽건물중 증가율(unit leaf rate; ULR), 엽면적비율(leaf area ratio; LAR) 및 비엽중(specific leaf area; SLA), 엽중률(leaf weight ratio; LWR)에서 통계적으로 유의하게 효과가 인정되었다(Table 3). 그리고 같은 재배기간의 춘화처리 효과는 ULR, LAR과 SLA에서 유의한 결과를 나타내었고, 온도처리 효과의 경우도 ULR, LAR과 LWR에서 유의성을 보였다. 정식 초기에 저온처리는 유묘의 생장을 저해시킬 수 있다는 보고가 있었고(Lee et al., 2011), 춘화처리 그리고 온도처리에 의한 초기 생장 차이가 생장해석 요인(ULR, LAR, SLA 및 LWR)들에 영향을 미쳐 통계적으로 유의차가 존재하였다. 그러나 춘화처리에 따른 엽면적은 처리간 유의차는 없었는데(Table 2), 이는 춘화처리 후 온도 및 토양 수분 결핍처리구에 영향을 받아 수확시에는 처리구들간 엽면적의 변이가 통계적으로 유의차가 나오지 않을 정도로 표준오차가 컸기 때문이었다.
세가지 처리 요인들(춘화, 온도 및 관수)의 모든 상호 작용 효과는 통계적으로 차이가 없었다. 일정시간의 작물 건물증가율을 나타내는 RGR 분석결과에서 지상부 상대생장률은 토양수분 결핍 4주 처리구들(SD4)에서 작게 나타났다. 춘화처리를 하고 25°C로 재배관리하에서 토양수분 결핍 처리를 4주간 한 처리구(V25SD4)에서 지상부 상대생장률은 0.092kg/kg/day로 모든 처리구들 중에서 가장 유의하게 낮았다. 반면에 충분히 관수된 처리구들(V30FI, NV25FI 및 NV30FI)은 모두 0.114kg/kg/day로 가장 높게 나타났다. 지하부 상대생장률은 지상부의 결과와 경향이 유사하였다. 본 실험의 결과와 동일하게 건조 스트레스 하에서 RGR의 감소에 대한 보고가 캐놀라(Habibi, 2014), 잠두(Ghassemi-Golezani et al., 2009), 콩(Li et al., 2013), 파인애플(Rebolledo-Martinez et al., 2005), 아킬레아(Khalil et al., 2011) 등의 작물에서 발표되었다. 또한, 고온 스트레스에 의한 RGR감소도 갯금불초(Song et al., 2010), 토마토(Kawasaki et al., 2013)에서 보고되었다. 가뭄과 고온은 식물이 생육 및 생존하는데 다양한 생리적 영향들을 주며, 그에 따른 효과들은 식물마다 상대적으로 차이를 나타내는 식물간 경합에 있어 중요한 요소들이다(Stasik and Jones, 2007). 이러한 환경 스트레스로 인한 RGR의 감소는 광합성과 생체중(Song et al., 2010), 엽내 질소함량(Karacic and Weih, 2006; Wieh and Nordh, 2002), 뿌리활력과 양분흡수(Nkansah and Ito, 1994) 감소로 인한 것이라 할 수 있다. 본 실험에서는 RGR이 고온에 따른 차이는 없었으나 가뭄에 의한 감소는 뚜렷하게 나타나 배추에서 가뭄에 의한 생육피해가 심하게 나타남을 확인할 수 있었다. 일정기간 동안 엽면적의 건물중 증가량을 의미하는 ULR은 춘화처리를 하고 30°C로 재배관리하에서 토양수분 결핍 처리를 2주간 한 처리구(V30SD2)에서 0.0052kg/m2/day로 가장 유의하게 낮았다(Table 3). 춘화처리와 고온(30°C)으로 재배관리시 단위 엽건물중 증가율이 낮은 경향을 보였다. 그리고 춘화처리를 하지 않고 재배관리 온도를 25°C로 낮춘 상태에서 토양수분 결핍 처리를 4주간 하였을 때 가장 높았다(NV25SD4; 0.0080kg/m2/day). 하지만, 해석결과가 평방미터당 g수준으로 형성되어 처리구들간의 실질적인 차이가 미비하다고 할 수 있다. 이와 반대로 LAR은 춘화처리를 하고 30°C로 재배관리하에서 토양수분 결핍 처리를 2주간 한 처리구(V30SD2)에서 32.30m2・kg-1로 가장 유의하게 높았고, 춘화처리를 하지 않고 재배관리 온도를 25°C로 낮춘 상태에서 토양수분 결핍 처리를 4주간 하였을 때 가장 낮았다(NV25SD4; 20.67m2・kg-1). SLA은 춘화처리를 하고 25°C로 재배관리하에서 토양수분 결핍 처리를 4주간 한 처리구(V25SD4)에서 1.154m2・kg-1로 가장 유의하게 높았다. 춘화처리를 하지 않고 재배관리 온도를 25°C로 낮춘 상태에서 충분히 관수를 하였을 때 가장 낮았으며(NV25FI; 0.756m2・kg-1), 충분히 관수가 이루어진 처리구들에서 SLA이 낮아지는 경향을 나타냈다. LWR은 춘화처리를 하고 30°C로 재배관리하에서 충분히 관수된 처리구(V30FI)에서 21.42kg・kg-1로 가장 유의하게 높았고, 춘화처리를 하고 25°C로 재배관리하에서 토양수분 결핍 처리를 4주간 한 처리구(V25SD4)에서 9.98kg・kg-1로 가장 낮았다. LWR의 경향은 충분히 관수된 처리구들(FI)이 전체적으로 높았고, 토양수분 부족이 오래 진행될수록(SD4) 낮아졌다. 토양수분 조건에 따른 엽면적 특성에 대한 연구들로 Rebolledo-Martinez et al.(2005)은 파인애플을 건조한 토양에서 재배하였을 때, 개화유도후 ULR은 낮아지고 LAR은 높아진다는 결과를 발표했고, Li et al. (2013)은 건조 스트레스에서 콩재배시 SLA과 LWR이 감소함을 보고했으며, Galmes et al.(2005)은 다양한 지중해 초화류와 관목류 육묘 시 파종 후 식물이 생장함에 따라 정상 수분 처리구와 수분 스트레스 처리구 간의 SLA, LAR, LWR 차이가 커졌다고 보고하였다. 이와 같은 건조에 의한 엽면적 감소 효과는 본 실험의 ULR, LAR과 LWR 결과에서도 확인할 수 있었다. 포플러(Karacic and Weih, 2006)와 버드나무(Wieh and Nordh, 2002)에서 RGR과 ULR이 양의 상관관계를 나타낸다는 보고와 같이 본 실험에서도 RGR과 ULR 모두 건조스트레스시 감소하는 경향을 나타내었다. 이는 RGR이 잎의 형태적 변화를 나타내는 LAR과 LWR에 양의 영향을 받기 때문으로 판단된다(Galmes et al., 2005).
그리고 초기 저온처리후 고온 조건시 배추 추대 여부를 조사한 결과 모든 처리에서 추대가 발생하지 않았다(결과 미 제시). 저온처리에 따른 추대 현상에 대해 Lee et al.(2011)은 배추 육묘시 야간온도 5와 10°C로 저온 처리하였을 때 추대현상이 관찰되지 않아 본 실험과 같은 결과를 보였다.
배추의 수량 및 결구 형성
배추의 수량은 춘화처리 후 고온에서 충분히 관수한 처리구(V30FI)에서 3,056kg/10a로 가장 많았으며, 토양수분 결핍 4주 처리구들의(V25SD4, V30SD4, NV25SD4 및 NV30SD4) 수량이 다른 처리구들에 비하여 적었다(Fig. 2A). 춘화처리 없이 적온(25°C)에서 충분히 관수한 처리구(NV25FI)의 수량을 100%로 하였을 때 토양수분 결핍 처리구(NV25SD2와 NV25SD4)들의 수량을 비교하면 각각 71% 및 34%였고, 단수 기간이 2주 증가하면 수량이 급격히 줄어든다는 결과를 얻었다. 그리고 춘화처리의 여부와 관계없이 온도관리를 고온(30°C)으로 하고 관수를 충분히 한 처리구들이(V30FI와 NV30FI) 적온(25°C)에서 충분히 관리한 처리구들 보다는 수량이 많았다. 배추 수확시 결구력을 조사한 결과, 단수처리구에서 중・하를 보였는데 특히 4주 단수처리구가 결구력이 약한 것으로 나타나 단수에 의한 식물체내 수분 부족은 결구력도 약하다는 것을 시사하였다. 충분히 관수를 한 처리구의 양배추의 생산량이 50.5톤/ha였으나 재배기간 중 관수를 하지 않은 처리구의 양배추 생산량은 17.5톤/ha로 절반 이상 감소하였다는 보고가 있었으며, 이러한 원인은 양배추 결구 중 수분 스트레스에 의하여 잎의 신장을 억제하여 엽면적과 엽수를 작아지게 하고 잎의 팽압과 삼투압을 낮추기 때문이었다(Maggio et al., 2005). 그리고 Sammis et al.(1988)은 상추와 배추에서 관수량을 늘렸을때 수량이 두 품목 모두 유의하게 증가한다는 결과를 보고하였다. 이는 가뭄과 같은 조건에서 상대적으로 낮은 증발산량이 배추와 상추의 건물생산량 감소의 원인으로 판단되며 이는 본 연구 결과와 유사하였다. 본 연구 결과, 여름철 고온기에 배추재배시 가뭄이 지속되면 생장이 지연되고, 결구가 불량해 지며 심할 경우 위조증상에 의한 결주로 수확량이 급격히 줄어들기 때문에 노지 포장에 관수를 효율적으로 할 수 있도록 재배적 기술이 현장에 투입되어야 할 것으로 판단되었다.