

서 언
재료 및 방법
버섯 배양・생육 및 상추 육묘 조건
버섯의 생육 시기 및 생육 온도에 따른 CO2 발생 속도 및 생육 특성 조사
버섯과 상추의 혼합 재배 실험 조건
결과 및 고찰
균긁기 후 경과 일수에 따른 버섯의 생육 특성
생육 시기 및 생육 온도에 따른 CO2 발생 특성
버섯과 상추의 혼합 재배 시 CO2 농도 변화 및 균형성 분석
서 언
버섯의 종균은 호기성 생명체로 호흡을 통해 CO2를 배출한다. 버섯재배 시 발생하는 고농도의 CO2는 버섯의 대 길이 생장과 갓 형태 발달에 나쁜 영향을 주기 때문에 환기를 통해 재배상 내부의 CO2 농도 관리가 필요하다(Jang et al., 2009). 환경적인 측면에서는 버섯재배 시 발생되는 고농도 CO2가 대기로 방출되어 온실 가스 문제를 야기시키기 때문에(Thavivongse and Buppachat, 2013) 이에 대한 대책이 필요하다.
밀폐생태계 생명지원시스템(Controlled Ecological Life Support System, CELSS) 분야에서는 인간의 호흡에 의해 발생한 CO2 흡수를 목적으로 작물을 활용하였으나(Daunicht, 1997; Tani and Kitaya, 1993), 적극적인 CO2 이용을 통해 작물의 효율적 재배를 유도하는 식물생산시스템과 개념적인 차이가 있었다(Son et al., 1999). 만약 고농도의 잉여 CO2를 식물생산에 직접 이용할 경우, 제기되었던 대기 중으로의 CO2 방출 문제들을 해결할 수 있게 된다. 이러한 관점에서 버섯과 상추를 재배하면서 두 작물 사이에서 이루어지는 CO2의 교환 양상을 추적하여 밀폐생태계 생명지원시스템의 하위 시스템에 관한 타당성을 검토하였으나(Kitaya et al., 1994), 활엽수 고목에 직접 표고버섯을 재배하였기 때문에, 공장형 재배방식에 사용되는 병 재배의 배지로부터의 CO2 발생이 고려되지 않았다. 새송이 버섯(Pleurotus eryngii (DC.) Quél)의 병 재배용 최적 배지 배합 비율 등의 배지와 균사에 관한 기본적 연구는 있으나(Kang et al., 2000; Lee et al., 2003), 배지로부터의 CO2 발생량을 분석한 예는 아직 없다.
상추의 경우 식물생산시스템에 관한 여러 연구에서 모델 작물로 사용되기 때문에(Kitaya et al., 1992; Massa et al., 2008; Seginer, 2003) 온도와 광도, CO2 등의 조건에 대한 생리적 특성들이 잘 알려져 있다. 따라서 버섯의 CO2 발생과 상추의 CO2 흡수 속도 비교를 통하여 지속적으로 두 작물을 재배할 수 있는 밀폐형 식물생산시스템을 설계하는 모델 작물로 사용하기에 적합하다.
버섯과 상추를 재배하는 밀폐형 식물생산시스템에서 CO2 농도 조건에 따른 작물의 품질 또한 고려하여야 할 요인 중의 하나이다. 새송이 버섯재배 시 2,400μmol・mol-1 이상의 CO2 농도 조건에서 버섯의 품질이 저하되고(Ryu et al., 2005), 상추는 CO2 농도가 높을수록 포화 형태로 광합성이 촉진되지만(Caporn, 1989), 작물이 사용하는 CO2를 생육시기별로 정량화함으로써 효율적인 CO2 공급 체계 구축이 필요하다. 따라서 버섯과 상추를 재배하는 밀폐형 혼합 식물생산시스템의 설계 시, 두 작물의 CO2 농도 조건에 맞도록 내부 CO2 농도 균형을 유지해 주어야 하며, 이를 위해서는 각 작물의 CO2 발생 속도와 흡수 속도에 근거한 개체 수의 조절이 필요하다.
본 연구는 새송이 버섯과 상추의 생육 시기에 따른 CO2 발생 속도와 흡수 속도를 측정하고, 배지의 CO2 발생 속도를 측정하여 밀폐형 혼합 식물생산시스템에서 두 작물의 재배 비율에 따른 CO2 농도 변화와 균형성의 분석을 목적으로 하였다.
재료 및 방법
버섯 배양・생육 및 상추 육묘 조건
본 실험에 사용한 버섯은 아인농산(경기도 안성시 금광면 소재)에서 재배한 새송이 버섯(Pleurotus eryngii(DC.) Quél)이며, 공시 균주는 경기농업기술원 버섯연구소의 새송이 2호이었다. 사용된 배지는 함수율이 65-68%이며, 톱밥:콘코프:첨가제의 비율이 1:1:2(v/v/v, %)이며, 첨가제는 비트펄프, 대두박, 밀기울, 건비지 등이 포함되었다. 950mL 폴리프로필렌 병에 충진된 570g의 배지는 121°C에서 60-90분간 살균하였다.
균사가 접종된 배지는 26-28일간 배양하였으며, 5-6일간 후배양 하였다. 배양실은 온도 21-22℃, 습도 65-70%를 유지하였다. 균 긁기 이후 생육실의 온도는 18°C에서 점차 15°C까지 낮추었으며, 습도는 초기 95%에서 후기에는 80-85%로 낮추었다. 육안으로 식별 가능한 크기의 자실체가 형성되는 시점(균 긁기 후 7일차)부터 이에 해당하는 버섯을 선별하여 실험에 사용하였다.
시험에 사용한 상추는 아시아 흑로메인 상추(Lactuca sativa L. cv. Asia Heuk Romaine, Asia Seed Company, Seoul, Korea) 품종을 스펀지 배지에 파종하여 3주간의 육묘 기간을 거쳐 정식 후 7일차의 본엽 10매된 것을 이용하였다. 담액식 수경 방식으로 육묘하였으며, 파종 후 첫 1주 동안에는 양액을 공급하지 않았고, 2주차부터 3주차까지는 EC 0.6dS・m-1의 야마자키 배양액을 공급하였다. 육묘상은 PPF 200μmol・m-2・s-1의 광도가 유지되도록 형광등을 사용하여 광을 조사하였으며, 광주기는 명기를 16시간, 암기를 8시간으로 설정하였다. 육묘상의 온도는 22°C, 습도는 70%로 유지하였으며, CO2 농도는 400μmol・mol-1으로 유지하였다.
버섯의 생육 시기 및 생육 온도에 따른 CO2 발생 속도 및 생육 특성 조사
버섯이 발생시키는 CO2의 변화를 측정하기 위하여 내부 온도를 일정하게 유지할 수 있는 밀폐형 아크릴 챔버(1.0m × 0.8m × 0.5m)에 버섯을 넣고 밀폐하였다. 계측과 제어는 데이터로거(CR1000, Campbell Scientific Inc, Logan, UT, USA)를 이용하였으며, CO2 측정 센서(LI-820, LI-COR, Lincoln, NE, USA)를 설치하였다.
균 긁기 후 경과 일수에 따른 버섯 자실체의 생육 특성을 분석하기 위하여 생체중과 갓 직경, 대 길이에 대한 측정을 실시하였다. 생체중의 경우 병 무게를 제외한 배지와 균사, 자실체가 포함된 무게를 측정한 뒤, 자실체를 제거하여 계산하였다. 갓 직경의 경우 타원체로 가정하여 장축과 단축의 길이를 측정한 뒤 2대의 평균값을 사용하였다. 갓 직경과 대 길이는 솎아내기를 실시한 자실체를 대상으로 하여 균 긁기 후 13, 15, 17일차에 측정하였다.
생육 시기별 CO2 발생 속도 측정을 실시한 버섯은 균 긁기 후 7, 9, 11, 13, 15, 17일차의 것을 이용하였으며, 균 긁기 후 13일차 되는 날에 2.5-3.0cm까지 커진 자실체 2대를 남기고 나머지를 솎아내는 작업을 실시하였다. 각 생육 시기별로 챔버 내부에 버섯을 5병씩 넣고, 3반복 측정하였다. CO2 농도를 5초 간격으로 측정하여 버섯의 CO2 발생 속도를 계산하였다. 배지의 CO2 발생 속도는 앞서 CO2 발생 속도 측정이 끝난 동일한 병의 버섯의 자실체 부위를 제거한 후 배지와 균사 부분을 다시 밀폐 챔버에 넣어 같은 방식으로 측정하였다.
|
Fig. 1. A closed plant production system cultivation consisting of two growth modules for the mushrooms (A) and lettuces (B). Air is circulated between the two modules exchanging CO2, O2, and H2O. |

생육 온도별 CO2 발생 속도 측정을 실시한 버섯은 균 긁기 후 13, 14일차의 것을 이용하였다. 생육 온도는 새송이 버섯의 생육 적온과 흑로메인 상추의 생육 적온을 포함하는 범위인 16.5°C, 18°C, 21°C, 24°C로 챔버 내부의 온도를 유지시켰다(Kim et al., 2004, 2012). 또한 솎아내기 작업의 영향을 알아보기 위하여 균 긁기 후 13일차의 버섯은 솎아내지 않은 상태로, 균 긁기 후 14일차의 버섯은 솎아내기 작업을 실시한 상태로 측정하였다. 각 측정 별로 챔버 내부에 버섯을 3병씩 넣고, 3반복 측정하였다. CO2 농도를 5초 간격으로 측정하여 버섯 1병당 CO2 발생 속도를 계산하였다. 실험에 사용된 버섯 병당 전체 생체중 값은 Table 1과 같다.
버섯과 상추의 혼합 재배 실험 조건
두 작물을 혼합 재배하며 CO2의 변화를 측정하기 위하여 내부 온도가 유지되는 밀폐형 아크릴 챔버(1.0m × 0.8m × 0.5m)를 2개 사용하여 CO2를 순환시키면서, 버섯과 상추를 재배하였다(Fig. 1). 각각의 챔버 내부 온도는 펠티에 소자를 이용하여 18°C와 22°C로 조절하였다. 버섯재배 챔버는 생육실 조건과 유사하도록 암막을 이용하여 암조건을 만들어 주었으며, 내부 습도는 65-70%로 유지하였다. 상추재배 챔버에는 담액 수경을 실시할 수 있도록 양액 탱크를 배치하였으며, 양액의 EC는 1.2dS・m-1가 유지되도록 조절하였다. 챔버 내부에 상추를 재식간격 10cm × 10cm로 하였으며, 온도 조절용 펠티에 소자에 응결된 내부 수분이 다시 양액탱크로 순환할 수 있도록 하였으며 내부 습도는 70%로 유지하였다. 광조건의 경우 LED(FGL-B1200, FC POIBE, Seoul, Korea)를 이용하여 적색:청색:백색 광의 비율을 8:1:1로 적용하였으며, 총 광량이 PPF 340μmol・m-2・s-1가 유지되도록 설정하였다. Kim et al.(2012)에 따르면 같은 광 조건 하에서 생육 온도 21°C일 때, 상추의 CO2 흡수 속도는 정식 후 5일차에 7.0μgCO2・s-1이고, 정식 후 10일차에 16.1μgCO2・s-1이다. 이 값들과 측정된 버섯의 CO2 발생 속도를 비교하여 실험에 사용할 버섯의 병 수, 상추의 개체 수를 Table 2와 같이 결정하였다.
펌프 내부의 오일 등에 의한 챔버 내부 공기 오염 문제 등 방지하기 위하여 공기 순환에는 다이아프램(diaphragm) 펌프를 사용하였다(Son and Park, 2001). 62L・min-1의 유속의 다이아프램 펌프(Boxer 7004, Uno International Ltd., London, UK) 2대 이용하여 두 챔버 사이의 기체를 순환시켰다. 순환된 기체가 각 챔버 내부에서 균일하게 혼합될 수 있도록 환기팬을 장착하여 내부 풍속을 0.3m・s-1로 유지시켰다.
밀폐형 혼합 생산시스템의 계측과 제어를 위하여 데이터 로거(CR1000, Campbell Scientific Inc., Logan, UT, USA)를 사용하였으며, 버섯 챔버와 상추 챔버의 CO2 농도를 2분 간격으로 순환적으로 측정할 수 있도록 CO2 측정 센서(LI-820, LI-COR, Lincoln, NE, USA)를 설치하였다. 각 챔버 내의 CO2 농도를 측정하여 5초 간격으로 데이터를 저장하였다. 챔버 내부의 CO2 농도에 따라 다이아프램 펌프의 작동을 유도할 수 있도록 AC/DC 컨트롤러(SDM-CD16AC, Campbell Scientific Inc., Logan, UT, USA)를 장치하여 두 챔버 사이의 기체 교환에 관한 자동화 시스템을 구축하였다. 버섯 생육 챔버의 경우 2,400μmol・mol-1 이상의 CO2 농도에서 버섯의 품질이 감소한다는 연구 결과(Ryu et al., 2005)에 근거하여, 기간에 따라 CO2 농도 2,000-2,500μmol・mol-1을 펌프 작동 값으로 설정하였으며, 상추의 경우 대기 중 CO2 농도인 400μmol・mol-1 이하로 내려가면 펌프를 작동하여 버섯과 상추 챔버의 내부 기체를 순환시킬 수 있도록 설정하였다. 생육 실험을 진행하는 동안 버섯과 상추의 재배 비율을 변화시키며 각 챔버의 CO2 농도 변화 양상을 기록하였다.
실험에 사용된 상추의 생육 특성을 알아보기 위하여 실험에 들어가기 직전 3개체에 대하여 지상부의 생체중을 측정하였고, 엽면적 측정기(LI-3100C, LI-COR, Lincoln, NE, USA)를 이용하여 엽면적을 측정하였다. 생육 실험이 끝난 후 9개체에 대하여 같은 방식으로 지상부의 생체중과 엽면적을 측정하였다. 또한 측정한 상추의 잎을 80°C에서 6일간 건조하여 건조 중량을 측정하였다(Table 3).
결과 및 고찰
균긁기 후 경과 일수에 따른 버섯의 생육 특성
솎음 처리 후 수확하기 전까지 자실체 무게는 지수 함수적인 증가 양상을 보였지만, 갓 직경과 대 길이는 선형적으로 증가하였다(Table 4). 자실체의 무게를 이용한 국립농산물품질관리원 농산물 표준 규격상 균 긁기 후 17일차 자실체는 크기 L에 해당하지만(NAQS, 2012), 갓 직경과 대 길이에 대한 기준이 없기 때문에 표준화된 품질을 측정하기 어렵다. 이를 보완하고자 Ryu et al.(2006)이 제시한 등급 기준을 따르면, 실험에 사용된 균 긁기 후 17일차인 새송이 버섯의 자실체 등급은 대 길이 기준으로 상품이거나, 무게 기준으로 특품에 해당하는 것으로 나타났다.
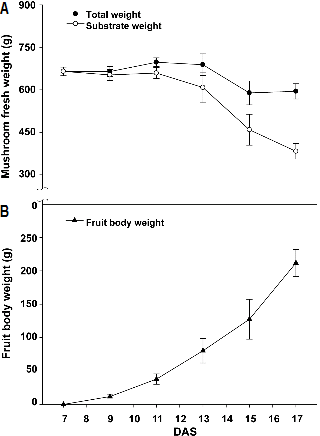
자실체가 발달하는 동안 1병에 대한 전체 중량과 배지 중량, 자실체 중량을 측정한 결과는 Fig. 2와 같다. 총 중량은 균 긁기 후 7일차부터 13일차까지 증가하였지만, 13일차에 솎아내기를 실시한 이후 감소하였다. 배지 중량은 균 긁기 후 11일차부터 17일차까지 감소하였고, 균 긁기 후 13일차에 솎아내기 작업을 하며 잘라낸 자실체의 평균 무게는 52.9g으로 나타났다. 감소한 배지 중량의 대부분은 수분으로, 자실체의 생체중 증가에 활용된 것으로 판단된다. 새송이 버섯의 균사 생장에 미치는 영향에 대해 조사한 결과에서 톱밥 배지의 수분 함량이 65%의 배지 함수율에서 균사 생장이 가장 좋은 것으로 나타났다(Kim et al., 1997). 따라서 자실체의 솎음을 통한 중량 손실 이외에도 배지의 함수율 변화에 따른 중량 변화도 고려하여야 할 것이다.
|
Fig. 2. Changes in total fresh weight, substrate weight (A) and fresh fruit body weight (B) of the mushrooms with days after scratching (DAS). Vertical bars represent the mean ± SD (n = 3). |
생육 시기 및 생육 온도에 따른 CO2 발생 특성
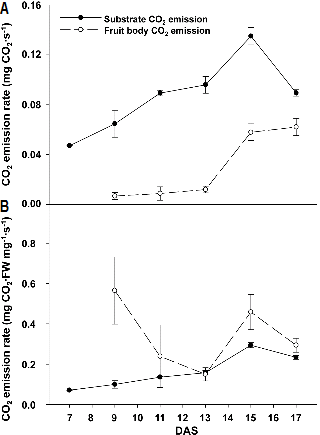
자실체 발달 기간 중 버섯 1병당 CO2 발생 속도를 측정한 결과 솎아내기 작업 전까지는 배지의 CO2 발생 속도가 자실체의 CO2 발생 속도에 비해 높았으나, 출하 시기인 균 긁기 후 17일차에는 그 차이가 줄어든 것으로 나타났다(Fig. 3A). 반면 Fig. 2에 나타낸 생체중으로 버섯의 CO2 발생 속도를 나누어 계산한 생체중 g당 CO2 발생 속도는 자실체가 배지에 비해 높은 값을 나타내었으며, 자실체 발달 초기인 균 긁기 후 9일차부터 13일차까지 점차 감소하였다(Fig. 3B). 균 긁기 후 13일차에 솎음 작업을 실시하면 자실체의 생체중 mg당 CO2 발생 속도가 약 3.1배 증가하다가 균 긁기 후 17일에 이르러 다시 감소하는 형태를 보였다. 이 결과를 통해 솎음 처리가 2일간에 걸쳐 남은 자실체의 호흡 활동을 촉진하는 것으로 추정할 수 있었다. 그러나 버섯의 호흡 속도 측정은 수확 후 자실체 품질 유지를 위하여 실시된 경우가 대부분이며(Lee et al., 2003) 자실체 발달 기간에 따른 CO2 발생량을 측정한 연구는 이루어지지 않았기 때문에, 솎음 처리가 버섯 자실체의 호흡 활동을 촉진하는 원인은 찾아낼 수 없었다.
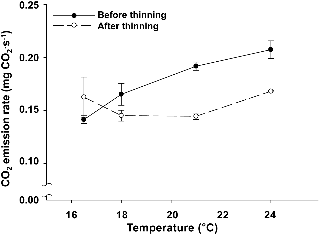
생육 온도에 따른 버섯 1병의 CO2 발생 속도는, 솎아내기 전 버섯 1병당 CO2 발생속도는 온도증가에 따라 증가하였으나, 솎아내기 후에는 16.5°C와 24°C에서 높은 CO2 발생 속도를 보이며, 18°C와 21°C의 온도에서 CO2 발생 속도가 낮아졌다(Fig. 4). 버섯보다 고온으로 설정된 상추 챔버 내부의 공기가 버섯에 도달할 경우, 솎아내기 전 버섯은 CO2 발생 속도가 약 1.4배 상승하는데 반해, 솎아낸 후에는 거의 일정하다는 것을 알 수 있다. 따라서 상추 챔버에서 버섯 챔버로 이동하는 공기를 냉각시켜 주는 장치를 설치하여 솎아내기 전 기간 버섯의 CO2 발생 속도를 유지시킬 필요가 있다. CO2가 새송이 버섯에 미치는 영은 주로 균사체에 연구 대상이었으며, 자실체에 관해서는 주로 Agaricus bisporus(양송이 버섯)등이 연구되었다(Jang et al., 2007). 국내외 새송이 버섯 병재배 관련 연구에서 Ryu et al.(2005)이 실시한 솎음 처리 조건 시 CO2 농도에 따른 자실체 특성 평가를 제외하고 솎음 처리에 의한 새송이 버섯 자실체의 CO2 발생 특성에 관한 연구가 추후 필요하다.
|
Fig. 4. Changes in CO2 emission rate per bottle before and after thinning of the mushrooms with growth temperature. Vertical bars represent the mean ± SD (n = 3). |
버섯과 상추의 혼합 재배 시 CO2 농도 변화 및 균형성 분석
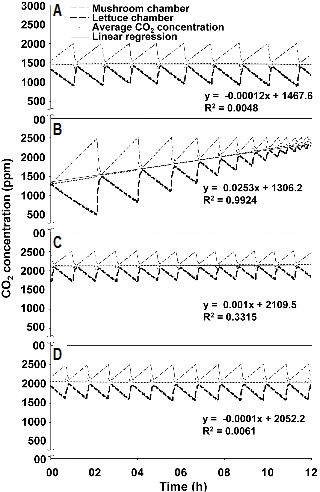
버섯과 상추의 혼합재배 시, 각 챔버 내부 CO2 농도 변화와 두 챔버의 평균 CO2 농도 변화에 대한 선형 회귀 분석 결과를 통하여 CO2 균형 유지 여부를 판단하였다(Fig. 5). 회귀 계수 a값이 0에 가까울수록 두 챔버 사이에 CO2 발생 및 흡수 속도가 평형에 가깝게 된다. 균 긁기 후 9일차 버섯 3병과 상추 9개체, 균 긁기 후 12일차 버섯 2병과 상추 9개체, 균 긁기 후 14일차 버섯 2병과 상추 11개체는 CO2 발생 및 흡수 속도의 평형을 이루는 것으로 나타났다. 즉, 균 긁기 후 9, 12, 14일차 새 송이 버섯 1병의 CO2 발생 속도는 정식 후 7, 10, 12일차인 상추 3, 4.5, 5.5개체의 CO2 흡수 속도에 대응한다.
반면 균 긁기 후 10일차 버섯 4병과 상추 11개체는 버섯의 CO2 발생 속도가 상추의 CO2 흡수 속도에 비해 크기 때문에 밀폐 챔버 내부의 CO2 발생 및 흡수 속도 평형이 이루어지지 않았다. 이 경우 강제 환기를 실시하지 않으면 밀폐형 식물생산시스템 내의 CO2 농도가 버섯의 품질에 영향을 미치는 농도 이상으로 상승하므로, 외부 공기를 유입하여 CO2 농도를 낮추거나 상추를 추가적으로 시스템 내로 투입할 필요가 있다. Jang et al.(2009)에 따르면 느타리버섯의 경우 생육 온도가 높아질수록, 생육 단계가 진전될수록 호흡량이 많아지기 때문에 CO2에 의한 환기 장해 현상이 일어나므로 생육 후기로 갈수록 환기량을 늘려야 한다. 실제 농가에서 상추와 버섯의 동시 재배를 실시하기 위해서는 환기의 효과를 추가적으로 고려할 필요가 있다.
Yang et al.(2012)과 Boswell et al.(2003)은 온도와 습도에 따른 새송이 버섯 균사 미세 구조와 생장 방식에 대한 모델을 구축한 바 있지만, 자실체의 생장에 관한 부분 모델이 포함되어 있지 않았다. 따라서 새송이 버섯의 배지와 자실체의 CO2 발생 속도에 대한 모델을 확립한 후, 상추의 CO2 흡수속도 모델과 비교하여 두 작물의 적정 재배 비율을 도출한다면, 효율적인 밀폐형 혼합 생산시스템의 구축이 가능할 것으로 판단된다.
