

서 언
재료 및 방법
식물재료 및 재배조건
생육특성 조사
식물체내 무기성분 함량의 분석
광합성 특성의 분석
통계분석
결과 및 고찰
월동무의 생육조건
월동무의 생육특성
식물체의 무기성분 조성
광합성 특성
서 언
농작물의 생산량은 생육과정에 영향을 미치는 주산지 기후와 갑작스런 한파나 고온과 같은 자연재해 발생 여부에 따라 크게 달라지며, 이러한 변화는 작물 수급에도 영향을 주어 가격 변동을 심화시키게 된다. 식물 생육에 있어 최적 온도 이상에서는 기온 상승이 작물의 재배기간을 단축시켜 바이오매스 축적을 위한 시간이 감소되 어 작물의 생산에 부정적으로 작용할 수 있다. 그러나 추운 지역이나 계절적으로 겨울철에는 기온 상승이 작물의 생산에 있어 우호적으로 작용하기도 한다(Ahad and Reshi, 2015). 기후변화에 관한 정부간 패널 보고서(IPCC, 2007)에 의하면 지구온난화 로 인하여 지구 전체의 평균 온도는 매 10년 마다 0.3°C씩 상승하고, 50 ? 100년 후에는 현재의 기온보다 1.6 - 6.4°C가 높아질 것으로 예상하고 있다. 최근 30년(1981 - 2010) 동안 우리나라의 연평균 온도가 0.7°C 상승하였고, 앞으로도 지속적으로 상승 할 것으로 예측된다(KMA, 2011).
무(Raphanus sativus var. hortensis)는 배추과(Brassicaceae)에 속하는 작물로써 뿌리뿐만 아니라 잎까지 식용이 가능한 대 표적인 근채류이며(Prakash and Hinata, 1980), 국내에서는 배추(Brassica campestris ssp. napus var. pekinensis)와 함께 김치 의 주재료로 사용되고 있는 중요한 채소이다. 더군다나 무는 국내에서뿐만 아니라 전 세계적으로 널리 생산되어 소비되고 있 으며, 각 나라의 기후와 기호에 맞게 다양한 품종들이 재배되고 있으며, 한국, 중국, 일본 등 동아시아에서는 주로 흰색의 크기 가 크고 긴 뿌리의 품종을 많이 재배하고 있다. 국내에서는 재배 작형에 따라 봄무, 고랭지무(여름무), 가을무, 월동무로 구분하 여 연중 생산되고 있으며, 제주에서는 월동무의 재배면적과 생산량이 다른 작형에 비해 월등히 많은 편이다(Ko and Kim, 2013). 월동무는 주로 9월 하순 또는 10월 상순에 파종하여 12월부터 익년 4월까지 수확하여 출하되고 있다. 무는 생육기간 중 10°C 이하의 저온에 장기간 노출되었을 때 조기 개화(추대형성)가 야기되며(Engelen - Eigles and Erwin, 1997), 이러한 현상 은 무의 생산성 및 상품성 저하의 직접적인 요인이 된다. 최근 겨울철에 빈번히 발생하는 폭설과 혹한, 일조부족, 잦은 강우 등 의 이상 기상의 영향으로 초기 생육부진과 추대 발생, 저온피해 등이 나타나 수량이 감소하고 품질이 저하되는 것으로 보고되 었다(Kim et al., 2013). 그러나, 제주지역은 아열대기후대로 겨울철 온도가 높을 뿐 아니라 온난화도 여느 지역에 비해 빠르게 진행되고 있어(JRMA, 2011), 기후온난화가 겨울철 월동무 재배시 저온 피해를 저감할 수 있는 요인으로 작용할 수 있을 것으 로 보인다. 따라서, 미래의 기후온난화에 대비한 월동무의 상승 온도에 대한 반응을 연구할 필요가 있다.
기후온난화에 의한 식물의 반응을 파악하기 위하여 작물의 재배조건 규명, 수확량 변동예측 모형 개발 등에 관한 연구들이 활발하게 이루어지고 있으며, 상승온도 조건을 설정하기 위하여 온도구배터널(temperature gradient tunnel system)이 이용되 고 있다. 온도구배터널은 온도를 제외한 다른 환경인자(광량, 대기 CO2 농도 등)를 자연 상태와 유사하게 유지할 수 있도록 고 안한 반폐쇄형 장치이다. 즉, 입구를 개방하여 외부 공기가 터널 내부로 유입되도록 하고 후미부에 6개의 소형 환기팬을 설치 하여 배기 속도를 제어함으로써 공기 흡입구에서부터 후미부 쪽으로 연속적인 온도 구배가 형성되도록 설계되어 대기온도 이 상의 고온에 대한 작물의 반응을 연구하는데 유용한 시스템이다. 본 연구진은 우리나라에서 생산 및 소비가 높은 채소 중에 하 나인 배추를 대상으로 온도구배터널을 이용하여 기후변화 시나리오에 의해 예측되는 봄철 온도 상승이 식물의 생장에 미치는 영향을 정량적으로 분석하여 보고한 바 있다( Oh et al., 2014).
본 연구에서는 온도구배터널 내에서 자라는 월동무의 생장, 광합성 및 무기성분 조성과 흡수 등을 조사하여 미래의 기후온 난화가 월동무 재배에 미치는 영향을 예측하고자 하였다.
재료 및 방법
식물재료 및 재배조건
무는 영동무(Raphanus sativus var. hortensis cv. Youngdong, Nongwoobio, Korea) 품종을 사용하였으며, 종자는 2013년 10 월 10일에 온도구배터널(25m × 3m × 3m) 내의 암갈색 비화산회토(동홍통, 미사질양토)에 50cm 간격으로 직파하였다. 온도 구배터널 내 토양은 종자를 파종하기 15일 전에 돈분발효퇴비(20kg·100m-2)와 복합비료(NPK 21 - 17 - 17, 5.2kg·100m-2)를 시 용하고 토심 20cm 깊이로 경운하여 토양과 고르게 혼합한 후 30cm 높이의 두둑을 만들었다. 토양수분은 재배기간 동안 2-?3일 간격으로 관수하여 포장 용수량 정도를 유지하였으며, 진딧물 등 병충해 방제를 위하여 재배기간 중 저독성 약제 인코니 도(0.5g·L-1, Hannong)와 오신(0.5g·L-1, Hannong)을 각각 2회씩 살포하였다.
온도구배터널은 농촌진흥청 온난화대응농업연구소(N33˚28´ E126˚31´, 해발고도 200m, Jeju, Korea)의 불소 필름으로 피복 된 시설을 이용하였으며, 온도는 터널의 입구, 중앙부, 후미부에 지상부로부터 2.5m 높이에 설치한 온도센서(1400 ? 101, LI - COR Inc., USA)를 이용하여 측정하였다. 시험기간 동안 터널 입구에 비해 중앙부, 후미부에서는 온도가 각각 4°C와 7°C 더 높게 유지되도록 복합환경제어시스템(TGC - Soldan, Soldan Crop., Korea)을 이용하여 제어하였다. 일사량은 지상부로부터 2.5m 높이에 설치한 광센서(LI200X, LI ? COR Inc., USA)를 이용해 측정하고 일일 적산일사량으로 나타내었다. 재배기간 (2013년 10월 10일 ~ 2014년 1월 20일) 동안의 온도와 일사량은 데이터로거(CR1000, Campbell Scientific Inc., USA)에 연결하 여 1시간 간격으로 기록하였다.
생육특성 조사
무의 생육특성은 파종 후 100일 후(2014년 1월 20일)에 터널 입구를 대조구로 하고 중앙부(대기온도 +4°C), 후미부(대기 온도 +7°C)에서 각각 10개체를 무작위로 선발하여 수확하고, 초장, 뿌리의 길이와 직경, 바이오매스, 엽수를 조사하였다. 식 물의 바이오매스는 지상부와 뿌리로 구분하여 각각 생체중을 측정한 후, 70°C 건조실에서 5일간 건조시켜 건물중을 조사 하였다.
식물체내 무기성분 함량의 분석
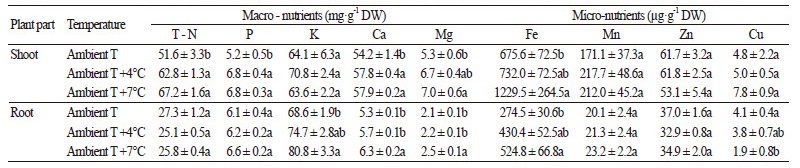
무기성분 함량은 농촌진흥청 농업과학기술원 표준분석법에 준하여 분석하였다(NIAST, 2000). 식물체는 지상부와 뿌리로 구분하여 건조하고 소형파쇄기(HR2860, Philips, China)로 균일하게 마쇄하여 분석시료로 사용하였다. 전질소 함량은 Kjeldahl 법, 유효인산은 Lancaster 법에 의해 분석하였으며, 치환성 양이온(K, Ca, Mg, Na)과 미량원소(Fe, Mn, Zn, Cu)는 분해용액 (HNO3 : H2SO4 : HClO4 = 10 : 1 : 4)을 첨가하여 전처리 및 여과과정을 거친 후 유도결합플라즈마 발광광도계(Inductively Coupled Plasma Spectrophotometry ? Mass, ICP ? Integra XL, GBC Scientific Equipment Pty Ltd., Australia)를 이용하여 분석하였으며, 건물 중 1g당 함량으로 제시하였다.
광합성 특성의 분석
CO2 고정률을 비롯한 기공전도도, 증산률, 수분이용효율은 660nm의 LED 광원이 부착된 휴대용 광합성 측정기 LCpro+ Portable Photosynthesis System(ADC Bio Scientific Ltd., UK)을 사용하여 20°C 조건에서 1,300μmol·m-2·s-1의 포화광을 조사하여 측정하였다. 그리고, 각 처리별로 광량 변화에 대한 CO2 고정률을 측정하여 광반응 곡선을 작성하고, 이를 토대로 최대광합 성률(maximum photosynthetic rate, Amax)과 호흡률(dark respiration rate, Rd)을 산출하여 제시하였다(Marshall and Biscoe, 1980; Oh et al., 2015).
통계분석
광반응 곡선은 SigmaPlot 10.0 통계 소프트웨어(Software, Erkrath, Germany)를 이용하여 쌍곡선 방정식을 기준으로 곡선 피 팅하여 나타내었다. 통계분석은 SPSS 통계 패키지 18.0(SPSS, Chicago, IL, USA)를 이용하여 수행하였으며, ANOVA에 이어 서 Duncan의 다중검정( p < 0.05)으로 평균치 간의 차이에 대한 유의성을 검정하였다.
결과 및 고찰
월동무의 생육조건
생육 기간 동안 온도구배터널 내부의 일일적산일사량은 흐리거나 비 또는 눈이 내린 날을 제외하고는 전반적으로 3,000W·m-2를 상회하였으며, 일평균 대기 온도는 터널 입구의 대기온도를 기준으로 중앙부, 후미부의 온도가 주야를 통하여 각각 4.5°C, 6.2°C 높게 유지되고 있는 것을 확인할 수 있었다(Fig. 1). 월동무의 재배기간 동안 터널 입구의 일평균 대기온도 가 8.9°C(일최저평균 ? 0.1°C, 일최고평균 19.9°C)를 나타내었으며, 터널의 중앙부에서는 일평균 13.4°C(일최저평균 4.3°C, 일최고평균 24.6°C), 후미부에서는 일평균 15.1°C(일최저평균 6.0°C, 일최고평균 25.6°C)로 터널 입구에 비해 각각 4.5°C와 6.2°C가 높았다. 그리고 재배기간 동안 터널 입구에는 일평균 20°C 이상을 넘는 날이 없는데 반하여 중앙부는 20°C 이상의 일수가 18일, 후미부는 26일이었다. 그리고 터널 입구에서는 11월 중순 이후로 일평균 10°C 이하의 온도에 지속적으로 노출 되는데 반해, 터널의 중앙부와 후미부에서는 10°C 이하의 일수가 각각 29일과 18일에 불과하였으며 주로 12월 중순 이후에 나타났다.
Fig. 1.
Changes in daily solar radiation (A) and daily mean air temperature (B) in a temperature gradient tunnel (TGT) used for radish cultivation. Dotted line in (B) represents daily mean ambient temperature in the entrance of the TGT; gray and black solid lines represent ambient temperatures in the middle and tail patrs of the TGT, respectively.
월동무의 생육특성
월동무의 지상부와 뿌리 모두 대기온도에서 보다 대기온도 +4°C와 대기온도 +7°C 조건에서 생장이 더 왕성하였다(Table 1). 지상부의 생체중과 초장은 대기온도에서 보다 대기온도 +4°C와 대기온도 +7°C 조건에서 유의하게 더 무겁고 길었다. 지상 부의 수분함량도 대기온도 +4°C와 대기온도 +7°C 조건에서 좀 더 높았다. 엽수와 건물중은 온도가 높아졌을 때 다소 증가 하였으나 통계적으로 유의한 차이는 없었다. 뿌리는 생체중뿐만 아니라 건물중도 대기온도에서 보다 대기온도 +4°C와 대 기온도 +7°C 조건에서 1.4배 이상 더 무거웠으며 뿌리의 길이와 직경 또한 1.2배 이상 더 컸다. 수분함량도 대기온도 +7°C 조건에서는 유의하게 높았다.
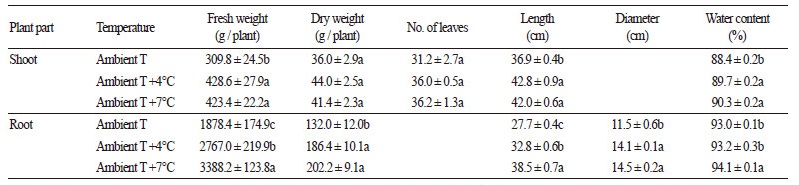
무의 생육 적온이 15 ? 20°C 범위에 해당하는 것으로 알려진 바(RDA, 2005), 겨울철 대기온도 조건에서 재배된 무의 생육 이 저조한 것은 Fig. 1에서 살펴본 바와 같이 생육기간 중 10°C 이하의 저온이 장기간 지속된 데에 기인하는 것으로 보인다. 따라서, 겨울철 상승 온도는 제주도 내 월동무의 재배에 우호적으로 작용할 것으로 보인다.
식물체의 무기성분 조성
다량원소 중에 Ca, Mg의 함량은 지상부와 뿌리에서 모두 대기온도에서 보다 대기온도 +4°C와 대기온도 +7°C 조건에서 재배하였을 때 증가하였다. 반면에, T - N, P 함량은 지상부에서는 대기온도에서 보다 대기온도 +4°C와 대기온도 +7°C 조 건에서 재배하였을 때 증가하였으며, 뿌리에서는 K 함량이 겨울철 온도가 높아졌을 때 증가하는 양상을 보였다. 그리고, 미 량원소 중에서 Fe의 함량은 지상부와 뿌리에서 모두 온도가 높아졌을 때 증가하는 반면에, Cu 함량은 뿌리에서 온도가 높아 짐에 따라 감소하였다(Table 2). 이러한 뿌리에서 Cu 함량의 감소는 춘파소맥(Triticum aestivum)의 경우에서 처럼 뿌리의 Cu 가 지상부로 전이된 결과로 보인다(Boatwright et al., 1976). 식물에서 무기양분의 흡수는 작물의 종류와 계절에 따라 차이가있고, 근권온도, pH, 토양의 무기양분 조성과 농도 등에 의해 영향을 받는 것으로 알려져 있다(Jarmal and Poldma, 2004; Tindall et al., 1990; Yildirim et al., 2001). 또한 생육적온 범위를 벗어난 온도에서 작물을 재배하였을 때 양분과 수분의 흡수가 저해되 기도 한다(Nakano, 2007). 본 연구에서 겨울철 온도 상승은 무 식물체의 T - N, P, K, Ca, Mg, Fe의 흡수를 증가시키고, 지상부 쪽으로 Cu의 전이가 일어나는 것으로 나타났다. 이와 같이 온도 상승이 양분 흡수를 촉진하는 예는 팔레놉시스(Phalaenopsis amabilis)에서도 보고되었는데, 근권의 온도 환경에 의해 N와 P보다는 K, Ca, Mg의 흡수에 크게 영향을 받으며, K 함량은 20°C 에서, Ca과 Mg의 함량은 25°C에서 가장 높았으며 팔레놉시스의 생육에 필요한 적정 양분 흡수는 20 - 25°C의 근권온도 범 위에서 이루어진다(Lee et al., 2011). 따라서 겨울철 상승 온도는 Table 1과 Table 2에서 보는 바와 같이 월동무의 재배에 있어 생육적온에 가깝기 때문에 생장뿐만 아니라 양분 흡수에도 긍정적인 효과가 있을 것으로 판단된다.
광합성 특성
CO2 고정률과 증산률은 터널 입구에서 겨울철 대기온도에 적응된 식물보다 대기온도 +4°C와 대기온도 +7°C 조건에서 재배하였을 때 다소 증가한 반면에, 수분이용효율은 감소하였다. 기공전도도는 재배조건에 따라 통계적으로 유의한 차이가 없었다(Fig. 2). 무의 최대광합성률(Amax)은 겨울철 대기온도에서 자란 잎에서 9.7μmol·m-2·s-1로 낮은데 반하여 대기온도 +4°C 와 대기온도 +7°C 조건에서는 각각 22.1μmol·m-2·s-1과 22.9μmol·m-2·s-1로 2배 이상 증가하였다. 호흡률(Rd)은 대기온도 +4°C 에서 다소 높았으나, 온도에 따라 통계적으로 유의한 차이는 없었다(Fig. 3). 전보에 의하면 봄철 노지에서 재배되는 무의 경 우 광합성은 15-25°C 범위에서는 엽온이 증가함에 따라 광합성 속도가 증가하고, 25°C 이상의 온도에서는 감소하였으며, 광 합성을 위한 최적의 온도는 24°C이다(Oh et al., 2015). 그러나 월동무의 경우에는 재배기간 동안 Fig. 1B에서 살펴본 바와 같 이 터널 입구의 일평균 대기온도는 8.9°C로 낮고 일최고 온도도 20°C 이내에 해당되므로 보다 왕성한 광합성을 위해서는 겨 울철 대기온도보다 다소 높은 온도가 필요한 것으로 판단된다. 오이(Cucumis sativus L.)의 경우에도 15°C 이하의 온도에서는 증산률이 낮지만 15?30°C의 범위에서는 온도가 증가함에 따라 증산률이 높아졌으며(Eguchi and Koutaki, 1986), 광합성과 수분흡수도 24°C에서 최대값을 나타내지만 그 이하의 온도에서는 낮았다(Yoshida and Eguch, 1989). 그리고 토마토 (Lycopersicon esculentum cv. Burpee 'Big Boy Hybrid')의 생육에 있어서도 최적온도인 25°C에서는 측정된 모든 생장 반응이 저 온에서보다 좋을 뿐만 아니라 양분흡수도 가장 높은 것으로 알려지고 있다(Tindall et al., 1990). 따라서 겨울철 온도상승은 월 동무의 증산률, 양분과 수분 흡수율을 극대화하고 이로 인해 광합성과 생장이 증가하는 요인으로 작용할 것으로 보인다. 한 편, 대기조건에서보다 대기온도 +4°C와 대기온도 +7°C 조건에서 월동무의 수분이용효율이 감소하고 있는데, 이는 겨울철 기온상승이 월동무 재배시 수분 부족을 야기할 가능성을 나타내 주고 있다 .
Fig. 2.
Photosynthetic rate (A), stomatal conductance (B), transpiration rate (C), and water use efficiency (D) in radish leaves grown for 90 days under different temperatures in a temperature gradient tunnel. Each column and error bar represents the mean ± SE of five replicates. Different letters above bars indicate significant differences between temperature treatments (p < 0.05)
Fig. 3.
Photosynthetic light-response curves (A), leaf photosynthetic capacity (B), and apparent dark respiration rate (C) in radish leaves grown for 90 days at different temperatures in a temperature gradient tunnel. Each value represents the mean ± SE of five replicates. Different letters above bars indicate significant differences between temperature treatments (p < 0.05).
이상의 결과를 종합해 보았을 때 급격하게 변화하는 미래의 기후 환경 하에서 겨울철 온도의 상승은 월동무의 양분 흡수 를 촉진하고 광합성을 증가시켜 무의 생산성을 증가시킬 것으로 보인다. 그러나 지구 온난화로 인한 기후변화는 식물의 생 육과 농산물 생산의 양적인 측면에서 뿐만 아니라 질적인 측면에 있어서도 변화를 야기할 가능성이 있다. 그리고 따뜻해진 겨울로 인해 월동해충이 증가하고 돌발 병해충이 발생하여 이에 따른 작물의 피해도 나타날 수 있을 것이다. 따라서 겨울 철 온도 상승으로 인한 월동무의 질적 변화, 병충해 발생 등에 대한 좀 더 체계적인 작부 체계를 연구할 필요가 있을 것으로 보인다.
