

서 언
재료 및 방법
실험재료
단백질 추출 및 이차원 전기영동(Two-dimensional electrophoresis; 2-DE)
MALDI-TOF/TOF MS의 측정, 데이터의 분석 및 정리
결과 및 고찰
대사(Metabolism)관련 단백질
에너지(Energy)관련 단백질
서 언
블루베리는 진달래과(Ericaceae) 산앵두나무속(Vaccinium)에 속하는 관목성 식물로 각종 무기염류, 비타민류, 섬유질을 포함하고 있으며, 과실의 식용 시 폐기될 부분이 거의없으므로 다양한영양성분의 이용효율이 매우 높다(Kang et al., 2009). 블루베리는 anthocyanin, flavonoid, lycopene 등 같은 다양한 기능성 물질을 가지고 있으며이들 성분은 항산화기능, 시력강화, 면역력 증진및 뇌졸중방지 등에효과가 있는 것으로 알려져있다(Routray et al., 2011; Chung et al., 2016). 특히 블루베리의anthocyanin 은 활성산소와 노화에 의한 알츠하이머와 파키슨병 같은 퇴행성 질환에 대한 그 효과가 검증 되었고, 자외선에 의한피부 노화를 억제하고, 콜라겐 파괴, 염증 등을 지연시킨다는 연구결과도 있다(Bae et al., 2009). 또한, 지방세포를 사멸시켜 다이어트에 대한 효과가 있으며, 대장암, 유방암 및 자궁경부암 등에도 효과가 있음이 확인 되었다(Routray et al., 2011). 이러한 다양한 기능성을 인정받아 뉴욕타임지에서는 블루베리를 ‘세계10대 슈퍼 푸드’로 선정하였으며, 그 관심과 인기는 더욱 높아졌다.
프로테오믹스는 게놈(genome) 에서 만들어지는 단백질의 총체를 의미하는 프로테옴을 연구하는 방법과 기술을 포괄적으로의미하는 말로‘단백질체 분석학’이라고도 한다. 일반적인 과정은 추출된 단백질을2차원전기영동(2-DE; two-dimensional electrophoresis) 으로 전개한 후 각각의 단백질 spot 을 coring 한 다음 질량 분석을 통하여 동정하는 것으로, 프로테옴의 대량 분석과 상호기능관계 지도의 작성, 구조분석 등을 통해 궁극적으로 특정 단백질과 이를 만드는 유전자의 기능을 동시에 밝혀낼수 있다(Chen, 2006). 이와 같은 프로테오믹스의 방법과 기술을 이용하여 현재까지 포도, 망고, 파파야, 오렌지, 토마토 등을 비롯한 다양한과실에서 프로테옴의 분석이 실시되었다(Novikova et al., 2003; Palma et al., 2011; Zeng, et al., 2011; De Ma-galhães Andrade et al., 2012; Manaa et al., 2013; Wu et al., 2013). 특히 포도에서는 성숙단계 및 차광에 따른 과피의 프로테오믹스, ABA 처리에 다른 프로테옴의 변화 등다양한 보고가 있었으나(Deytieux et al., 2007; Marzia et al., 2010; Niu et al., 2013; Yuan et al., 2014), 블루베리의 프로테옴에 대한 분석은 아직까지 보고 된 바가 없다. 따라서, 본 연구에서는 래빗아이 블루베리인 ‘Bluegem’ 의 과피에서 발현되는 프로테옴을 분리·동정하기 위한 실험을 수행하였다.
재료및방법
실험재료
본 실험에서는 래빗아이 블루베리(Vaccinium ashei) ‘Bluegem’ 을 사용, 2012 년 8월에수확하여 냉장 보관된 과실 중 평균무게 2.1g, 평균 횡경 17.8mm, 종경13.1mm 인 과실의 과피를 과육과 분리하고 단백질 추출에 이용하였다.
단백질추출및 이차원전기영동(Two-dimensional electrophoresis; 2-DE)
단백질 추출은 과육과 분리된 블루베리 과피 2.5g 을 액체질소에 마쇄하여phenol 법으로 실시하였다(Hurkman et al., 1986). 추출한 단백질은 Bradford 법으로 정량하였다(Bradford, 1976). 이차원 전기영동은 Lee et al.(2012) 의 방법에 따라 실시되었다. 등전점(isoelectronic-focusing; IEF) 전기영동은 17cm immobilized pH gradient gel(IPG) strip(pH4-7, Bio-rad, CA) 에 단백질을 로딩하고PROTEAN IEF Cell(Bio-rad, CA) 에서 실시하였으며, SDS-PAGE 12% polyacrylamide gel(30% acrylamide/ 0.8% bis-acrylamide 16mL, 375mM Tris-HCl(pH = 8.8) 10mL, dH2O 13.6mL, 10% SDS 0.4mL, TEMED 20µL, 10% APS 80µL)을 이용하여PROTEAN® II xi Cell(Bio-rad, CA) 에서 실시하였다.
2-DE 가 끝난 gel 은 Coomassie Brilliant Blue G-250(CBBG-250, Bio-rad, CA) 를 이용하여 염색하였으며(Matsui et al., 1999). GS-800 Calibrated Densitometer(Bio-Rad) 를 사용하여 스캔하였다. 5번의 전기영동을 실시하였으며, 가장 고화질의이미지를 갖는gel을 선택하여master gel 로 정하고, 그 gel에서 육안으로 식별 가능한단백질 spot 의 각각에 고유 번호를 부여하였다.
MALDI-TOF/TOF MS 의 측정,데이터의 분석 및 정리
CBBG 로 염색된 gel에서 단백질 spot 을 coring 하고, 탈색, reduction, alkylation 및 In gel digestion 을 하였다(Kim et al., 2004). MALDI-TOF / TOF MS 분석은 355nm 에서 작동하는 200 Hz ND: YAG laser 를 사용하는ABI 4800 Plus TOF-TOF Mass Spectrometer(Applied Biosystems, Framingham, MA, USA) 로 실시하였다. MS / MS spectra 는 검색엔진으로서 MASCOT (http://www.matrixscience.com) 와 ProteinPilot v.3.0 을 사용하여NCBInr database(http://www.ncbi.nlm.nih.gov/) 에서 50ppm 의 peptide 와 fragment ion mass tolerance 로 검색되었으며, 동정된 단백질은 ‘GO-MIPS funcat conversion table’(http:// geneontology.org/external2go/mips2go) 의 기준에 따라 분류되었다.
결과및고찰
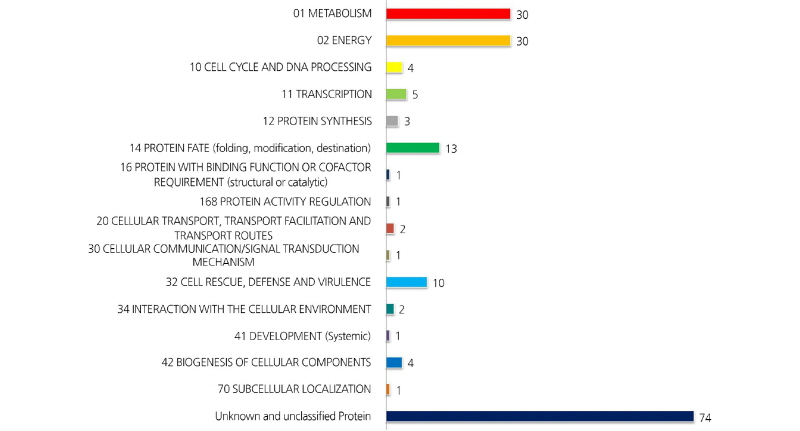
전개된 2-DE image 에서 최대810 개의 단백질 spot이 육안으로 확인되었다. 이중5반복의 실험 중 최소3개의 gel 에서 나타나고coring 이 가능한 247 개 단백질 spot 의 MALDI-TOF / TOF MS 분석을 실시하였다. 분석된 spot 중 score 값이45이하인것, signal peak 가 검출되지 않은 것 등을 제외하고, 182 개의 단백질 spot 을 NCBI database 에서 검색하였다. 검색된 단백질들은 아래와 같이‘GO-MIPS funcat conversion table’ 의 기준에 따라 분류하고 정리하였다(Figs. 1, 2). 검색된 182 개의 단백질spot 중 74개는 unknown 또는 unclassified 단백질, 대사(metabolism) 와 에너지 관련 단백질이 각각30개, 14 개 는 단백질fate (folding, modification, destination) 와 관련된 단백질, 10 개는cell rescue, defense 및 virulence 와 관련된 단백질로 나타났다(Table 1).

Fig. 1.
High-resolution 2-DE of total proteins extracted from the skin of rabbiteye blueberry (Vacciniumashei ‘Bluegem’).
(A) Metabolism-relatedproteins, (B) energy-relatedproteins, (C) proteinfate,cellrescue,defense, virulence-relatedpro-teins,and (D)miscellaneousproteins.Theproteinswereseparated on IPGstrips(pH 4-7),followedbySDS-PAGEon 12% polyacrylamidegels.Gels werestained withcolloidal coomassiebrilliant blueG-250. The spotnumbersindicateproteins that were identified in the skin of rabbiteye blueberry.
대사(Metabolism)관련 단백질
단백질spot 327, 344, 378, 379 및 456은 아미노산 대사와 관련된 glutamine synthetase 로 동정되었는데, 포도 과피단백질체분석에서도 착색이 진행됨에 따라 그 발현량이 증가하는 것으로 보고되었다(Deytieux et al., 2007).
Spot 390 과 799 는 nucleotide 대사와 관련된 adenosine kinase isoform2T 와 nucleotide diphosphatekinase 로 각각 동정되었다. Adenosine kinase isoform 2T 는 이전의 연구에서 토마토에 염과Ca 처리 시 발현량이 감소되는 것으로알려졌으며(Manaa et al., 2013), 동일 계통 고추의 세포질웅성불임 화분과 가임 화분을비교하면 세포질웅성불임 화분에서 그 발현량이 더낮은 것으로 보고되었다(Wu, et al., 2013). Nucleotide diphosphate kinase 는 파파야에서 성숙이 진행됨에 따라 발현량이 감소되는것으로 알려져 있다(Huerta-Ocampo et al., 2012).
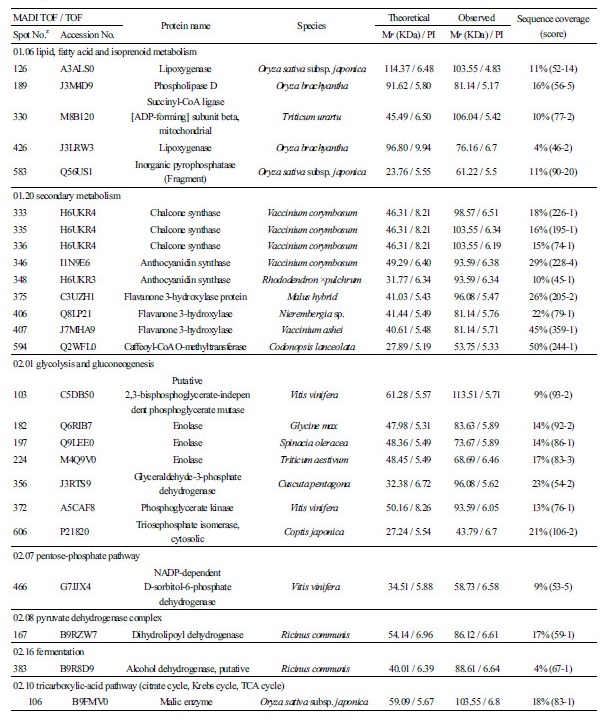
C-compound 및 carbohydrate 대사와 관련된 단백질들로써spot 74, 205, 316, 368 은 RuBisCO 로, spot 184 는 UDP-glucose pyrophosphorylase 로, spot 332, 724 는 formate dehydrogenase 로 각각 동정되었다. Lipid, fatty acid 및 isoprenoid 대사와 관련해서는 spots 126, 426 이 lipoxygenase 로, spot 189 가 phospholipase 로, spot 330 이 succinyl-CoA ligase 로, spot 583 이 inor-ganic pyrophosphatase 로 각각 동정되었다. Inorganic pyrophosphatase 는 토마토와 살구에서 과실이 성숙됨에 따라 그 발현량이 감소되는 것으로도 확인되었다(Rocco et al., 2006; D’Ambrosioa et al., 2013).
이차대사(Secondary metabolism) 와 관련해서는 anthocyanin 의 생합성을 조절하는 다수의효소단백질들이 동정되었는데, spot 333, 335, 336 이 chalcone synthase 로, spot 346, 348 이 anthocyanidin synthase 로, spot 375, 406, 407, 594 가 flavanone 3-hydroxylase 로, spot 202 가 caffeoyl-CoA o-methyltransferase 로 각각 동정되었다. 이전의 단백질체 관련 연구에서도 과실의 anthocyanin 생합성 경로에 관련된 다양한 효소 단백질들이 확인되었는데, 특히 포도에서cinnamate-4-hydroxylase, 4-coumarate-CoA ligase, chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, dihydroflavonol 4-reductase, an-thocyanidin synthase, UDPG-flavonoid-3-o-glucosyltransferase, anthocyanidin reductase, leucoanthocyanidin reductase, flavonol synthase 등 다양하게 확인되었다(Routray et al., 2011; Kayesh et al., 2013).
과실의 anthocyanin 생합성과 관련된 연구는 단백질체 연구 이전부터 효소활성과유전자 발현에 대하여 많은 연구가 되어왔다(Honda et al., 2002; Jaakola et al., 2002; Kim and Yun, 2015; Li et al., 2012; Zhang et al., 2017). Honda et al.(2002) 는 사과의 경우 과피에서 적색계통의anthocyanin 축적 증가와함께chalcone synthase(CHS), flavanone 3-hydroxylase(F3H), dihy-droflavonol 4-reductase(DFR), anthocyanidin synthase(ANS), UDP glucose: flavonoid 3-Oglucosyltransferase(UFGluT) 의 다섯 가지 유전자의발현이 증가한다고 보고했다. 블루베리에서도 항산화 물질 특히 anthocyanin 과 관련하여 유전자수준에서의 다양한 연구가 보고되었다. Li et al.(2012) 는 블루베리 ‘Northland’ 를 이용하여 Illumina 염기서열 분석을하여얻은 결과를통해de novo assemblies 를 수행34,364 개의unigen 을 생성하였고, 또한 블루베리 과실의 발달 과정에 따라anthocyanin 함량
과 관련 유전자의 발현 수준을 비교해 본 결과 파란색 과실의 과피에서 CHA 와 F3H 유전자의 발현수준이 높았다고 보고하였다. 또한 블루베리의 F3H 유전자의 기능분석에 대한연구에서도 F3H 을 포함한CHS, DFR, 그리고ANS anthocyanin 생합성과 관련된 유전자발현이 식물조직의anthocyanin 함량과 상관관계가 있음을 확인시켜주었다(Ziang et al., 2017). 동정된 단백질 중caffeoyl-CoA o-methyltransferase 는 리그닌생합성 경로에서 중요한 효소로, 포도에서는 적색과피보다 백색과피에서더 많이 발현되는 것으로 보고되었다(Niu et al., 2013). 본 연구에서는 블루베리 과실의 발달 단계에 따라 단백질체를 비교 분석하지는 않았지만, 과피에서 동정된전체 단백질체를 확인한 것으로추후 블루베리의 과실발달 단계별로 발현되는 유전자혹은 단백질, 효소활성 비교분석에 대한 연구에유용한 정보가 될 것으로 사료된다.
에너지(Energy)관련단백질
해당작용(glycolysis) 과 glucogenesis 관련해서는 spot 103 이 phosphoglycerate mutase 로, spot 182, 197, 224 가 enolase 로, spot 356 이 glyceraldehyde-3-phosphate dehydrogenase 로, spot 372 가 phosphoglycerate kinase 로, spot 606 이 triose phos-phate isomerase 로 각각 동정되었다. 토마토와 포도의 경우 성숙이 진행됨에 따라 해당작용과 관련된 효소들의 발현량이 감소하였지만, 살구에서는 오히려 반대의 경향을 나타내는 것으로 보고되었다(Rocco et al., 2006; D’Ambrosioa et al., 2013; Deytieux et al., 2013). 본 실험의경우블루베리 과피에서도 다양한해당작용 관련 효소가확인되었으므로, 과실 성숙 단계별로 각각의 발현량 변화를 구명할필요가 있을 것으로 사료된다.
오탄당 인산경로(pentose-phosphate pathway) 와 관련된 단백질로써는 spot 466 이 D-sorbitol-6-phosphate dehydrogenase 로 동정되었다. Pyruvate dehydrogenase complex 와 관련해서는 spot 167 이 dihydrolipoyl dehydrogenase 로 동정되었는데, 이 효소단백질은 포도의 착색기때ABA 를 처리하면 과육에서 발현량이 감소하는 것으로 알려졌다(Marzia et al., 2010).
발효(fermentation) 와 관련된 단백질로써spot 383 은 alcohol dehydrogenase 로 동정되었다. Alcohol dehydrogenase 는 포도 과피의 차광처리 시 그 발현이 감소되는 것으로 보고되었는데(Niu et al., 2013), 살구에서도 과실이 성숙함에 따라dihydroli-poyl dehydrogenase 와 더불어 alcohol dehydrogenase 의 발현이감소되는 것으로알려졌다(D’Ambrosioa et al., 2013).
TCA cycle 과 관련해서는 spot 322 가 isocitrate dehydrogenase 로, spot 90 이 succinate dehydrogenase 로, spot 106, 107 이
Table 1. Identification of proteins expressed in the skin of rabbiteye blueberry by MALDI-TOF / TOF MS
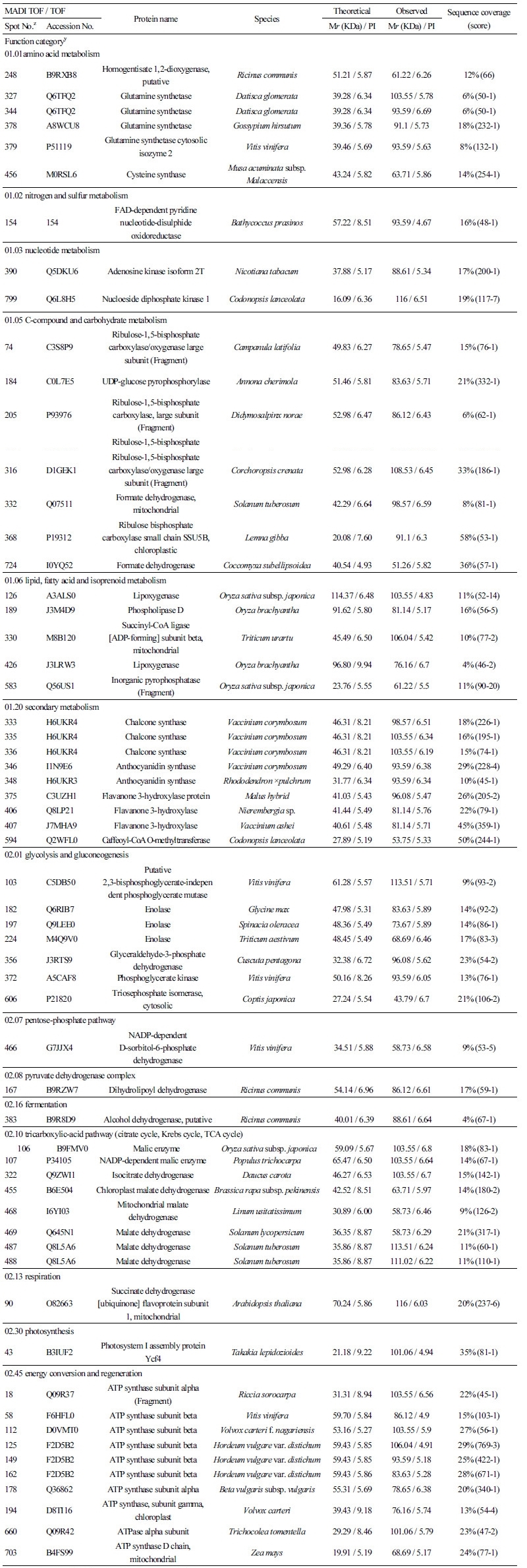 |
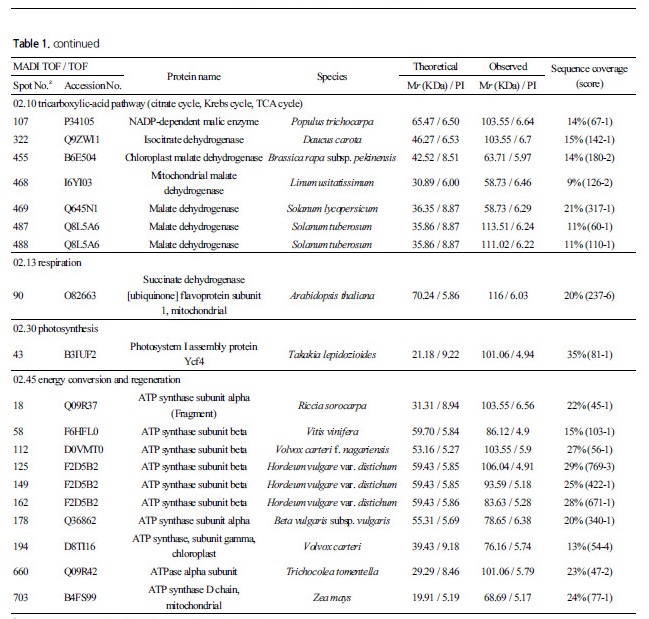
malic enzyme 으로, spot 455, 468, 467, 487, 488 이 malate dehydrogenase 로 각각 동정되었다. Isocitrate dehydrogenase 는 복숭아 과실의 성숙과 함께 그 발현량이증가하는 것으로보고되었다(Nilo et al., 2012). Succinate dehydrogenase 는 MIPS 분류에서02.13 인 호흡(respiration) 관련 단백질에 속하는 것으로 되어있지만, TCA cycle 과 관련된효소로 적색 과피의 포도 보다백색 과피의 포도에서 착색기 전에 더 많이 발현하는 것으로 보고되었다(Niu et al., 2013). Malic enzyme 은 malate dehydro-genase 로도 불리는데, 포도의 경우 착색기 때ABA 를 처리하면 과육에서 발현량이 증가하나, malic acid 의 함량은 착색기 전부터 이미 높은 상태로 그 함량의 변화가 없는 것으로 보고되었다(Marzia et al., 2010).
광합성의명반응계와 관련해서는 광계I의 생합성에 관여하는 단백질로 spot 34 가 Ycf4 로, spot 18, 58, 112, 125, 149, 162, 178, 194, 660, 703 가 ATP synthase 로 각각 동정되었다. 명반응계에서ATP synthase 는 엽록체의틸라코이드에 위치하며ADP 와 Pi를 ATP 로 전환하는 과정을 촉매화하는데, 다수의 ATP synthase 가 동정되는 것으로 보아 블루베리 과피에서도 다량의 ATP 가 생합성 되는 것으로 사료된다.
