

서 언
재료 및 방법
시험재료 및 수관위치 구분
광합성 특성 조사
잎과 과실의 특성 조사
잎과 과일의 조직학적 형태 조사
가용성 당 함량 분석
통계분석
결과 및 고찰
수관위치에 따른 투광률과 광합성 특성
수관위치에 따른 잎의 생장과 조직학적 특성
수관위치에 따른 과실 특성과 세포조직 및 당 조성
서 언
고품질 사과 다수확 기술에 관한 요구도가 높아짐에 따라 과수원의 자연환경 뿐만 아니라 수관내부에서의 미세 환경에 대한 연구가 많이 진행되고 있다(Jung, 2002; Kim et al., 2003; Oh et al., 1997; Park et al., 2007). 균일한 착과를 위해서는 무엇보다 수관내부 무효공간을 최대한 줄여야 한다. 하지만, 여름철 연중 강수량의 70%가 집중되는 고온 다습한 기후(Seo, 2010) 조건에서 습관적으로 적용하는 강한 절단전정과 지나친 시비(Kim et al., 1996)는 영양생장을 촉진 및 연장시켜 수관 내부의 무효공간을 만든다. 이러한 이유로 수관내부의 과실 착색이 지연되는 등 품질이 떨어지게 된다(Kim et al., 1996).
수관위치에 따른 과실 품질은 투광량에 의해 가장 많은 영향을 받으며, 투광량은 수관의 상부에서 하부로, 외부에서 내부로 그리고 수관이 복잡할수록 불량해진다(Park et al., 2007). 또한, 수관내부의 투광률이 17% 이하로 낮아지면 개화에 큰 지장을 주고, 부분 차광만으로도 그 부위의 개화를 억제시킨다(Auchter et al., 1926).
투광량의 차이는 잎의 책상조직이 차지하는 비율과 두께에 영향을 미친다(Cowart, 1935; Oh et al., 1997). 이러한 차이에 의해 광량이 적은 위치의 수체 조직이 연화되어 병 저항성이 약화되는 원인이 된다. 또한, 수관위치별 투광량은 과실의 무게와 정의 상관관계가 있으며, 과실 품질에 큰 영향을 미치는 요인이 된다. 뿐만 아니라, 광합성 효율에도 영향을 주어 과실 수확량에도 직접적인 영향을 미친다(Barritt et al., 1991; Jung, 2002).
이와 같이 수관 내 투광량의 중요성이 오래 전부터 강조되었지만(Auchter et al., 1926), 우리나라에서 육성 보급된 ‘홍로’ 사과나무의 투광량에 따른 수관위치별 과실 품질 및 조직학적 특성에 관한 연구는 부족한 실정이다.
이에 본 연구에서는 ‘홍로’/M.9 세장방추형 수형에서 수관내부의 무효공간을 줄이고 고품질 과실 생산을 위한 재배기술의 기초자료를 제공하기 위하여 미세환경이 다른 수관위치에서의 투광량, 광합성 특성, 잎과 과실의 특징 등을 구명하고자 수행하였다.
재료 및 방법
시험재료 및 수관위치 구분
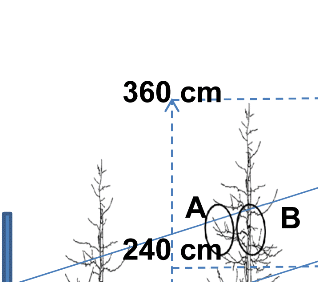
수관위치에 따른 광합성 특성, 잎의 생장, 과실품질 그리고 잎과 과실의 조직학적 분석을 위해 전북 장수군 송천리(고도 510m)에 위치한 참솔농장에서 열간거리 4m, 주간거리 2m로 재식된 수고가 3.6m 정도인 10년생 세장방추형 ‘홍로’/M.9 사과나무 중 표준이 될 수 있는 나무를 선정하여 광합성 특성은 6주를, 잎과 과실특성은 3주를 사용하였다.
수관위치를 Fig. 1과 같이 4지점(A, B, C, D)으로 구분하였는데, A는 수관 상단부(240cm 이상)의 주간으로부터 측지 길이의 1/2 바깥쪽 부분(수관상단-외부), B는 상단부의 1/2 안쪽 부분(수관상단-내부), C는 수관 하단부(240cm 이하)의 1/2 바깥쪽 부분(수관하단-외부), 그리고 D는 하단부의 1/2 안쪽 부분(수관하단-내부)을 각각 선정하였다.
광합성 특성 조사
수관위치에 따라 광합성속도, 증산속도, 기공전도도, 광도를 Fig. 1과 같이 수관위치별로 정상적으로 잎 전개가 완전하게 진행된 직후의 잎을 휴대용 광합성측정장치(LC pro+, ADC Bioscientific Ltd., England)를 이용하여 2011년 7월 초 맑은 날 오전(10시-12시)에 수관위치별로 6엽씩 측정하였다.
잎과 과실의 특성 조사
수관위치에 따른 잎의 크기는 2011년 신초생육이 정지되는 8월초에 신초의 기부에서 3-5번째 잎을 위치별로 주당 10엽씩 3주에서 조사하였다. 수관위치에 따른 과실 품질 특성은 2011년 적숙기인 9월 10일에 수관위치별로 주당 과실 10개씩 3주에서 총 30개의 과실을 수확하여 조사하였다. 과경은 종경과 횡경의 최대 부위를 측정하고 과형지수를 계산하였다. 과피색은 색차계(300R, Minolta, Japan)를 이용하여 측정하였는데, L 값은 0(black) - 90(light)까지의 값으로 표시하고, a 값은 -60(green) - +60(red)으로, b 값은 -60(blue) - +60(yellow)으로 표시하였다. 경도는 지름 8mm 탐침을 가진 과실경도기(fruit texture analyser, Korea)를 사용하여 과피를 제거하지 않고 측정하였다. 과실 당도는 비파괴 디지털 당도계(AMAICA, Astem Co., Japan)를 이용하였으며, 산도는 Whatman No. 2 여과지에 여과한 과즙을 5mL 채취하여 증류수 50mL를 섞은 후, 1% phenolphtalein을 지시약으로 0.1N NaOH로 적정하여 변색점을 종말점으로 하여 malic acid 상당량으로 환산하였다.
잎과 과일의 조직학적 형태 조사
수관위치에 따른 잎 단면과 과피의 조직 검경을 위해 햇빛을 가장 잘 받는 수관외부(수관위치 A)와 차광이 가장 많이 되는 수관내부(수관위치 D)로 구분하여 잎은 2011년 8월초 기부에서 3-5번째 잎을 채취하였고, 과실은 9월 10일 적숙기에 수확하여 실험 재료로 사용하였다. 현미경 검경은 Luft(1973)의 방법을 따라서 epon block을 만들었고, P.A.S. 염색법으로 염색하여, 광학현미경(Axioskop 2, Carl Zeiss Co., Germany)으로 엽조직은 200배율, 과육조직은 100배율로 검경하였다. 잎 단면의 책상조직 길이를 비교하기 위해 검경된 이미지를 Photoshop 프로그램을 이용하여 조사하였다. 과실의 표피 세포의 크기와 과피의 두께는 타닌 세포가 있는 층까지의 범위를 과피로 구분(Lim et al., 1993)하여 두께와 단위면적당(1,000µm2) 세포의 수를 조사하였다.
가용성 당 함량 분석
2011년 적숙기인 9월 10일에 수확된 과실의 가용성 당 함량을 조사하였다. 과실은 3개씩 착즙하여 원심분리한 다음 0.45µm syringe filter로 여과한 후 refractive index detector가 장착된 HPLC(1050, Agilent, USA)로 분석하였다. Column은 Zorbax carbohydrate(4.6 × 150mm, 5µm)를 사용하였고, 이동상은 water:acetonitrile = 20:80(v/v)으로 1.0mL・min-1의 속도로 흘려 보냈다. 당 표준용액은 rhamnose, xylose, sucrose, D-(+)-glucose, D-(-)-fructose, maltose를 증류수에 혼합하여 0.0625-0.5% 농도 범위로 준비하였다. 각 표준용액에 대한 검량곡선을 작성하였으며, peak의 면적으로 개별 당 성분의 함량을 %로 산출하였다.
통계분석
통계분석은 Windows용 SAS system, release 8.01(SAS Institute Inc, Cary, NC, USA)를 이용하여 수행하였으며, 유의수준 5%에서 Duncan 다중검정(DMRT)을 실시하였다.
결과 및 고찰
수관위치에 따른 투광률과 광합성 특성
2011년 7월 초 오전 10시부터 12시경 수관위치별 평균 광도는 수관상단-외부가 1,688µmol・m-2・s-1로 가장 높았고, 수관하단-내부가175µmol・m-2・s-1로 가장 낮은 것으로 나타났다. 또한 수관위치에 따른 광도는 위쪽이 아래쪽에 비해 높았고, 외부가 내부에 비해 높은 것으로 조사되었다(Table 1). 수세가 보통인 사과나무에서 수형과 품종에 상관없이 수관 상단부에서 하단부로 내려갈수록 일조량이 감소한다는 Forshey(2000)의 결과와 동일한 경향을 보였다.
수관상단-외부의 광합성 속도는 15.8µmol・m-2・s-1로 가장 활발하였으며, 수관하단-내부(3.5µmol・m-2・s-1)에 비해 약 4.5배 높은 것으로 나타났다(Table 1). 사과나무에서 수관위치별 광합성 속도는 투입되는 광도의 차이와 관련이 있으며, 보통 사과나무의 수관외부가 수관내부보다 3-4배 정도 높다고 하였다(Mika and Antosqewski, 1972).
수관위치에 따른 증산속도와 기공전도도는 각각 수관상단-외부가 5.01mmol・m-2・s-1, 0.19mol・m-2・s-1로 가장 높은 결과를 보여 광합성속도와 기공전도도 간에는 정의 상관을 갖는다는 Giuliani et al.(1997)의 보고와 유사한 경향을 보였다.
수관위치에 따른 잎의 생장과 조직학적 특성
수관위치에 따른 잎의 생장특성을 조사한 결과는 Table 2와 같다. 수관위치별 엽장은 수관외부의 것이 더 길고, 엽폭은 수관내부의 것이 더 넓은 경향을 보였다. 잎의 두께는 수관하단-내부의 잎이 0.195µm로 유의적으로 가장 얇았고, 다른 위치에서는 유사하였다.
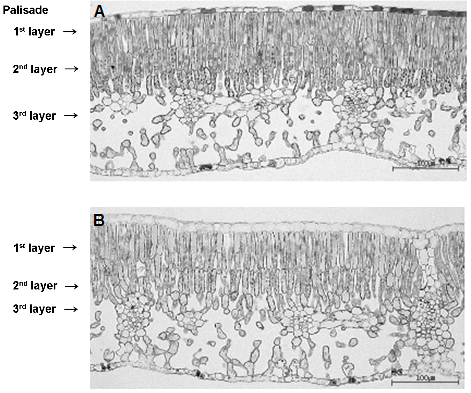
수관위치에 따른 엽육조직의 차이를 조사하고자 수관외부와 수관내부로 나눠 광학현미경으로 검경하였다(Fig. 3). 수관외부의 잎은 큐티클층의 두께가 두껍고, 책상조직의 세포 크기가 크고 치밀하게 정렬되어 있었다(Fig. 3A). 이에 반해, 수관내부의 잎은 책상조직 세포들이 엉성하고 불규칙적으로 정렬되어 있으며, 특히, 3층의 책상조직 중 가장 안쪽 책상조직의 발달이 매우 약한 것을 관찰할 수 있었다(Fig. 3B). 수관외부 잎의 1층, 2층 책상조직의 두께는 각각 110.4, 72.8µm이었고, 수관내부 잎은 68.5, 51.3µm로 햇빛을 적게 받는 내부 잎이 현저히 얇은 것을 알 수 있었다. 하지만, 잎 두께에 대한 전체 책상조직이 차지하는 비율은 외부 잎에서 36.4%, 내부 잎에서 35.2%로 큰 차이는 없는 것으로 나타났다(Table 3). Oh et al.(1997)은 ‘후지’ 양광면 잎의 책상조직의 비율은 60%로 음광면 잎의 44%보다 현저히 높았지만, ‘조나골드’의 경우 거의 차이가 없어 잎에서 책상조직이 차지하는 비율은 품종에 따라 다르다고 하였다. 위의 연구결과와 비교해 볼 때 ‘홍로’는 ‘후지’나 ‘조나골드’에 비해서 광부족에 의한 엽조직의 반응이 덜 민감한 것으로 판단되었다. 사과의 책상조직과 잎의 두께는 광도와 직접적인 상관이 있으며, 이러한 현상은 사과는 물론 열대 야자나무와 같은 다른 작물의 경우에서도 동일하게 나타나고 있다(Doud and Ferree, 1980; Reyes et al., 1996; Wooge and Barden, 1987). 또한, 광도가 감소되면 엽중이 가장 영향을 많이 받아 그늘에서 자란 잎이 양광에서 자란 것보다 가볍다고 하였다(Oh et al., 1997; Wooge and Barden, 1987). 이러한 조직학적 구성 차이에 의해 광량이 적은 수관 위치의 수체조직은 연화되어 병 저항성이 약화되며, 또한 영양생장이 촉진되어 신초의 길이가 길어지는 것으로 생각된다.
수관위치에 따른 과실 특성과 세포조직 및 당 조성
수관위치에 따른 과실 특성 조사 결과(Tabel 4), 과중은 수관상부의 과실이 더 무거웠으며, 수관외부가 내부보다 무거웠다. 가용성 고형물 함량은 수관상단-외부의 과실이 가장 높았고, 수관하단-내부가 유의적으로 낮았으며, 경도는 수관하단-내부의 과실이 가장 높았다. 그리고 산 함량과 과형지수는 차이가 없었다.
수관위치별 과실의 착색정도를 조사한 결과(Table 4), Hunter a 값은 수관상단-외부의 과실이 38.8로 가장 높았고, 수관하단-내부의 과실은 26.4로 유의적으로 가장 낮게 나타났다.
수관위치별 적숙기 착색의 진행 정도를 Fig. 2와 같이 시각화하여 비교해 본 결과 시기별 착색도가 큰 차이를 보였다. 수관상단-외부 과실의 착색이 30% 정도 진행된 8월 20일경에 수관하단-내부 과실의 착색은 전혀 진행되지 않았음을 관찰할 수 있었다. 이러한 결과는 수관 내 광환경에 의한 차이로 판단되며 투광량이 높은 수관상단-외부는 품질이 좋았으나 이에 비해 투광량이 적은 수관하단-내부 과실은 품질이 떨어지는 것으로 나타났다. 이와 관련하여 Han and Yoon(2001)은 광투과율과 과실의 무게는 정의 상관을 보여 햇빛을 많이 받는 부위에 결실된 과실일수록 과중이 증가하고, 가용성 고형물 함량은 높았으나 산 함량은 투광률과 관계가 없다고 하였다. 그리고 과피의 착색에 있어서는 수관내 광조건이 좋을수록 Hunter a 값이 증가한다고 하였다(Yang, 2008). ‘Delicious’ 사과의 경우도 수관내 광투과율이 30% 이하로 떨어졌을 때 화아 발달, 과실 크기, 과실 품질 및 생산성이 현저히 떨어졌다(Barritt et al., 1991). 따라서 이러한 수관위치별 투광률에 따른 현저한 과실 품질 차이는 내부 조직 수준까지 영향을 미칠 것으로 생각된다.
수관위치에 따른 과실의 유리당 함량(Table 5)은 수관상단-외부가 10.04%로 가장 높았고, 수관하단-내부가 8.36%로 가장 낮았다. 가용성 당 중 glucose는 수관위치별로 차이가 가장 많이 나타났는데, 수관상단-외부(2.38%)가 수관하단-내부(1.71%)보다 높았다. 반면, fructose과 xylose는 수관위치별로 유의한 차이가 나타나지 않았고, sucrose는 수관하단-내부가 1.76%로 다른 수관위치에 비해서 낮았다. 사과의 당도는 발육 초기에는 낮으나 후기에 갈수록 점차 증가하여 성숙기에 이르면 유과기의 약 5배 이상이 된다(Beruter, 1985; Song et al., 2003). 보통 사과의 당 조성은 성숙기에 과당 45-60%, 자당 20-40%, 포도당 20% 내외, 솔비톨이 3-8%를 차지한다(Pavel and Dejong, 1995; Yamada et al., 1994). 과실의 당도 및 당 조성의 차이는 품종 자체의 유전적 특징으로 결정되기도 하나 재배 환경에 의해서도 큰 차이를 보이는 것으로 알려져 있다(Ackermann et al., 1992). 이러한 결과는 가용성 당의 농도가 성숙기 이후에 급속한 증가 양상을 보이며, 성숙 초기에는 과당이 증가하고 후기에는 자당이 증가한다는 Song et al.(2003)의 보고로 볼 때 수관내부 과실은 동일한 시점에 성숙이 늦어진다는 것을 알 수 있었다.




















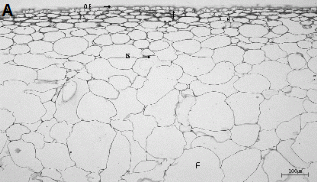
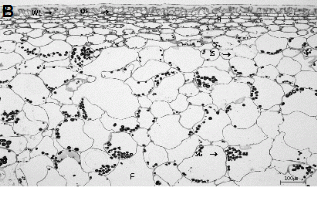
수관위치에 따른 과실 표피층 조직을 관찰한 결과(Fig. 4), 수관내부 과실은 현저히 왁스층이 발달되어 있으며, 외표피의 두께가 수관외부 과실에 비해 두꺼웠다. 또한, 아표피층 세포도 수관내부 과실이 외부 과실에 비해 조밀하면서 두꺼웠다(Fig. 4A).
수관외부와 내부 과실 모두의 외표피와 아표피층의 일부 세포들에서 떫은 맛을 느끼게 하는 탄닌이 관찰되었지만, 과육 세포에서는 전혀 나타나지 않았다. 수관내부의 과실 조직에서는 전분이 아직 당으로 전환되지 못한 상태로 남아있는 것을 관찰하였다(Fig. 4B). 이러한 점으로 볼 때 수관내부에 착과된 과실은 차광으로 인해 아직 적숙기에 도달하지 못한 상태이므로 수관외부의 과실과 동시에 수확하면 안 되고, 이때 수확하면 품질이 낮아질 것으로 판단되었다.
과실의 과피 두께를 조사한 결과(Table 6), 수관내부 과실은 133.3µm이었고, 수관외부 과실은 83.3µm로 조사되었다. 또한, 단위면적(1,000µm2)당 과피의 세포 수는 수관내부 과실이 3.63개, 수관외부 과실이 2.57개로 조사되어 수관내부 과실의 과피 세포가 외부보다 크기가 작고 조밀하게 구성되어 있었다.
이상의 결과를 종합해 보면, 사과나무의 수관위치에 따른 잎과 과실의 특성은 투광량에 의해 영향을 받았는데, 투광량이 적은 곳에서는 잎이 얇아지고 켜졌으며, 과실은 숙기가 지연되었다. 따라서, 수령과 수형 그리고 관리방법에 따라 달라지는 수관내부의 광환경을 개선해 주기 위한 지속적 노력과 수관 부위별로 숙기에 따라 분할수확이 필요하다고 생각된다.
