

서 언
2020년 기준 우리나라의 토마토 재배면적은 5,521ha로 그 중 수경재배 면적을 제외한 토경재배 면적은 4,939ha이다(KOSIS, 2021). 토마토 수경재배 면적은 매년 증가하고 있지만(RDA, 2018) 아직 토경재배 농가의 비율이 89%로 여전히 높은 실정이다. 토경재배 토마토는 주로 생식용으로 생산되고 7단 이하의 저단 화방까지 수확하는 단기재배로 이루어져 완숙토마토보다 방울토마토 재배가 선호된다. 그러나 토경재배 시설의 토마토는 연작으로 인해 염류장해와 토양전염성 병의 피해를 입기 쉬우며 겨울철 저온 피해에도 취약하여 결과적으로 생산량 감소로 인한 경제적 손실을 초래할 수 있다. 대목은 항상성을 유지하여 염류 스트레스 피해를 저감시킬 수 있고(Orosco-Alcala et al., 2021) 생물적 스트레스를 유발하는 토양전염성 병원균이 접수로 이동하는 것을 막을 수 있으며(King et al., 2008) 낮은 지온에서도 양수분 흡수율을 증가시켜 총 수량을 유지할 수 있다(Gálvez et al., 2021). 특히 풋마름병원균과 같은 토양전염성 병원균은 토양 내에 수년간 생존 가능하기 때문에 화학적 또는 생물학적 방제가 어렵다. 이러한 피해를 저감시키기 위해서는 토마토 대목 품종을 개발하여 접목묘를 사용하는 것이 최선의 방법이다. 국내에서 토마토 접목에 사용되는 대목은 주로 외국에서 개발된 품종으로 비용이 높은 단점이 있어, 대목 종자값 등의 초기 투자 비용은 농민에게 부담이 될 수 있다. 토마토 육묘과정에 투입되는 비용을 줄이기 위해서는 종자 비용 절감을 위한 국산 대목 품종의 개발과 성능 분석이 필요하다.
토마토 접목묘는 생물적·비생물적 스트레스를 극복하기 위해 사용될 뿐 아니라 생육 후기까지 세력을 유지할 수 있는 능력이 있어 많은 수량을 얻기 위해서도 사용된다(Stanghellini et al., 2019). 대목의 뿌리 활력은 양수분 흡수 효율을 높여 잎 등 영양기관을 발달시키고, 이에 따라 광합성 효율이 증가되어 수량과 수확 기간이 늘어난다(Lee and Oda, 2003). 토마토 접목묘를 사용하면 영양생장이 활발한 생육 초기의 광합성 능력이 높아지고 상대생장률이 증가하며 뿌리 발달이 촉진되어 총 수량이 증대된다(Latifah et al., 2021). 토마토의 생장과 발달에 중요한 요인인 온도는 발달속도에 따라 동화산물을 분배하는데 큰 영향을 미친다(Ploeg and Heuvelink, 2005). 토마토는 아열대성 원예작물로 12°C 이하가 되면 생장이 멈춘다(Criddle et al., 1997; Ploeg and Heuvelink, 2005). 12°C 이하의 낮은 근권부 온도는 뿌리 생장에 영향을 주는데 대목은 낮은 지온 조건에서도 양수분 흡수가 원활하게 이루어져 저온에 내성을 가진다(Schwarz et al., 2010).
엽록소형광 기법은 스트레스에 의한 광합성 변화를 비파괴적으로 대량 분석할 수 있어 생물적·비생물적 스트레스에 따른 식물생장 변화를 분석하기 위해 자주 활용된다(Oh and Koh, 2005). 엽록소형광 기법에는 Quenching, Kautsky effect(chlorophyll a fluorescence induction), JIP test(chlorophyll fluorescence induction curve) 등이 있다. 그 중 JIP test는 환경조건과 생육상태에 따라 변화하는 광계Ⅱ의 반응을 조사하는데 적합하다(Kalaji et al., 2011). JIP test는 엽록소가 암상태를 거치고 원래의 상태로 돌아오는 형광유발 시점(Origin) 이후 최소 형광변동 지점(Jump)을 지나 형광 중간수준(Intermediate)에서 최대 형광점에서의 형광유도량(Peak)을 표현하는 방식으로 OJIP(Origin Jump Intermediate Peak)라고 부른다(Govindjee, 1995; Yoo et al., 2013). OJIP는 광계Ⅱ의 광화학적 활성상태를 알아내기에 적합하고(Schansker et al., 2006), 엽록소형광 지수는 단독으로 측정되는 것보다 OJIP 단계별로 측정된 값이 그 활성상태를 더 정확하게 설명한다(Force et al., 2003; Živčák et al., 2008). OJIP 지수는 광계Ⅱ 전자전달의 순차적 흐름에 따라 정량화된 것이며(Baghbani et al., 2019) F0, FM, FV/FM, PIABS 및 ET0/RC 등이 있다. OJIP를 측정하여 대목의 종류별 광합성 효율을 객관적인 수치를 통해 평가 가능할 것으로 판단된다.
따라서 본 연구는 토경재배 시 방울토마토 대목 종류에 따라 나타나는 생육과 수량의 차이를 비교 분석하고 엽록소형광으로 측정한 광합성 효율을 평가하여 국내 개발 대목의 성능을 검정하고자 수행하였다.
재료 및 방법
실험재료
대목 품종 및 계통 4종류 ‘Powerguard’(National Institute of Horticultural and Herbal Science, Korea), ‘IT173773’ (National Institute of Horticultural and Herbal Science, Korea), ‘20LM’(Bunongseed, Korea), ‘B-blocking’(Takii, Japan)과 방울토마토(Solanum lycopersicum var. cerasiforme) 접수 품종 1종류 ‘Nonari’(Namnong, Korea)를 사용하여 접목한 처리구 4개와 접목하지 않은 처리구 1개를 실험에 사용하였다. 2020년 9월 3일에 접목할 대목 및 접수 종자를 128구 트레이에 파종하였고, 파종 후 18일에 대목묘는 40구 트레이에 가식하였다. 파종 후 25일에 합접법을 이용하여 접목하였다. 접목 직후, 온도 26°C 및 습도 80% 이상을 유지한 접목 활착실에 6일간 두었다. 실생묘는 접목묘와 같은 식물체 크기로 정식하기 위해 9월 10일에 40구 트레이에 파종하여 육묘하였다.
식물재배 및 재배환경
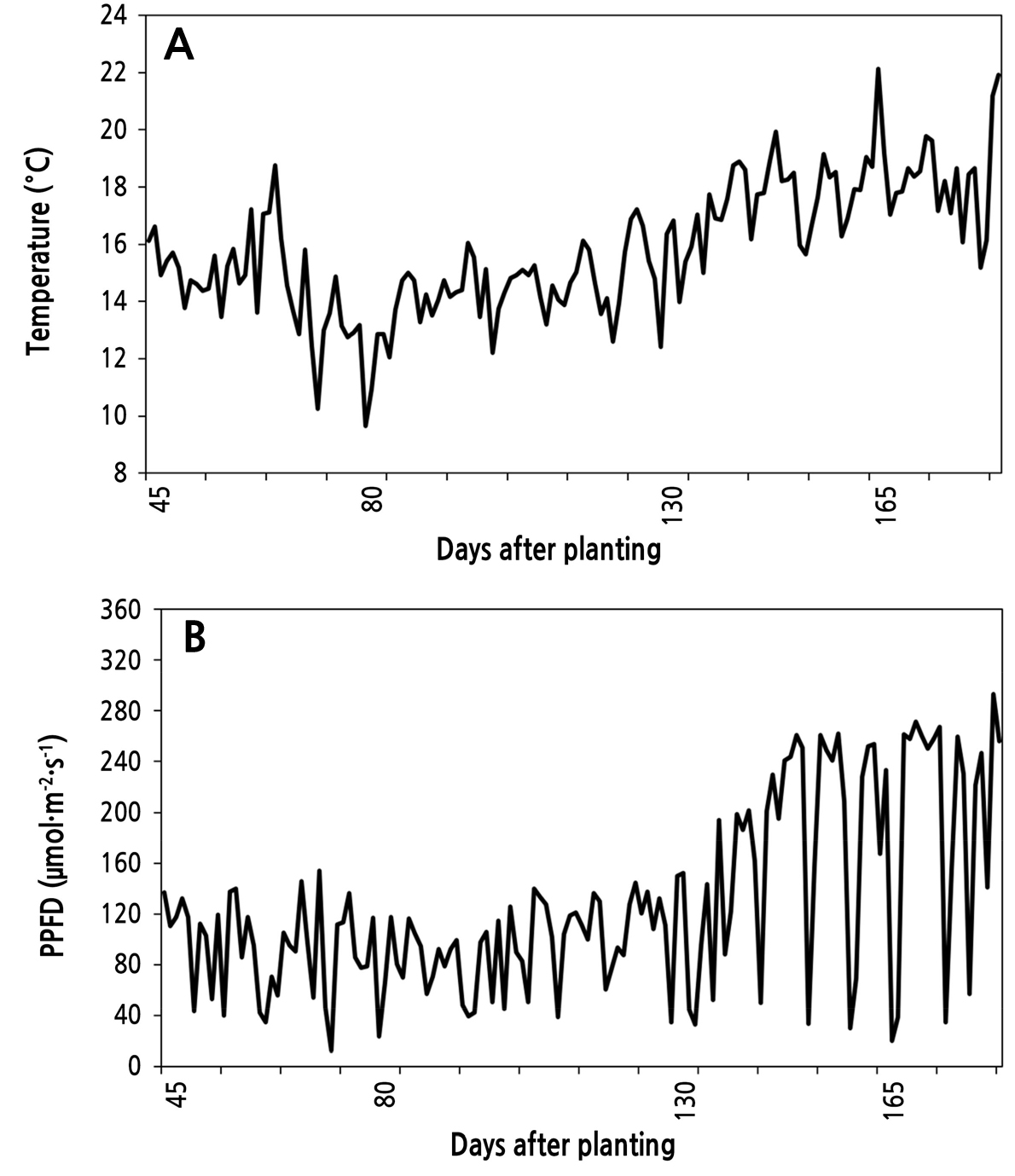
본 실험은 연동형 비닐하우스(폭 7m, 길이 72m, 동고 5.5m; 전북 정읍 소재)에서 토경재배로 수행했다. 접목처리구 4개와 비접목처리구 1개를 15주씩 난괴법 3반복으로 재식거리 35cm × 60cm로 하여 2020년 10월 20일에 정식하였다. 정식 전 유기질 비료(유기물 70% 이상, 수분 15% 이하, N-P-K 합 7% 이상)를 10a 당 15포 살포하고 경운하여 기비로 시용하였다. 생육 초기인 10월과 11월에는 N-P-K:30-10-10의 액비를 1주일에 1회 관수하고 생육 중기인 12월과 1월에는 N-P-K:19-19-19의 액비를 2주일에 1회 관수하며 수확기인 2월, 3월 및 4월에는 N-P-K:10-10-30의 액비를 1주일에 1회 관수하였다. 본 포장은 사양토로, 토양수분센서(SMEC 300, Spectrum, USA)를 사용하여 토양 수분함량 20% VWC 내외를 유지하도록 관리하였다. 생육 초기와 후기의 토양 EC는 5.8dS·m-1와 5.2dS·m-1로, 토양과 증류수를 1:5 비율로 하여 30분간 진탕한 후 측정하였다. 식물체 관리는 정식 후 측지를 주기적으로 제거하고 화방 내 꽃은 적화하지 않았으며 정식 후 93일에 7단에서 적심하였다. 온실 내부 온도와 일사량은 HOBO data logger(MX1104, Onset, USA)를 이용하여 수집하였다. 환경데이터 수집은 정식 후 43일(2020년 12월 1일)에 시작하여 정식 후 184일(2021년 4월 21일)까지 15분 마다 측정하였으며, 온도는 일평균으로 계산하였고 일사량은 일출부터 일몰까지의 값을 평균하여 그래프로 나타내었다(Fig. 1). 일사량은 lux 단위로 측정되어 환산계수 54로 측정값을 나누어 PPFD(µmol·s-1·m-2) 단위로 나타내었다(Thimijan and Heins, 1983).
생육조사 항목 및 측정방법
토마토 생육 비교를 위해 정식 후 59일(2020년 12월 17일)부터 정식 후 87일(2021년 1월 14일)까지 5주 동안 생장강도, 개화위치, 초장, 엽장 및 엽폭을 조사했다. 생장강도는 7일 전 생장점이 있던 위치를 표시한 유인줄의 줄기직경을 측정하였으며 측정할 부위가 마디라면 마디 바로 아랫부분을 측정하였다(Kim et al., 2010). 개화위치는 한 개 이상의 만개한 꽃이 있는 최상위 화방과 생장점 사이 거리를 측정하였다. 초장은 근권부부터 생장점까지의 거리를 측정하였다. 엽장과 엽폭은 개화한 최상위 화방의 하부엽 전체의 가로와 세로 길이를 측정하였다.
수량 및 과실특성 조사방법
토마토 수량은 정식 후 114일(2021년 2월 8일)부터 정식 후 165일(4월 2일)까지 1주일 간격으로 처리당 15주를 조사하였다. 수확한 토마토는 개체별로 개수, 평균 과중, 과경(과실 세로길이) 및 과폭(과실 가로길이)을 측정하였다. 과중은 전자저울(KS-308, Dretec, Japan)로 조사하였으며 과경과 과폭은 디지털 버니어 캘리퍼스 300mm(IP67, Futuro, Switzerland)를 이용하여 처리별로 과실 5개를 1회씩 조사하였다.
OJIP 측정 방법 및 지수
OJIP는 정식 후 45일, 80일, 130일 및 165일에 엽록소형광 측정기(Fluorpen FP-110, Photon Systems Instruments, Czech Republic)를 이용하여 개화한 최상위 화방의 하부엽의 맨 끝부분에 있는 잎을 대상으로 측정하였다. 잎은 leaf clip을 사용하여 1시간 암처리 후 처리별로 7개체를 1회씩 측정하였다. OJIP 측정 후 FluorPen(Photon Systems Instruments, 1.1.0.3 Version, Czech Republic) 프로그램을 통해 산출된 OJIP 지수 F0, FM, FV/FM, PIABS 및 ET0/RC 등을 사용하였고 각 지수를 표에 정의하였다(Table 1). F0는 엽록소분자들이 광계Ⅱ 반응중심으로 전달되기 전 방출하는 에너지의 정도를 나타낸 지수로, 값의 증가는 불활성 상태의 엽록소분자가 많아져 에너지 포획능력이 감소한 것을 의미한다(Baker and Rosenqvist, 2004). FV/FM은 엽록소형광 분석 연구에서 가장 많이 쓰이는 지수로 광계Ⅱ 최대 광화학적 효율을 나타내며 변동 형광 값(FV = FM–F0)과 최대 형광 값(FM)에 기인하여 계산된다(Yoo, 2012). PIABS는 광계Ⅱ의 광에너지 흡수 능력, 전자전달효율 및 전자고정효율의 세 가지 광합성 활성 정도를 통합적으로 나타낸 지수로(Thach et al., 2007) 광합성 기구의 전반적인 활력도를 나타낸다(Strasser et al., 2000; Živčák et al., 2008). ET0/RC는 QA를 QA-로 환원시키는 광계Ⅱ의 전자전달효율을 나타내는 지수로 스트레스를 받으면 값은 감소한다(Yoo et al., 2013).
Table 1.
Definitions of parameters obtained from the recorded chlorophyll fluorescence OJIP (Origin Jump Intermediate Peak) transients
통계분석
통계분석은 R(RStudio, 2022.02.0 Version, USA)을 이용하여 평균 간 비교를 Duncan’s multiple range test(DMRT) 검정을 통해 95% 신뢰수준에서 차이를 확인하였다.
결과 및 고찰
주당 평균 누적 수량은 ‘20LM’ 처리구가 1,339g으로 가장 많았고 ‘B-blocking’(1,142g), ‘Powerguard’(1,013g), ‘IT173773’(997g) 순이었으며 비접목처리구는 988g으로 가장 적었다(Fig. 2). 수확 초기의 비접목처리구는 높은 수량성을 보이다가 정식 후 144일부터 수량이 감소하기 시작하였고 동시에 ‘20LM’은 수량이 증가하였다. ‘20LM’의 주당 평균 누적 수량은 정식 후 144일(4화방)에서 정식 후 165일(7화방)까지 705g에서 1339g이 되어 90%의 증가율을 보였다. ‘IT173773’은 수확 초기부터 처리구 중 수량이 가장 적었고 수확 후기까지 증가 추세를 보이지 못하였다. ‘20LM’은 상업용 대목 품종인 ‘B-blocking’보다 주당 평균 총 수량이 197g 높아 수량성이 높은 것으로 판단된다.
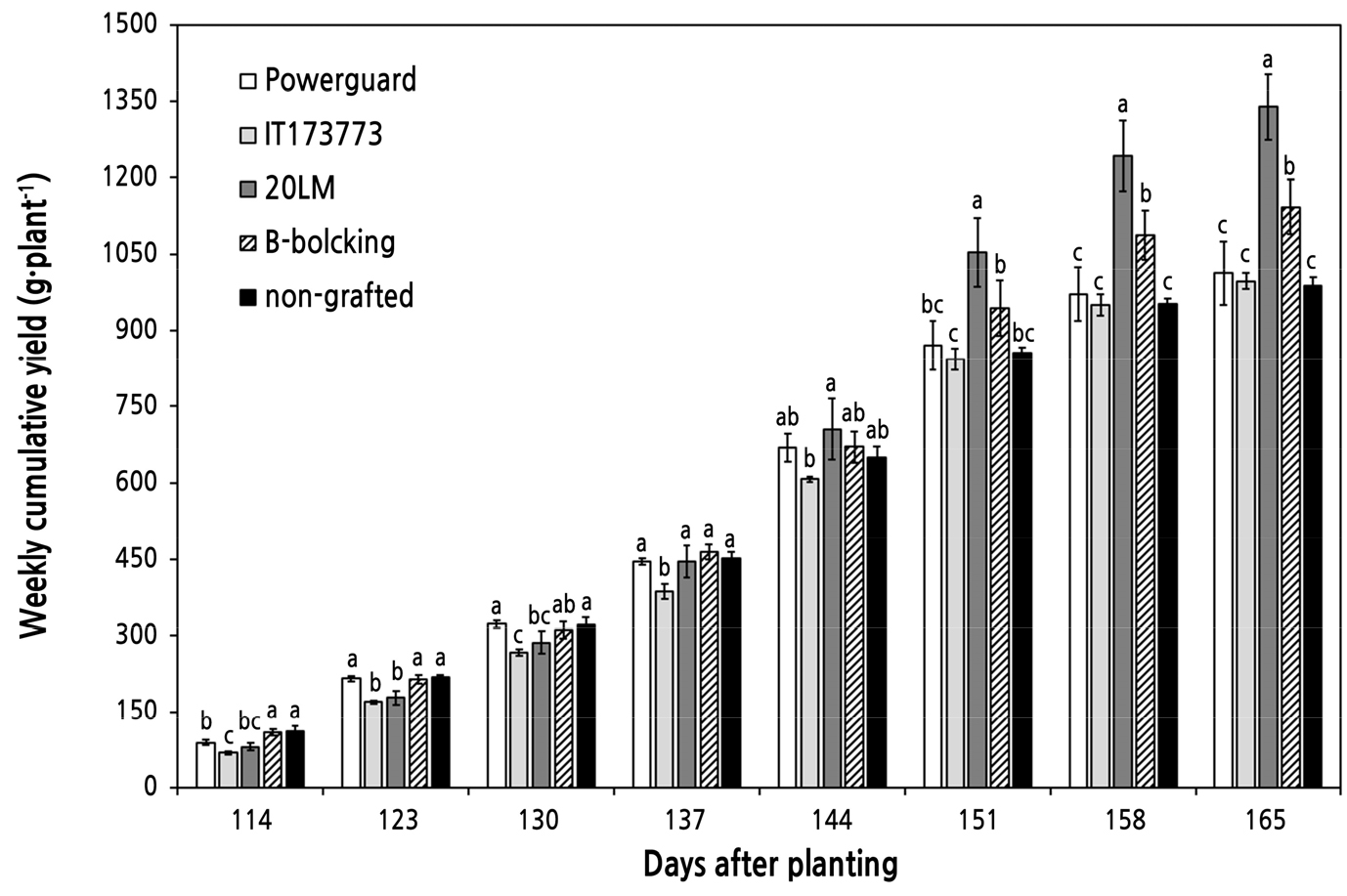
Fig. 2.
Weekly cumulative fruit yield of non-grafted and grafted cherry tomatoes. Four tomato cultivars, ‘Powerguard’, ‘IT173773’, ‘20LM’, and ‘B-blocking’, were used as rootstocks in soil cultivation. Different letters above bars indicate significant differences by Duncan’s multiple range test at p < 0.05.
‘20LM’의 평균 과중은 14.5g으로 가장 높았고, ‘Powerguard’, ‘IT173773’ 및 비접목처리구의 평균 과중은 각각 11.3g, 11.7g 및 11.6g으로 낮았다(Table 2). 과실 수는 처리 간 유의한 차이를 보이지 않았다. 정식 후 158일에 ‘20LM’의 과경과 과폭은 각각 44.7mm와 28.3mm로 높았다. 본 실험을 수행한 포장은 EC 5.2dS·m-1 이상의 염류집적 토양으로 간주된다. 염류 스트레스는 작물의 양수분 흡수를 방해하여 수분포텐셜이 감소됨에 따라 기공이 닫히고 광합성이 저해되는 문제를 일으킨다(Zribi et al., 2008). 하지만 접목묘는 염류농도가 높은 토양조건에서도 수분 이용 효율이 높으며 항산화효소의 활성이 높아 생육 저하와 광합성 저해 등의 피해를 저감시키는 이점이 있어(He et al., 2009) 본 실험에서 모든 접목처리구가 비접목처리구보다 수량성이 높았던 것으로 보인다. 접목처리구 중 ‘20LM’의 수량이 가장 많은 것으로 보아 ‘20LM’은 뿌리 활력이 다른 대목에 비해 높고 양수분 흡수율과 수분 이용 효율이 증가하여 수확 후기까지 세력을 유지한 것으로 판단된다.
Table 2.
Average fruit weight, the total number of fruits, fruit length, and fruit diameter of non-grafted and grafted cherry tomatoes at 158 days after planting in soil cultivation
| Graft combination |
Fruit weight (g) |
The number of fruits |
Fruit length (mm) |
Fruit diameter (mm) | |
| Scion | Rootstock | ||||
| Nonari | Powerguard | 11.3 cz | 89.8 a | 39.5 c | 25.5 b |
| Nonari | IT173773 | 11.7 c | 85.6 a | 38.9 c | 25.5 b |
| Nonari | 20LM | 14.5 a | 92.6 a | 44.7 a | 28.3 a |
| Nonari | B-blocking | 12.7 b | 90.2 a | 41.8 b | 28.0 a |
| Nonari | - y | 11.6 c | 85.3 a | 36.8 d | 24.9 b |
Table 3.
Changes in the growth strength (mm) of non-grafted and grafted cherry tomatoes grown in soil cultivation
| Graft combination | Growth strengthz (mm) | |||||
| Scion | Rootstock | 66y | 73 | 80 | 87 | |
| Nonari | Powerguard | 9.6 ax | 10.4 a | 8.4 bc | 9.7 a | |
| Nonari | IT173773 | 10.1 a | 10.5 a | 8.9 ab | 9.7 a | |
| Nonari | 20LM | 10.1 a | 10.6 a | 9.9 a | 10.3 a | |
| Nonari | B-blocking | 10.0 a | 10.2 a | 9.1 ab | 9.7 a | |
| Nonari | - w | 9.3 a | 8.6 b | 7.3 c | 7.9 b | |
zGrowth strength was expressed as the average diameter of the stem. The stem diameter was measured at the place where the growing tip was at 7 days before measurements were taken.
정식 후 66일의 생장강도는 처리 간 유의한 차이를 보이지 않다가 정식 후 73일에 비접목처리구가 8.6mm로 감소하였다(Table 3). 정식 후 80일에 ‘20LM’의 생장강도는 9.9mm로 높았으며 비접목처리구는 7.3mm로 낮았다. 정식 후 80일의 경우 모든 처리구가 7–9mm의 상대적으로 낮은 생장강도를 보였는데, 이 시기의 온실 내부 일평균 온도가 9–14°C의 저온에 2주간 노출되어 있었기 때문으로 판단된다. 조사기간 중 온실 내부 일평균 온도는 정식 후 64일에 10.3°C까지 낮아졌으며 정식 후 79일에는 일평균 온도가 9.7°C까지 낮아졌다(Fig. 1A). 토마토는 아열대성 원예작물로 12°C 이하가 되면 생육이 멈추며(Criddle et al., 1997; Ploeg and Heuvelink, 2005) 온도뿐만 아니라 광량은 생육에 영향을 미친다(Stanghellini et al., 2019). 10°C 이하의 일평균 온도를 기록한 기간의 광량 또한 낮아(Fig. 1B) 생육이 부진해진 것으로 판단된다. 생장강도를 측정하면 작물의 힘이 강한지 약한지의 세력을 판단할 수 있는데(Stradiot and Battistel, 2003), 정식 후 80일에 비접목처리구의 생장강도가 낮아진 이유는 저온 스트레스를 받아 세력이 약해졌기 때문으로 판단된다. 지온이 낮으면 뿌리 생육은 부진해지고 양수분 흡수율이 감소하여 작물의 세력 저하로 이어지기 때문에 저온기에 근권부의 활력 정도는 중요하다(Bristow et al., 2021). ‘20LM’은 저온에 내성을 가진 대목으로 저온 스트레스 피해를 극복하여 비접목처리구보다 생장강도가 높았던 것으로 보인다. 대목별로 저온 내성에 대한 근권부 성능에는 차이가 있으며 본 실험에서는 ‘20LM’의 뿌리 활력이 가장 높았고 이에 따라 총 수량이 가장 많았던 것으로 판단된다.
정식 후 66일, 73일 및 80일에 ‘Powerguard’의 개화위치는 22.2cm, 20.8cm 및 24.3cm로 처리구 중 가장 낮은 값을 보였다(Table 4). 정식 후 80일에 ‘20LM’의 개화위치는 다른 처리구에 비해 8–36% 더 높았다. 정식 후 87일에 비접목처리구의 개화위치는 22.3cm로 가장 낮았다. 개화위치를 측정하면 영양생장과 생식생장의 균형을 판단할 수 있는데(Stradiot and Battistel, 2003), 접목처리구 중 ‘Powerguard’는 영양생장이 약해 수확기 중반에 일찍 생식생장상으로 전환되어 생육 후기에 수량이 감소한 것으로 판단된다. 정식 후 59일과 66일의 초장은 처리 간 유의한 차이를 보이지 않다가 ‘Powerguard’의 초장이 정식 후 73일부터 감소하기 시작하여 정식 후 87일에 223.1cm로 가장 짧았다(Table 5). 정식 후 87일에 ‘Powerguard’의 엽폭은 29.2cm로 가장 작았다(Table 6). 영양기관의 생장이 둔화되면 작물 세력은 약화되고 수량이 감소한다(Latifah et al., 2021). ‘Powerguard’는 저온 스트레스를 받아 생장이 약해져 총 수량이 적었으나, ‘20LM’은 같은 환경조건에서 저온에 내성을 가져 수확 후기까지 세력을 유지해 총 수량이 많았던 것으로 판단된다. 이처럼 토마토 생육과 수량 결과는 서로 다른 접목 조합에 따라 다르게 나타난다(Soe et al., 2018). 동일한 처리구 ‘Powerguard’를 수경재배 시설에서 실험한 결과, 생육 후기의 생장강도와 개화위치 값이 낮았고 본 실험 결과와 흡사하게 주당 평균 총 수량이 적었다(Lee et al., 2021). 접수로 완숙토마토 품종인 ‘Gama’를 사용한 실험 결과, ‘Powerguard’의 생육과 수량이 가장 우수하였다(Lee et al., 2020). 대목과 접수 간의 친화성이 높은 접목 조합은 대목에서 접수로의 양수분 흐름이 원활하고 생육이 증진되며 광합성 효율이 높다(Sharma et al., 2019). 반대로 접목 친화성이 좋지 않은 경우 대목의 효과를 보기 어려워 대목 ‘Powerguard’와 방울토마토 접수 품종 ‘Nonari’는 접목 친화성이 낮은 것으로 판단된다. 동일한 대목임에도 완숙토마토 또는 방울토마토 접수에 따라 그 성능이 다르게 나타나는 것을 확인하였고 여러 접목 조합을 검정하는 실험이 필요한 것으로 사료된다.
Table 4.
Changes in the flowering position (cm) of non-grafted and grafted cherry tomatoes grown in soil cultivation
| Graft combination | Flowering positionz (cm) | ||||||
| Scion | Rootstock | 59y | 66 | 73 | 80 | 87 | |
| Nonari | Powerguard | 27.2 ax | 22.2 b | 20.8 c | 24.3 c | 25.1 ab | |
| Nonari | IT173773 | 26.6 a | 40.3 a | 33.3 a | 31.6 ab | 32.3 a | |
| Nonari | 20LM | 22.8 a | 30.3 ab | 27.8 ab | 34.6 a | 31.9 a | |
| Nonari | B-blocking | 26.0 a | 33.4 ab | 26.8 bc | 32.3 ab | 30.3 ab | |
| Nonari | - w | 28.7 a | 29.1 ab | 23.4 bc | 26.7 bc | 22.3 b | |
Table 5.
Changes in the plant height (cm) of non-grafted and grafted cherry tomatoes grown in soil cultivation
| Graft combination | Plant height (cm) | ||||||
| Scion | Rootstock | 59z | 66 | 73 | 80 | 87 | |
| Nonari | Powerguard | 146.7 ay | 165.4 a | 186.8 b | 204.0 b | 223.1 b | |
| Nonari | IT173773 | 147.2 a | 167.2 a | 191.7 ab | 208.8 ab | 229.8 ab | |
| Nonari | 20LM | 148.3 a | 167.8 a | 190.8 ab | 208.8 ab | 229.3 ab | |
| Nonari | B-blocking | 145.3 a | 164.9 a | 191.7 ab | 209.7 ab | 232.1 a | |
| Nonari | - x | 150.0 a | 169.4 a | 193.7 a | 210.9 a | 231.7 a | |
Table 6.
Changes in the leaf length and width (cm) of non-grafted and grafted cherry tomatoes grown in soil cultivation
| Graft combination | Leaf length and width (cm) | |||||||||||||||
| 59z | 66 | 73 | 80 | 87 | ||||||||||||
| Scion | Rootstock |
Leaf length |
Leaf width |
Leaf length |
Leaf width |
Leaf length |
Leaf width |
Leaf length |
Leaf width |
Leaf length |
Leaf width | |||||
| Nonari | Powerguard | 49.5 by | 42.8 ab | 43.5 b | 32.3 a | 45.7 b | 34.9 b | 36.8 b | 23.4 b | 41.3 b | 29.2 d | |||||
| Nonari | IT173773 | 53.5 ab | 49.0 a | 46.2 ab | 35.9 a | 51.3 a | 50.1 a | 49.5 a | 43.7 a | 50.4 a | 46.8 ab | |||||
| Nonari | 20LM | 55.3 a | 44.5 ab | 46.7 ab | 35.8 a | 52.2 a | 49.5 a | 51.5 a | 47.7 a | 51.8 a | 48.6 a | |||||
| Nonari | B-blocking | 53.2 ab | 44.8 ab | 48.8 a | 38.8 a | 49.3 ab | 38.3 ab | 50.5 a | 41.5 a | 49.9 a | 39.9 c | |||||
| Nonari | - x | 51.3 ab | 39.3 b | 44.6 b | 34.1 a | 51.5 a | 43.5 ab | 48.8 a | 39.2 a | 50.2 a | 41.3 bc | |||||
정식 후 45일과 130일의 OJIP 구간별 엽록소형광의 밀도는 처리 간 유의한 차이가 없었다(Fig. 3A and 3C). 정식 후 80일에 OJIP의 P구간에서 ‘B-blocking’의 값은 34539로 높았고 비접목처리구의 값은 28381로 낮았다(Fig. 3B). P구간의 값은 OJIP 지수 FM과 동일한 값으로, 스트레스를 받은 식물은 광계Ⅱ 활성이 저하되어 FM이 감소한다(Bussotti et al., 2011). OJIP는 스트레스 반응에 의해 곡선이 변화하는데(Baghbani et al., 2019) 정식 후 80일에 처리 간 곡선 형태가 다른 이유는 저온 때문인 것으로 판단된다. 저온 스트레스 피해의 주원인은 세포막의 손상으로, 이 손상이 엽록체로 이어져 광합성이 저해된다(Yadav, 2010). 비접목처리구는 저온 스트레스를 받아 세포가 손상되어 광계Ⅱ의 활성이 저하되고 FM 값이 감소한 반면, ‘B-blocking’은 저온에 내성을 가져 FM 값을 유지한 것으로 판단된다. 정식 후 80일에 비접목처리구의 생장강도가 낮은 이유는 저온에 의해 광합성 효율이 감소되고 생장이 지연 및 둔화된 것으로 판단된다.
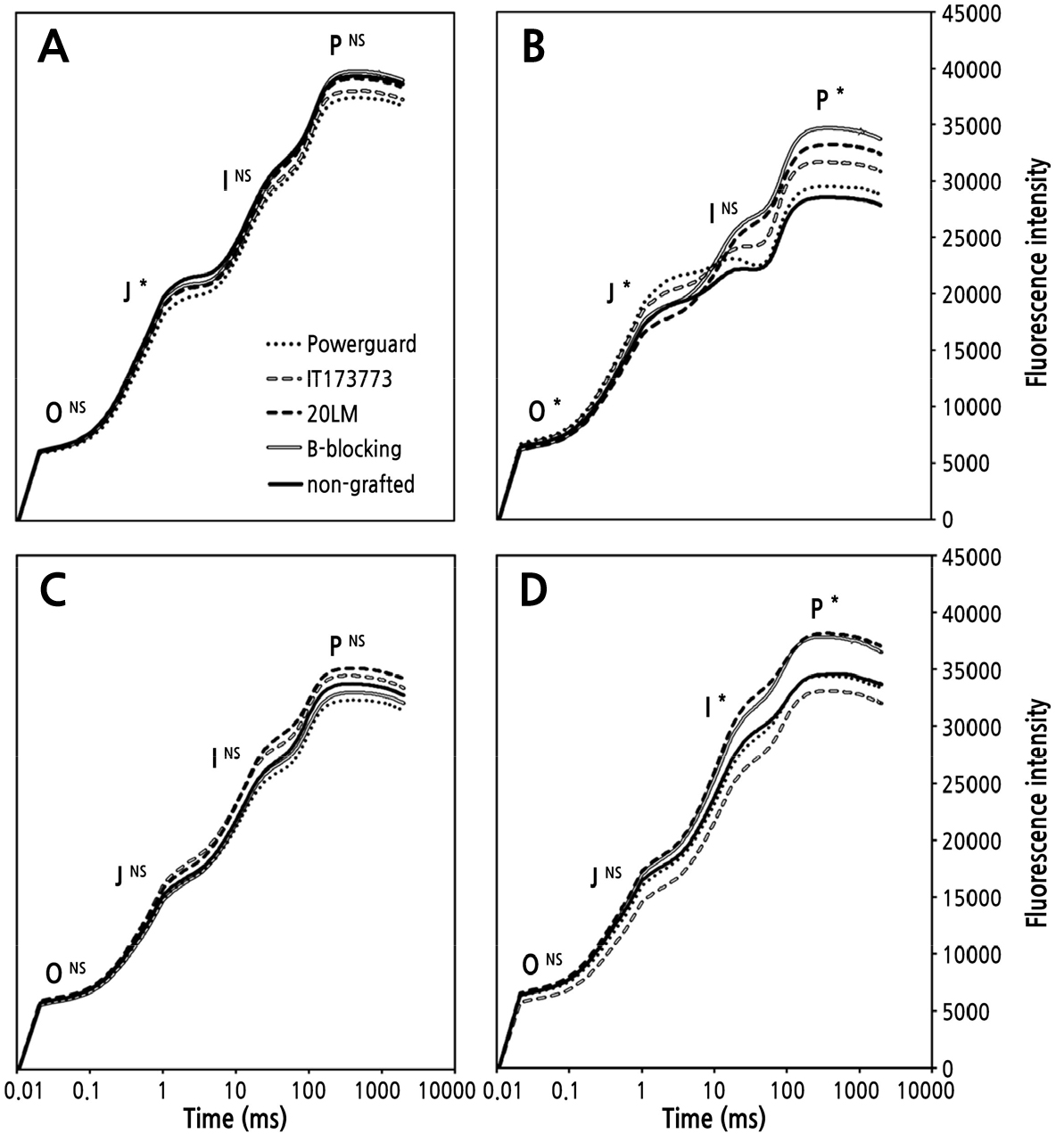
Fig. 3.
Changes in the chlorophyll fluorescence OJIP transients (OJIP curves), plotted on a logarithmic time scale, of non-grafted and grafted cherry tomatoes grown in soil cultivation measured at 45 days after planting (A), 80 days after planting (B), 130 days after planting (C), and 165 days after planting (D).
정식 후 165일에 ‘20LM’과 ‘B-blocking’의 P구간은 37979와 37680으로 나머지 세 처리구보다 높았다(Fig. 3D). 정식 후 165일에 ‘20LM’과 ‘B-blocking’의 주당 평균 누적 수량은 1,339g과 1,142g으로 많았다(Fig. 2). ‘20LM’과 ‘B-blocking’ 처리구는 생육 후기까지 광합성 효율이 높고 이에 따라 수량이 많은 것으로 판단된다. 이로써 생육 후기의 OJIP 차이는 처리 간 작물 세력유지 능력에 따른 광합성 효율의 차이인 것으로 보인다.
정식 후 45일과 130일의 각 지수는 OJIP의 결과와 마찬가지로 처리 간의 차이가 거의 없었다. 정식 후 80일에 ‘Powerguard’의 F0는 6957로 높았다(Table 7). ‘Powerguard’는 저온 스트레스를 받아 에너지 포획능력이 저하되어 F0가 증가하였고, 이는 광계Ⅱ 반응중심의 활성 저하로 이어져 광합성 효율이 감소하여 생장이 줄어들어 개화위치와 엽폭 등의 값이 낮았던 것으로 판단된다. 정식 후 80일에 ‘Powerguard’와 비접목처리구의 FV/FM은 0.762와 0.767로 낮았다. 일반적으로 FV/FM 값이 0.8보다 낮을 때는 광계Ⅱ의 반응중심이 손상을 입거나 불활성 상태라고 할 수 있다(Bolhàr-Nordenkampf et al., 1989; Oh et al., 2012). 정식 후 80일에 ‘20LM’과 ‘B-blocking’의 PIABS는 3.260과 3.226으로 나머지 세 처리구에 비해 유의적으로 높았고 ‘IT173773’은 1.838로 낮았다. PIABS는 다른 지수에 비해 스트레스에 민감하여(Živčák et al., 2008) 처리 간 값의 차이가 큰 것으로 판단된다. 정식 후 80일에 ‘20LM’과 ‘B-blocking’의 ET0/RC는 0.863과 0.824로 높았고 ‘Powerguard’는 0.544로 낮았다. ‘20LM’과 ‘B-blocking’은 저온조건에서도 광계Ⅱ의 전자전달효율이 높은 것으로 판단된다.
Table 7.
Minimal fluorescence yield (F0), maximal fluorescence yield (FM), maximum quantum yield (FV/FM), performance index (PIABS), and electron transport flux (ET0/RC) measured at 45, 80, 130, and 165 days after planting in non-grafted and grafted cherry tomatoes in soil cultivation
| Days after Planting | Graft combination | F0 | FM | FV/FM | PIABS | ET0/RC |
| 45 | Powerguard | 6177 bz | 37309 a | 0.834 a | 3.984 a | 0.813 a |
| IT173773 | 6419 ab | 37954 a | 0.831 a | 3.551 a | 0.826 a | |
| 20LM | 6307 ab | 39024 a | 0.838 a | 4.012 a | 0.831 a | |
| B-blocking | 6340 ab | 39663 a | 0.839 a | 4.081 a | 0.832 a | |
| non-grafted | 6484 a | 39236 a | 0.834 a | 3.590 a | 0.804 a | |
| 80 | Powerguard | 6957 a | 29390 ab | 0.762 c | 1.078 b | 0.544 c |
| IT173773 | 6794 ab | 31514 ab | 0.782 bc | 1.838 b | 0.701 b | |
| 20LM | 6355 b | 33047 ab | 0.804 ab | 3.260 a | 0.863 a | |
| B-blocking | 6391 b | 34539 a | 0.813 a | 3.226 a | 0.824 a | |
| non-grafted | 6621 ab | 28381 b | 0.767 c | 1.498 b | 0.657 b | |
| 130 | Powerguard | 5849 a | 32168 a | 0.818 a | 4.349 a | 0.824 ab |
| IT173773 | 6099 a | 34294 a | 0.822 a | 4.224 a | 0.779 b | |
| 20LM | 6123 a | 34928 a | 0.825 a | 4.615 a | 0.835 ab | |
| B-blocking | 5788 a | 32848 a | 0.823 a | 4.701 a | 0.830 ab | |
| non-grafted | 5985 a | 33578 a | 0.820 a | 4.511 a | 0.858 a | |
| 165 | Powerguard | 6526 ab | 34242 c | 0.809 bc | 3.764 ab | 0.903 b |
| IT173773 | 5965 b | 32935 c | 0.819 abc | 4.657 a | 0.908 ab | |
| 20LM | 6827 a | 37979 a | 0.820 ab | 4.267 ab | 0.949 a | |
| B-blocking | 6645 ab | 37680 ab | 0.824 a | 4.527 a | 0.924 ab | |
| non-grafted | 6624 ab | 34453 bc | 0.807 c | 3.371 b | 0.941 ab |
정식 후 130일에 ‘IT173773’의 ET0/RC는 0.779로 처리구 중 가장 낮았고(Table 7), 이때 ‘IT173773’의 주당 평균 누적 수량은 268g으로 가장 적었다(Fig. 2). 정식 후 165일에 비접목처리구의 FV/FM은 0.807로 가장 낮았다. 정식 후 130일과 165일에 측정된 모든 처리구의 FV/FM은 0.8 이상으로 광계Ⅱ의 활성상태는 양호하나, 정식 후 165일에는 처리 간 유의한 차이를 보였다. 정식 후 165일에 비접목처리구의 PIABS는 3.371로 낮았고, ‘Powerguard’의 ET0/RC는 0.903으로 낮은 반면 ‘20LM’은 0.949로 높았다. 광합성 효율이 떨어지면 FV/FM과 PIABS, ET0/RC의 값이 감소하는데(Sousaraei et al., 2021) 비접목처리구의 경우 생육 후기의 FV/FM과 PIABS가 낮아 결과적으로 수량이 적은 것으로 판단된다. 접목 토마토는 실생 토마토에 비해 광합성 효율이 높아 수량이 많은 것으로 알려져 있으나 접목 조합에 따라 그 성능이 다르게 나타나기 때문에(Soe et al., 2018) ‘Powerguard’와 ‘IT173773’의 경우 광계Ⅱ 전자전달효율이 낮아 광합성 효율이 떨어져 수량이 적은 반면, ‘20LM’은 전자전달효율이 높아 생육 후기까지 작물 세력을 유지하여 수량이 많은 것으로 판단된다. 처리 간 OJIP 지수를 이용한 광합성 효율 측정을 통해 대목 종류에 따른 작물 세력의 평가가 가능한 것으로 판단된다.
