

서 언
우리나라의 감 산업은 최근 20여 년간 재배면적 확대에 따라 그 규모가 급격히 증가하여 과수산업의 중요한 위치를 차지하고 있다. 그러나 재배기술의 기본이 되는 수체 생리에 대한 지식은 해외 문헌에 주로 의존해 왔다. 이에 우리나라 기후조건에 맞지 않는 정보가 많아 재배자들에게 과학적인 과원 관리 방법을 제시하는데 어려움을 겪고 있다. 특히 양분 관리의 기초가 되는 수체의 양분 조성 변화에 대한 자료는 매우 부족한 실정이다.
감나무는 결과모지로 남긴 1년생 가지의 선단 부위에서 봄에 눈이 트고 신초가 자라면서 줄기, 잎, 과실(화뢰)이 함께 생장한다. 신초는 5월 하순경 개화 무렵까지 길이 생장이 이루어지지만 잎과 과실은 개화 후에도 생장을 계속한다(Nii, 1980; Yoon et al., 2012). 다른 과수에서 생장 초기 어린 잎은 동화능력이 작기 때문에 어느 정도 자란 후라야 광합성산물을 생장에 이용하며 다른 기관으로 분배할 수 있다(Hale and Weaver, 1962; Oliveira and Priestley, 1988; Quinland and Weaver, 1969). 따라서 봄 동안의 신초생장은 저장양분의 영향을 크게 받아, 발아와 신초생장 직전에 영구기관의 탄수화물과 질소화합물 이동이 시작되고, 그것이 선단조직에 집중되어 생장에 사용된다(Davis and Sparks, 1974; Loescher et al., 1990; Oliveira and Priestley, 1988; Titus and Kang, 1982). 감은 개화기 무렵에 뿌리 활동이 본격적으로 시작되어 토양으로부터의 양분 흡수가 늦으며, 발아부터 개화기 사이에 신초와 화뢰 생장이 동시에 이루어지므로, 저장양분이 생장 초기의 영양생장 뿐만 아니라 과실생장에 직접적인 영향을 줄 수 있다(Choi et al., 2005; Hirata et al., 1974; Kim et al., 2009; Park and Kim, 2011).
원활한 신초생장을 위해서는 생장 초기에 유기화합물과 더불어 무기양분의 분배도 중요한데, Stassen et al.(1983)은 10년생 복숭아 나무에서 발아부터 이후 8주까지 새로운 생장에 사용한 질소(N)의 80%가 영구조직의 저장기관으로부터 유래되었음을 보고하였다. 같은 기간 동안 인(P)은 43%, 칼륨(K)은 40% 이하가 영구조직으로부터 유래한 반면 마그네슘(Mg)은 새로운 흡수를 통하여 얻어진 것에 의존하는 것 같다고 하였다. 그러나 감의 경우 잎의 연간 양분조성 변화는 여러 연구자들에 의해 보고되어 왔지만(Clark and Smith, 1990; George et al., 1995; Park, 2002; Park et al., 2003), 신초생장 초기 줄기, 잎, 과실 등 부위별 양분 변화를 자세히 추적한 연구결과는 찾기 어렵다.
이 시기의 신초 양분 조사는 나무의 생리를 이해하고 과학적인 양분 관리를 위한 기초자료 도출을 위해 필요하다. 본 연구에서는 남부지방 단감의 주품종인 ‘부유’를 대상으로 신초생장이 왕성한 전엽기부터 신장이 정지한 후까지 주기적으로 줄기, 잎, 과실 부위별 양분의 조성과 분배 양상을 파악하고자 하였다.
재료 및 방법
1998년과 1999년에 시료로 채취한 감나무는 경남 사천시 정동면 예수리에 소재하는 사양토의 경사지 과원에 6 × 5m 거리로 재식되어 있었으며, 시험을 시작할 때 18년생이었다. 2005년에는 경남 김해시 진영읍의 평지과원에 6 × 3m 거리로 재식된 10년생 감나무를 사용하였다. 조사 과원은 과다 착과가 되지 않도록 5월 상・중순에 관행적으로 적뢰를 하였는데, 신초 길이가 20cm 미만일 때는 1개, 그 이상으로 신초의 세력이 강할 때는 2개의 화뢰를 남기는 방법으로 하였다. 그 결과 정단신초당 남긴 평균 화뢰수는 1998년에 1.8개, 1999년에 1.5개, 2005년에 1.7개였다. 시험 과원은 관행적인 시비와 병해충 방제로 관리 상태가 양호하여, 양분의 과다 또는 부족 증상이 나타나지 않았고 병해충 피해도 미미하였다. 한 나무에 완전히 전개된 잎이 5% 정도 보일 때를 기준으로 한 전엽기는 1998년에 4월 11일, 1999년에 4월 19일, 2005년에는 4월 17일이었으며, 만개기는 각각 5월 21일, 5월 25일, 5월 22일이었다. 3년간 신초의 길이 생장이 끝난 시기는 각각 5월 25일, 5월 31일, 5월 30일이었으며, 최종 길이는 각각 26.9, 33.0, 36.2cm였다.
본 연구의 영양분석과 해석은 선행 연구(Yoon et al., 2012)에서 생장 조사에 사용되었던 ‘부유’ 감나무(Diospyros kaki cv. Fuyu)의 신초 시료를 대상으로 하였다. 과원을 3구역으로 나누어 구역별로 서로 다른 나무의 수관 외부에서 길이 23 ± 8cm인 결과모지의 정단신초를 10개씩 총 30개를 각 시기마다 채취하였다. 시료 채취는 1998년에는 4월 20일부터 6월 1일까지 2주 간격, 1999년에는 4월 21일부터 5월 31일까지 2주 간격으로 하였으며, 2005년에는 4월 25일부터 5월 31일까지 3-4일 간격으로 채취하였다. 전엽 직후 채취한 신초는 줄기와 과실(화뢰)의 크기가 너무 작아 분리할 수 없었으므로 잎 시료에만 포함시켰으며, 줄기와 과실로 분리가 가능한 시기부터는 세 부위로 나누어 시료로 사용하였다.
분석용 시료는 80°C에서 48시간 건조시켜, 20mesh 체를 통과하도록 분쇄한 후 가용성당, 전분, 아미노산, 단백질 등의 유기화합물과 무기원소를 분석하였다. 가용성당은 건조시료 100mg에 80% ethanol(v/v) 0.9mL를 첨가하여 80°C 항온수조에서 30분간 용출시킨 후 15,000 × g에서 5분 동안 원심분리하여 상징액을 모으는 과정을 2회 반복하여 추출하였다. 남은 잔사에 4.5N HClO4 0.9mL를 첨가하여 실온에서 30분간 용출시킨 후 15,000 × g에서 5분 동안 원심분리하여 얻은 상징액을 전분으로 간주하였으며, 이 추출도 2회 반복하였다. 추출 후 anthrone 반응(McCready et al., 1950)으로 가용성당과 전분을 정량하였으며, 포도당(glucose)을 표준당으로 사용하였다. 아미노산은 건조시료 100mg에 80% ethanol(v/v) 0.9mL를 첨가하여 80°C에서 30분간 용출시킨 다음 15,000 × g에서 5분간 원심분리하여 상징액을 모으는 과정을 2회 반복하여 추출하였고, 표준 아미노산으로 L-leucine을 사용하여 ninhydrin법(Yemm and Cocking, 1955)으로 정량하였다. 단백질은 아미노산을 추출한 후 남은 잔사에 0.1N NaOH0.9mL를 넣고 실온에서 30분간 용출시킨 후 15,000 × g에서 5분간 원심분리하여 상징액을 모으는 과정을 2회 반복하여 추출하고, 표준 단백질로 bovine albumin을 사용하여 Bradford 방법(Bradford, 1976)으로 정량하였다. 무기원소는 농촌진흥청 식물체 분석법(NIAST, 2000)에 따라 건조시료 500mg에 H2SO4와 HClO4 분해액을 첨가하여 전열판에서 분해 후 총 N은 Kjeldahl법으로, P는 vanadate 반응으로 분석하였고, K, Ca, Mg는 원자흡광장치(AA-6501F, Shimadzu Co., Tokyo)로 측정하였다.
분석한 성적으로 일정량의 시료 속에 포함되어 있는 양분의 농도와, 농도와 건물중을 곱한 부위별 함량을 구하였고, 아래의 상대생장률(RGR, relative growth rate) 공식을 응용하여 신초 신장 초기와 종료기에 신초당 양분 함량의 상대적인 변화율을 구하였다.
양분 함량의 상대적인 변화율 = (ln W2 - ln W1)/(t2 - t1)
ln : 자연대수, t1, t2 : 조사시점
W1 : t1 때의 양분 함량, W2 : t2 때의 양분 함량
결과 및 고찰
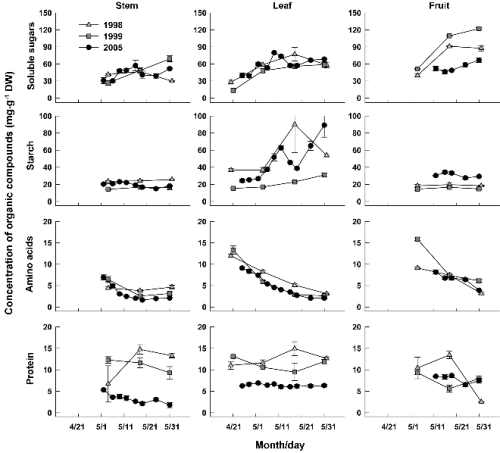
신초생장기 동안 신초의 부위별 유기화합물의 농도 변화는 Fig. 1과 같았다. 가용성당 농도는 잎과 과실에서 높아지는 경향이었으나 줄기에서는 일관된 변화가 없었다. 전분 농도는 줄기와 과실에서 큰 변화가 없었고 잎에서만 연차와 시기별로 폭 넓은 차이를 보이며 증가하는 양상이었다. 그 결과 신초의 세 부위 중에서 잎의 전분 농도가 가장 높아, 2005년 5월 하순의 경우 31-89mg・g-1로 줄기의 3배 이상이었다. 이시기 호주 지역의 ‘부유’에서 줄기의 전분 농도는 신초가 신장할 동안 증가한다고 한 반면(George et al., 1994), 일본에서는 본 연구결과처럼 뚜렷한 변화가 없다고 보고되었다(Fukuda and Kuroi, 1949). 이러한 결과의 차이에는 기후적인 요인이 어느 정도 작용했을 것으로 생각된다. 아미노산 농도는 신초생장과 함께 세 부위에서 모두 감소하였는데, 2005년의 경우 줄기에서 5월 2일 6.9mg・g-1에서 5월 31일 2.1mg・g-1로, 같은 시기 잎에서는 7.4mg・g-1에서 2.1mg・g-1로, 과실에서는 5월 12일 8.2mg・g-1에서 5월 31일 3.9mg・g-1로 낮아졌다. 초기 생장 동안에 줄기와 잎의 단백질 농도는 1998년과 1999년보다 2005년에 현저히 낮은 농도를 나타냈으나 세 부위에서 경시적인 증감 변화는 명확하지 않았다.
이들 탄수화물과 질소화합물의 신초당 함량은 잎에서 가장 높아 5월 동안에 각 양분 총 함량의 60% 이상을 차지하였다(Fig. 2). 잎의 가용성당과 전분 함량은 5월 중순부터 증가가 정체되었던 1998년을 제외하면 초기 생장기 동안 큰 폭으로 증가하였고 2005년에 가장 많았다. 이 시기에 감나무 어린잎의 광합성 능력이 낮음에도 불구하고(George et al., 1997) 잎의 탄수화물 함량이 급격히 증가한 것은 저장양분의 역할이 컸기 때문일 것이다(Loescher et al., 1990; Park and Kim, 2011). 1998년 5월 하순에 양분의 증가가 이루어 지지 않은 것은 신초 시료채취 때에 발생한 실험오차 때문으로 여겨진다. 가용성당 함량은 줄기와 과실에서도 증가하는 경향이었으나 증가 폭은 크지 않았다. 반면 줄기와 과실의 전분 함량은 농도 변화에서도 알 수 있듯이 증가가 미미하였다. 탄수화물과 마찬가지로 아미노산 및 단백질 함량은 잎에서 급속히 증가하여 5월 하순에는 줄기와 과실에 비해 각각 2배 이상 많았다. 그러나 아미노산과 단백질 함량이 1998년 5월 하순에 과실에서 감소한 경우를 제외하면, 신초생장기 동안 신초와 과실에서 꾸준히 증가하는 경향인 것으로 파악되었다. 사과나무에서 질소화합물은 나무의 휴면이 끝나면 저장기관에서 단백질의 가수분해와 함께 새로운 생장기관으로 이동하는 현상(O’Kennedy and Titus, 1979; Titus and Kang, 1982)과 마찬가지로, 본 연구에서도 저장된 질소화합물이 신초로 공급된 것으로 판단된다.
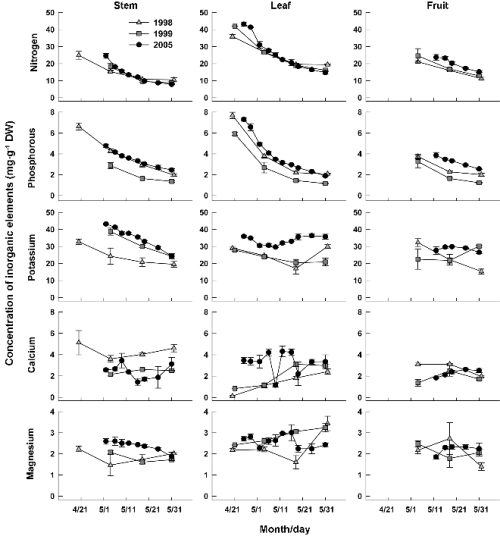
Fig. 3은 줄기, 잎, 과실의 시기별 무기원소 농도 변화를 나타낸 것이다. 세 부위에서 N 농도는 생장 초기에 가장 높았다가 생장이 진행됨에 따라 낮아졌는데, 줄기와 잎의 경우 5월 중순까지 큰 폭으로 감소하였고 이후 5월 하순까지는 완만하게 감소하는 양상이었다. P 농도는 신초의 모든 부위에서 감소하였으며 그 정도는 잎에서 가장 뚜렷하였다. 2005년의 경우 P는 5월 2일에 줄기에서 4.8mg・g-1로부터 5월 31일에는 2.5mg・g-1로 낮아진 반면, 잎에서는 이 기간에 4.9mg・g-1에서 1.9mg・g-1로 감소하였다. K 농도는 생장이 진행됨에 따라 줄기에서는 감소하였고, 잎과 과실에서는 일관된 변화가 없이 해에 따라 다른 양상을 보였다. 잎의 K 농도는 다른 해보다 2005년에 특히 높게 유지되기도 하였다. Ca와 Mg 농도는 부위별로 경시적인 변화가 뚜렷하지 않았고 해에 따른 변이도 컸다. 생장 초기 감나무 줄기와 과실의 무기원소 농도 변화는 기존성적이 없어 비교할 수 없으나, 잎의 N과 P 농도는 전엽기에 가장 높았다가 이후에 급격히 감소한다는 선행 연구결과(Choi et al., 2011; Clark and Smith, 1990; George et al., 1994)와 유사하였다. 잎의 K 농도도 변화가 크지 않아 기존 연구결과(Clark and Smith, 1990; George et al., 1994)와 크게 다르지 않았다. 그러나 잎의 Ca 및 Mg 농도는 전엽 후 점진적으로 증가한다고 하였으나(Clark and Smith, 1990; George et al., 1994) 본 연구의 2005년에는 이러한 경향이 나타나지 않았는데, 이런 차이가 재배환경에 따른 영향인지는 추후 검토해야 할 것으로 생각되었다.
|
Fig. 1. Changes in concentrations of organic compounds in stem, leaves, and fruits (including flower buds) of ‘Fuyu’ shoot from the time of foliation to growth cessation. Bars indicate SE. |
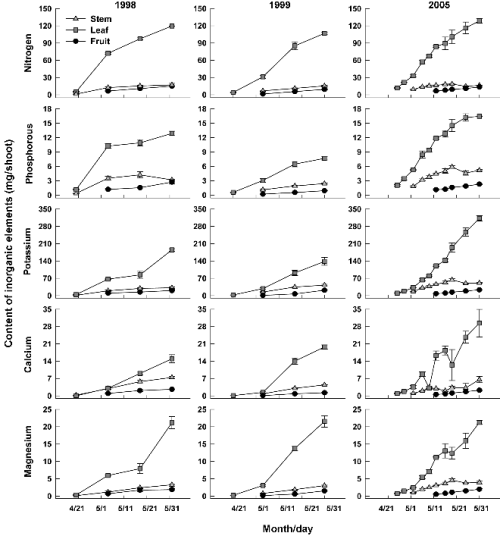
경시적인 농도의 증가가 없었음에도 불구하고 신초생장에 따른 건물중 증가로 인하여(Yoon et al., 2012), 무기원소의 신초당 부위별 함량은 모두 증가하였다(Fig. 4). 특히 잎의 무기원소 함량이 가장 많고 증가도 뚜렷하였는데 5월 중순 이후에는 잎이 각 무기원소 총 함량의 54-82%를 포함하였다. 이는 초기 생장기 동안 잎이 신초 건물중의 60% 이상을 차지하기 때문(Yoon et al., 2012)으로 볼 수 있다. 감나무의 뿌리 활동과 양분흡수가 5월 이후부터 본격적으로 이루어지는 점(Mowat and George, 1994; Nakamura, 1935)을 감안하면, 이와 같은 초기 신초생장기 동안의 무기원소는 상당 부분이 영구기관에 저장되었던 것으로부터 유래된 것으로 추정된다. Tagliavini et al.(1997)은 배나무에서 발아 후 55일에 단과지와 신초의 잎에 포함된 N의 50%가 저장되어 있던 양분에서 유래한 것이라 하였는데, 감의 경우 5월 24일 신초의 N 함량이 신초생장기에 저장기관에서 감소된 N 함량과 비슷한 것으로 보고되었다(Kim et al., 2009).
|
Fig. 2. Changes in contents of organic compounds in stem, leaves, and fruits (including flower buds) of a ‘Fuyu’ shoot from the time of foliation to growth cessation. Bars indicate SE. |
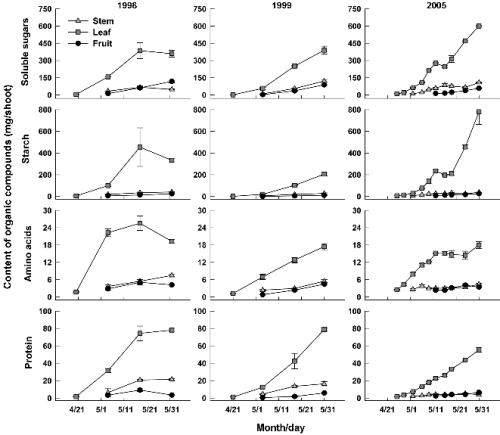
Table 1은 정단신초의 길이 생장이 완료된 때의 단위 신초당 유기화합물과 무기원소 함량을 나타낸 것으로 신초생장을 위해 축적된 양분의 양을 의미한다. 3년간 가용성당과 전분 함량은 각각 526-768과 245-844mg의 범위에 있었으며 2005년에 가장 높았다. 탄수화물에 비해 아미노산과 단백질 함량은 현저히 적었는데 각각 26-31과 66-103mg 범위였다. 무기원소 중에서는 K 함량이 203-388mg으로 가장 많았고 N이 다음 순으로 132-159mg이었다. 다른 해보다 2005년에 아미노산과 단백질을 제외한 양분의 함량이 대체적으로 많은 것은 그 해에 신초생장이 왕성했기 때문(Yoon et al., 2012)로 생각된다.
신초로 분배되는 양분의 경시적 증가 양상을 파악하기 위하여 초기 생장기간 동안에 양분 함량의 상대적인 변화율을 구하여, 신뢰할 수 있는 조사가 가능한 가장 이른 시기의 값과 외형적인 신초생장이 정지된 시기의 마지막 값을 서로 비교하였다(Table 2). 3년간 조사한 유기화합물과 무기원소의 상대변화율은 모두 초기 값이 마지막 값보다 월등히 높았다. 이러한 결과는 정단신초의 전체 생장기간을 놓고 볼 때 전엽 직후의 전반부가 생장 정지시기에 가까운 후반부보다 양분 증가속도가 월등하게 높음을 의미한다고 볼 수 있다. 전엽 직후의 신초 양분 증가는 당년의 동화산물이나 뿌리 흡수와는 사실상 무관한 결과라 할 수 있으므로, 초기의 높은 증가속도는 나무의 저장양분에 의해 결정된 것이라 할 수 있다. 그러나 상대변화율이 조사년도에 따라 차이가 나는 것을 볼 때, 저장양분뿐만 아니라 당년의 기상이나 재배 조건에 의한 영향도 큰 것으로 생각된다.
|
Fig. 3. Changes in concentrations of inorganic elements in stem, leaves, and fruits (including flower buds) of ‘Fuyu’ shoots from the time of foliation to growth cessation. Bars indicate SE. |
|
Fig. 4. Changes in contents of inorganic elements in stem, leaves, and fruits (including flower buds) of a ‘Fuyu’ shoot from the time of foliation to growth cessation. Bars indicate SE. |
본 연구결과를 통해 신초생장기 동안 신초 각 부위의 양분 조성 변화를 조사하여 영양 생리적 특성을 파악하였고, 신초의 각 부위로 분배되는 시기별 양분량을 구함으로써 감나무의 단위 신초생장에 필요한 양분 함량을 산출할 수 있었다. 저장양분의 분배가 신초의 초기 생장기에 특히 활발했던 점으로 미루어 이 시기에 저장양분의 역할이 매우 중요함을 알 수 있었다. 따라서 원활한 신초생장을 위해서는 이에 관여하는 유기・무기양분의 축적이 전년부터 적절히 조절되어야 할 것으로 판단된다.
