서 언
재료 및 방법
유년성 지속기간 구명시험
자생 마삭줄의 유년상과 성년상 잎의 형태 조사
군집분석과 주성분 분석
결과 및 고찰
유년성 지속 기간 구명
국내 자생 마삭줄의 유년기와 성년기 잎의 형태 조사
군집분석과 주성분 분석
서 언
마삭줄(Trachelospermum L.)은 상록 덩굴 식물로서 전 세계에 30여종(species)이 존재하며 국내에는 주로 경남, 전남, 전북, 충남 서해안, 경기도 덕적도까지 분포하는데(Lee, 2002; Park et al., 2011). 마삭줄(Trachelospermum asiaticum var. intermedium)과 털마삭줄(Trachelospermum jasminoides var. pubescence)이 주를 이룬다(Kang and Shim, 1997; Ryoo, 2005).
마삭줄은 한국, 중국, 일본에서는 옛날부터 약용식물로 해열, 진통, 거풍(擧風), 종기 지혈의 효능이 있고 산후 어혈(産後 瘀血), 혈압 강하, 관절통, 통풍, 신경통, 요통, 타박상, 편도선염, 부스럼 등의 치료에 이용하는데 재배보다는 주로 자연에서 채취해서 이용하고 있다. 반면에 서양에서는 주로 지피식물로 이용되며 국내에서도 최근에 일부 조경재료로 사용하기도 하나 내한성이 약하여 남부 해안지역에서만 주로 이루어진다(Ingaki et al., 1972; Abe and Yamaguchi, 1986; Hotta et al., 1989; Moon, 2015).
마삭줄은 잎의 변이가 심한 식물로 지금까지 분포(Park et al., 2011), 번식(Moon, 2015), phylogenetic relationship(Moon et al., 2019b) 등에 대한 연구는 있었으나 국내외에서 유년기(juvenile stage)에서 성년상(adult phase)으로 전환하는 기간에 대한 연구는 없었다. Moon et al.(2019a)은 남해안 7곳에 자생하는 마삭줄의 엽형(leaf shape)을 조사하여 유년형(juvenile form) 잎이 성년형(adult form)에 비해 상대적으로 엽형지수(leaf shape index)가 큰 것과 성년상 줄기에 나타나는 접촉굴성(thigmotropism)을 확인하였다. 아울러 유년기를 지나 성년상이 되면 유백색 엽맥 무늬가 옅어지거나, 사라지는 현상도 보고하였다. 또한, 이와 같이 마삭줄의 잎의 변이와 줄기의 굴성이 변하는 것은 환경적 요인 외에 이 식물의 상적변환(phase change)과 밀접한 관련이 있다고 하였다(Moon et al., 2019a). 목본 식물중 상적변환시 극적인 형태의 변화가 나타나 외형이 크게 달라지는 전형적인 식물로 잉글리쉬아이비(Hedera helix), 유칼립투스(Eucalyptus), 슈도파낙스(Pseudopanax) 등 몇 가지 식물들이 알려져 있다. 유칼립투스의 유년형 잎은 둥근 형태를 띠지만 성년형 잎은 버드나무 잎처럼 피침형이 되고 슈도파낙스유년형 잎은 긴 가시모양을 하며 아래를 향하지만 성년상이 되면 잎이 짧고 넓어지며 위를 향한다(Hartmann et al., 2011). 마삭줄처럼 유년기에 포복 생장을 하는 아이비(Hedera helix)의 경우 유년기가 약 5–10년간 지속되며 성년상이 되면 잎의 결각이 사라지고 둥근 형태의 성년형 잎이 나타나며 주로 포복생장을 하던 줄기는 직립하게 된다(Hartmann et al., 2011). 마삭줄의 경우에도 잎과 줄기에 변화가 나타나는데, 마삭줄의 유년성(juvenility) 줄기는 주로 포복생장을 하지만 부착근을 내어 수목이나 바위에 부착하여 오르는 성질을 함께 가지고 있다. 성년상이 나타나면 아이비의 경우처럼 포복생장을 멈추고 직립하는 성질과 함께 감는 굴성이 추가적으로 나타난다. 이때 새로 출현하는 성년형 잎은 유백색의 선명한 엽맥이 사라지며 유년기에 비해 더 크고 둥근 형태를 띤 잎이 출현한다(Moon et al., 2019a). 식물의 잎의 형태는 온도나 착생부위 등에 따라 변화된다(Chitwood and Sinha, 2016). 또한 과수, 채소, 화훼류의 생육상의 변화는 외적으로 일장이나 온도, 내적으로 내생 호르몬이나 C/N율에 따라 유기된다(Park, 2019). 생식생장기로 전환이 되면 외적인 잎의 형태가 배추는 로제트형으로 변화하고, 고수(coriander) 등은 세엽화가 되며, 서양겨자무는 톱니처럼 잎이 변화하는 현상 등이 나타나 상변화를 알 수 있다(Park, 2018). 이처럼 재배하는 초본 작물의 경우 대체로 재배기간이 목본식물에 비해 짧고 그 기간 내에 변화가 나타나므로 해당 작물의 생육상 변화를 쉽게 감지하고 대처할 수 있지만 목본 식물에서의 유년성 지속기간은 장미(Rosa spp.)의 경우처럼 짧게는 20–30일, 유럽 너도밤나무(Fagus sylvatica)처럼 길게는 30–40년이 경과해야만 유년성이 타파되어 개화 등 변화가 나타나므로(Hartmann et al., 2011) 이러한 목본 식물의 경우 원하는 식물의 생산이나 번식, 기타 원예적 이용에 있어서는 그 식물에 대한 유년성 지속기간이나 상적변환에 대해 충분한 이해가 필요하다. 그러나 자생 마삭줄의 상변화와 이와 관련된 유년기에 대해서는 알려진 바가 거의 없다. 따라서, 마삭줄의 외형적인 형태변화를 관찰하여 상적변환이 이루어졌는지를 가늠할 수 있는 지표가 설정된다면 분화 생산이나 식용꽃 생산 등 재배적 이용뿐만 아니라 마삭줄의 유년상(juvenile phase)과 성년상의 서로 다른 형태와 특성을 이용한 번식이나 계획적인 육종에도 적용될 수 있어 유년성 지속기간과 형태 구명은 매우 중요하다(Matsoukas, 2014). 그러나, 국내외에서 마삭줄의 분포, 자생지역, 식물이나 종간의 유연관계 등의 연구(Park et al., 2011)는 이루어졌으나 지금까지 마삭줄의 유년성 지속기간의 구명이나 성년상에 나타나는 잎과 줄기의 형태(엽형, 엽면적, 엽두께, 엽맥무늬 여부, 줄기의 굴성 여부)변화에 대한 연구는 없거나 부족한 상태이다 따라서, 본 연구는 마삭줄의 재배시험과 자생지 조사를 통해 개화 전 후의 잎과 줄기에 공통으로 나타나는 특성을 구명하여 이를 상변화의 지표로 설정하고 파종 후 상변화가 나타나는 시기까지 경과한 기간을 환산하여 국내 자생 마삭줄의 유년성 지속기간을 구명하는데 그 목적이 있다.
중국 야생종 모란(Han et al., 2021)과 30종의 이란 돌나물(Taghipour et al., 2021), 그리고 46종의 감귤(Kaur et al., 2022) 등은 군집분석을 수지도(dendrogram)로 나타내어 유전적 유연관계를 보고한 바 있는데, 국내 자생 마삭줄에 있어서는 군집분석을 통한 유전적인 다양성에 관하여 연구된 바 없었다. 또한, 주성분 분석(principal component analysis, PCA)은 방대한 자료를 간단하게 표기하여 다변량 자료 분석에 도움을 주는 통계 방법으로써, 야생 국화의 유전적 다양성(Nguyen et al., 2020)과 북부 키프로스와 터키에서 수집한 30종의 멜론의 형태적 특성(Yılmaz et al., 2021)을 주성분 기여도 도출법으로 분석 보고된 바 있어, 이를 적용한 마삭줄 지역종 분석도 필요하다.
이에 본 연구는 국내 자생 마삭줄(Trachelospermumjasminoides)의 잎과 줄기의 형태적 변화를 통해 상변화 여부를 판단할 수 있는 지표를 설정하고, 상변화시까지 유년성지속기간을 구명하고 군집분석과 주성분 분석을 통해 지역별로 군집화와 형태적 특성의 주요 기여도를 알아보기 위하여 수행하였다.
재료 및 방법
유년성 지속기간 구명시험
본 연구는 경기도 남양주시 조안면 삼봉리 ‘풀나무 농장’에서 수행되었다. 자생 마삭줄의 유년성 지속기간을 구명하기 위하여 2015년 4월 26일 풀나무 농장에서 채취한 보성산 털마삭줄(Trachelospermum jasminoides) 종자를 직경 18cm 플라스틱 포트에 독일산 배양토(Blumenerde, GRAMOFLOR Co., Vechta, Germany)를 사용하여 25립 파종한 후 발아 된 11개체를 육묘하였다. 육묘한지 3년이 되는 2018년 5월 12일 줄기 길이가 70–100cm 사이로 자란 6개체를 1-6까지 개체별 고유 번호를 부여하고 나무 펜스를 지지대로 하여 약 25cm 간격으로 정식하였다. 홀수(1, 3, 5)는 줄기가 위를 향하도록 하여 케이블타이를 사용하여 부착하였고 짝수(2, 4, 6)는 부착하지 않고 아래를 향하도록 하여 폭 7m, 측고 3m, 동고 5m, 길이 25m의 3연동형 비닐 하우스 온실 포장에 정식하여 관리하였다. 피복재료로 0.1mm 폴리에틸렌(polyethylene: PE) 필름을 사용하였고. 겨울철 시설 내 야간 온도가 –7°C 이하로 내려가지 않도록 자동 온도조절 온풍기를 설치하여 관리하였다. 봄부터 가을까지는 지붕, 측창 등을 개방하여 외부 환경에 가깝도록 실내온도, 습도 등을 유지하였다. 온실내 포장의 서쪽 1m지점의 3m 높이에 55% 차광막을 설치하여 오전에 볕이 들고 오후에는 그늘이 지게 하였다. 3m이상 상향으로 자라 차광막보다 높이자란 줄기는 별도의 차광을 하지 않았다. 관수는 토양의 건습을 고려 관행적인 방법으로 수분관리를 하였다. 재배온실의 토양은 EC 0.32dS/m, pH 5.5로 조사되었고 별도의 시비는 하지 않았다. 재식 후 매년 생장의 변화를 관찰하던 중 파종 후 6년째인 2021년 5월에 6개체 중 2개체에서 개화하였다. 개화 등 변화가 나타난 위치를 확인하고자 6개체에 1-6까지 부여된 고유번호로 각 개체의 원줄기를 표시했고, 원줄기에서 나온 1차 분지(아들줄기, secondary vine)의 경우 1번 개체를 예로 들면 1-1, 그리고 1차 분지에서 나온 1차 분지(손자줄기, tertiary vine)는 1-1-1과 같은 방법으로 표시하였다.
조사 항목은 줄기 길이(Stem length: SL, cm, 지제부로부터 정단부까지 길이), 줄기 방향(Direction of stem: DOS, 펜스에 부착하여 위로 자라는 줄기는 Up, 펜스에 부착하지 않고 포복하는 줄기는 Down), 첫 굴성 위치(Location of first thigmotropism: LOT, cm, 지제부로부터 첫 굴성이 나타난 지점까지 거리), 개화 위치(Location of flowering: LOF, cm, 개화 위치까지 지제부로 부터 거리), 첫 성년형 잎의 출현위치(Location of first adult leaf: LAL, cm, 지제부로부터 첫 성년형 잎까지 거리), 마지막 유년형잎의 위치(Location of last juvenile leaf: LJL, cm, 지제부로부터 마지막 유년형잎까지 거리) 등을 조사하였다.
또한, 개화가 이루어진 1번과 6번개체를 대상으로 유년기 잎과 성년기 잎의 특징을 조사하였다. 성년형 잎의 조사방법은 2개체에서 개화한 총 5개의 꽃차례에서 꽃자루가 포함되고 하부에 생장이 완료된 성년형 잎이 4매이상 부착된 4개 가지를 무작위로 선발하고 각 가지의 화경으로부터 2번째 마디 이하에 부착된 생장이 완료된 잎을 위에서 아래 순서대로 4개씩 채취하였고, 유년형 잎의 조사는 위와 동일한 2개체의 아래쪽 포복줄기에 부착된 가지에서 조사 가능한 4개 가지를 무작위로 선발하여 생장점으로부터 2번째 마디 이하에 부착된 생장이 완료된 잎을 위에서 아래 순서대로 4개씩 채취하였다. 유년형과 성년형 각 각 4개의 가지에서 각 4개씩 총 32개의 잎을 조사하여 가지당 4반복으로 총 16반복 하였다. 각각 엽폭(Leaf width: LW, cm), 엽장(Leaf length: LL, cm), 엽면적(Leaf area: LA, cm2), 엽두께(Leaf thickness: LT, mm)를 엽면적 측정기(LI-3100, LI-COR, NE, USA)와 디지털 캘리퍼스(Digimatic Caliper; CD-15CPX, Mitutoyo, Kawasaki, Japan)를 사용하여 측정하고 유년형과 성년형의 엽형지수(Leaf shape index: LI, LI = LW/LL)를 산출하여 비교하였다. 유 ․ 성년형을 구분하기 위해 각 요소의 약어 앞에 유년형은 J-, 성년형은 A-로 표기하였다. Microsoft Excel program(version 2019, Microsoft Corp., CA, USA)을 사용하여 표준오차를 표시하였고, SPSS statistics program(version 26, IBM Corp., NY, USA)을 사용하여 독립표본 T검정을 진행하였다.
자생 마삭줄의 유년상과 성년상 잎의 형태 조사
자생 마삭줄의 유년상과 성년상 잎 특성을 알아보기 위하여 2018년 5월부터 2021년 5월까지 3년에 걸쳐서 강원도 고성에서부터 동해안을 따라 부산까지 그리고 대구 이남지역의 남해안, 서해안은 진도에서부터 강화도까지 전국의 마삭줄 분포지를 조사하였다. 동해안 쪽은 최북단지역으로 경주, 내륙은 청도(경북), 전주(전북), 남쪽은 제주(구좌), 서귀포, 오동도(여수), 진도, 부산 그리고 서해안은 강화도(인천), 백아도(인천) 굴업도(인천), 익산, 신시도(군산) 등 13곳에서 샘플을 채취하였다(Table 1). 조사방법은 각 지역별로 상부에 개화가 이루어지고 하부에 개화하지 않은 포복줄기가 존재하는 개체를 먼저 발견한 순서대로 무작위로 선발하여 재배 시험과 동일한 방법으로 조사 가능한 유년형과 성년형 가지 각 1개씩을 무작위로 선발하고 잎을 4개씩 채취하여 총 8개의 잎을 같은 방법으로 조사하였다. 전국 13개지역에서 지역별 1개체씩 총 104개의 잎을 조사하여 개체당 4반복으로 하였다. 각각 엽폭, 엽장, 엽면적. 엽두께를 엽면적 측정기(LI-3100, LI-COR, NE, USA)와 디지털 캘리퍼스(Digimatic Caliper; CD-15CPX, Mitutoyo, Kawasaki, Japan)를 사용하여 측정하고 유년형과 성년형의 엽형지수(LI = LW/LL)를 산출하여 비교하였다. 유 ․ 성년형을 구분하기위해 각 요소의 약어 앞에 유년형은 J-, 성년형은 A-로 표기하였다. Microsoft Excel program(version 2019, Microsoft Corp., CA, USA)을 사용하여 표준오차를 표시하였고, SPSS statistics program(version 26, IBM Corp., NY, USA)을 사용하여 독립표본 T검정을 진행하였다.
Table 1.
Collection areas of native Trachelospermum in Korea
군집분석과 주성분 분석
SPSS statistics(version 24, IBM Corp., NY, USA)을 사용하여 마삭줄 채취 지역별 유년기와 성년기 잎의 형태 요인(엽폭, 엽장, 엽형지수, 엽면적, 그리고 엽두께)에 따른 잎의 형태 유사도를 알아보기 위해 위의 조사방법과 같이 무작위로 선발된 유년형, 성년형 가지 각 1개씩의 잎 4개를 채취하여 조사한 데이터를 군집분석하여 수지도(dendrogram)로 나타내었고, 형태적 특성의 주요 기여도를 알아보고자 주성분 분석(principal component analysis, PCA)(Choi et al., 2021)을 실시하였다.
결과 및 고찰
유년성 지속 기간 구명
전남 보성산 털마삭줄(Trachelospermum jasminoides) 종자를 파종 후 5년째인 2020년 가을(8–9월) 줄기의 상부에 감는 굴성이 나타나고, 6년째인 2021년 5월 개화반응이 나타났다. 공시한 6개체의 29개 줄기의 길이는 280–830cm였다(Table 2). 원줄기의 길이는 1번 개체가 830cm로 가장 길었고 2번 개체가 410cm로 가장 짧았다. 3개체(1, 5, 6)의 5개 줄기는 감는 굴성이 나타났고, 그 중 2개체(1, 6)에서 개화하였다. 첫번째 굴성이 나타난 부위까지 지제부로부터 최고 거리는 1번 개체 720cm, 최저 거리는 6번 개체 420cm였다. 정단부로 부터 아랫 쪽으로 첫 굴성이 나타난 지점까지 거리는 최대 약 190cm(1), 최단 80cm(5)로 나타났다. 개화가 이루어진 1번과 6번 개체를 대상으로 줄기의 방향과 길이를 조사한 결과 1번 개체의 경우 9개의 줄기 중 상 방향 5개, 하 방향 4개 줄기였다. 그 중 개화 한 3개 줄기(1, 1-5, 1-5-2)의 길이는 670–830cm로 1번개체에서 가장 긴 줄기 상위 3개에 해당하며 모두 상 방향이었다. 그리고 상 방향 중 길이가 340cm로 가장 짧은 줄기(1-2)에도 성년형 잎이 나타났다. 그러나 하 방향의 4개줄기중 3개줄기(1-1,1-4, 1-2-1)는 370–490cm로 상 방향의 가장 짧은 줄기 보다 길었지만 하 방향의 모든 줄기는 잎과 줄기에 외적 형태변화가 나타나지 않았다. 또한 개화가 이루어진 6번 개체에서는 상 방향 4개 줄기, 하방향 3개 줄기로 개화가 이루어진 2개 줄기는 모두 상 방향이었으며 원줄기(6)가 610cm로 가장 길었으나 원줄기를 포함한 하 방향 3개 줄기는 잎과 줄기의 형태적 변화 및 개화 반응이 나타나지 않았다. 반면 상향 4개 줄기 중 가장 긴 줄기(6-2)는 530cm로 개화는 이루어지지 않았지만 감는 굴성과 성년형 잎이 출현하였고, 가장 짧은 줄기(6-2-1)에서 굴성은 나타나지 않고 성년형 잎의 출현과 함께 개화하였다. 또한 상방향인 나머지 개화줄기(6-4-1)도 350cm로 성년형 잎이 나타나며 개화가 이루어졌다. 다만 상방향 중 1개의 줄기(6-4)에서는 개화 등 상변화가 나타나지 않았다. 또한 조사된 총 6개체의 29개 줄기는 상방향 14개, 하방향 15개로 이중 성년상이 나타나거나 개화한 줄기는 11개였고 모두 상 방향이었다. 이러한 결과는 상향줄기 14개중 11개에 해당하는 79%의 줄기에서 상변화가 나타난 반면 하 방향의 경우는 개화가 이루어진 개체에서 가장 긴 줄기(6)는 물론이고 조사된 총 15개 하향 줄기는 외형의 변화나 개화가 전혀 이루어지지 않은 것으로 이는 자라난 줄기의 방향이 상 방향인 경우 줄기의 길이가 긴 줄기에서 개화 등 성년상이 먼저 나타나는 경향이 있지만 줄기의 길이 보다는 방향이 상적변환을 촉진하는 요인으로 작용한 결과이다. 본 연구에서 나무펜스를 지지대로 하여 홀수개체(1, 3, 5)가 상 방향이 되도록 한 시험 방법은 2번개체를 제외한 5개 개체에서 의도와는 달리 방향이 제각각 새로운 줄기를 형성하며 자랐으나 결과적으로 줄기의 방향이 상변화를 촉진하는 요소로 작용하였을 것으로 판단되었다. 다만 같은 개체의 줄기라도 첫 개화시 상향으로 자란 줄기에서 개화가 이루어지고 아래로 향한 줄기에는 상변화가 나타나지 않은 것은 마삭줄의 번식과 밀접한 관련이 있을 것으로 추측되었다. 즉, 포복성을 띠는 유년성 줄기는 주로 지표면을 따라 영양번식을 하고, 부착근을 형성하며 물체에 기어오른 유년성 줄기는 부착물을 넘어 다시 내려가거나, 계속 올라가 종자번식을 하게 되는데 이는 유년성 줄기에 동시에 존재하는 2가지 성질(포복성과 부착하여 오르는 성질)이 복합적으로 작용하여 환경에 따라 번식장소와 방법을 선택하는 것으로 여겨진다. 즉, 자연상태에서 작은 바위나 수목 등 극복할 수 있는 낮은 장애물일 경우 타고 넘어가는 기작으로 나타나며 큰 바위나 수목을 만나 계속하여 높이 오른 줄기는 광 환경이 바뀌고 바람의 영향을 받게 되므로 새로운 환경에 적합한 종자번식을 선택하는 과정에서 줄기의 굴성이 변하여 포복성을 띠던 줄기는 직립성을 나타내며 감는 줄기가 나타나는 것으로 추측된다. 이로 인해 같은 개체에서도 줄기의 방향에 따라 번식방법이 달라지고 종자번식을 선택하여 상향으로 자란 줄기는 먼저 상변화의 특징이 나타나 같은 개체에서도 유년과 성년상이 극명하게 갈리는 것으로 추측되었다. 아이비(Hedera helix), 송악(Hedera rhombea) 등 에서도 같은 줄기에 유년상과 성년상이 극명하게 나타난다(Hartmann et al., 2011; Park, 2019). 본 연구에서 조사된 총 29개 줄기는 상향 13개, 포복 16개로 나타났고 그중 상향 후 포복 1개(3-1), 포복 후 상향 2개(1-2, 3) 등 몇 가지 유형이 나타났지만 개화줄기를 비롯하여 엽형의 변화, 엽맥무늬의 변화, 접촉굴성의 변화 등이 나타난 줄기의 최종 방향은 모두 상향이었다(Table 2). Kato et al.(2012)은 밝은 광환경에 심어 자라는 T. asiaticum의 기는 싹은 음의 굴광성을 나타내어 어두운 곳으로 빠르게 자라 지지 숙주에 도달할 수 있다고 했다. 즉, 기는 줄기는 광도가 낮은 상태에서 빠르게 생장하여 다시 원줄기 방향으로 자라 상향하는 특성을 가진다고 했다. 본 연구에서 공시재료로 사용된 자생 마삭줄(T. jasminoides)의 하향 줄기가 땅에 발근했다가 다시 상향하는 특성이 나타난 것은 국내에 자생하는 마삭줄 중에서 털마삭줄(T. jasmioides) 줄기도 자라면서 재차 지지식물에 부착하여 상향으로 자라서 굴성을 나타내고 개화를 하려는 특성이 있음을 보여준다. 즉, 하향줄기는 포복경 마디에서 딸기의 런너처럼 부정근을 형성하여(Park, 2019), 지표면에 뿌리를 내리고 양분을 흡수하여 자라다가 어느 시기가 되면 다시 상향으로 자라는 특성이 있음을 의미한다. 전국 마삭줄 분포지 조사와 관찰을 하는 과정에서 바위에 기어오르거나 수목에 감는 줄기를 갖는 개체, 또는 평지에서도 서로 얽히며 상향으로 감는 줄기를 갖는 개체에서 주로 개화가 이루어지는 것을 관찰하였다. 이는 지면에서 나무줄기를 타고 곧장 올라가 충분한 햇빛을 받고 상변화 단계가 되면 마삭줄은 오른 방향(반시계방향)으로 감으면서 생장하는데(Moon et al., 2019a), 이는 개화 후에 녹두꼬투리처럼 생긴 무거운 꼬투리(15–20cm)를 달리게 하고 익으면 꼬투리가 자동으로 열리면서 바람에 의하여 털이 달린 종자를 멀리 비산되게 하기 위한 환경적인 적응으로 사료된다. 본 연구 결과에서 첫 개화시 상변화가 일어난 모든 줄기의 방향이 상 방향으로 나타난 것은 줄기의 방향이 상변화를 촉진하는 결과이며 이는 마삭줄의 개화를 위해 유년기간을 단축하기 위한 물리적 수단으로 줄기의 방향을 인위적으로 선택할 수 있음을 의미하며 이는 마삭줄의 재배나 이용시 유용한 수단이 될수 있을 것으로 판단되었다.
Table 2.
Changes in leaf and stem characteristics during the juvenile and adult phases of native Trachelospermum jasminoides. (6th year of cultivation, May 2021)
| No. of plantsz | SLy (cm) | DOS |
LOT (cm) |
LOF (cm) |
LJL (cm) |
LAL (cm) |
| 1 | 830 | Up | 640 | 520 | 520 | 520 |
| 1-1 | 370 | Down | - | - | - | - |
| 1-2 | 340 | Up | - | - | 210 | 220 |
| 1-2-1 | 490 | Down | - | - | - | - |
| 1-3 | 320 | Down | - | - | - | - |
| 1-4 | 370 | Down | - | - | - | - |
| 1-5 | 820 | Up | 720 | 570 | 340 | 460 |
| 1-5-1 | 520 | Up | - | - | - | - |
| 1-5-2 | 670 | Up | 610 | 560 | 340 | 460 |
| 2 | 410 | Down | - | - | - | - |
| 2-1 | 310 | Down | - | - | - | - |
| 2-2 | 305 | Down | - | - | - | - |
| 3 | 530 | Up | - | - | 225 | 255 |
| 3-1 | 500 | Down | - | - | - | - |
| 3-2 | 280 | Down | - | - | - | - |
| 4 | 570 | Up | - | - | 265 | 345 |
| 4-1 | 490 | Up | - | - | 130 | 270 |
| 4-2 | 240 | Down | - | - | - | - |
| 5 | 660 | Up | 580 | - | 400 | 440 |
| 5-1 | 500 | Down | - | - | - | - |
| 5-2 | 440 | Down | - | - | - | - |
| 5-3 | 390 | Up | - | - | - | - |
| 6 | 610 | Down | - | - | - | - |
| 6-1 | 300 | Down | - | - | - | - |
| 6-2 | 530 | Up | 420 | - | 320 | 320 |
| 6-2-1 | 280 | Up | - | 280 | 280 | 280 |
| 6-3 | 450 | Down | - | - | - | - |
| 6-4 | 360 | Up | - | - | - | - |
| 6-4-1 | 350 | Up | - | 310 | 280 | 310 |
마삭줄의 파종 후 감는 줄기는 2020년 가을에 처음으로 나타났다. 감는 굴성은 주로 정단부 쪽의 회전운동으로 나타나므로 굴성이 처음 나타날 당시(2020년 8월–9월) 그 지점에는 해당 줄기의 정단부가 위치했을 것으로 추측되었다. 따라서, 굴성이 나타난 지점으로부터 최단 80cm에서 최장 190cm의 새로 자란 줄기들은 이미 유년기가 완료된 화숙(Ripeness to flower)상태에 접어든 줄기로 판단되었다. 화숙 상태의 식물은 그 식물이 처한 환경에 따라 개화 반응이 나타나거나 그렇지 않을 수도 있다고 하였다(Davies et al., 2018). 첫 굴성이 나타난 지점의 아래쪽 부위는 당시(2020년 8월–9월)모두 유년성 잎이 부착된 줄기였지만 이듬해(2021년 5월) 새로 성장을 시작하는 원줄기(1, 3, 4, 5), 아들 줄기(1-2, 1-5, 4-1, 6-2), 또는 손자 줄기(1-5-2, 6-2-1, 6-4-1)에서는 성엽의 출현이나 개화 등이 모두 굴성이 나타난 지점의 아랫쪽 부위에서 이루어졌다(Table 2). 이는 굴성이 나타난 5개 줄기 중 3개 줄기(1, 1-5, 1-5-2)에서 개화하고 2개 줄기(6-2-1, 6-4-1)는 개화하지 않았으나, 첫 굴성이 나타난 모든 줄기에서 성년형 잎이 출현한 결과로 이는 개화여부에 상관없이 상적변환이 이루어진 것을 의미한다 반면, 굴성을 나타내지 않고 개화한 2개 줄기를 포함해 개화한 5개의 모든 가지에서 성년형 잎이 먼저 나타나고 개화가 이루어 진 것은 개화 전에 반드시 성년형 잎이 출현하는 특성의 결과이다. 따라서 줄기에 첫 감는 굴성이 나타나는 것과 엽맥이 사라지며 유년에 비해 크고 둥근 형태의 성년형 잎이 나타나는 현상은(Moon et al., 2019a) 본 연구의 재배시험에 사용된 자생 마삭줄 중 털마삭줄(T. jasminoides)의 상적변환의 특성이 될 수 있을 것으로 판단되었다(Fig. 1). Fig. 1에서 나타난 엽맥 무늬와 형태가 달라지고(A) 줄기의 굴성(B)이 형성되는 특성은 환경적 요인 외에 마삭줄의 유년기와 성년기에 따른 변화임을 구명한 선행 결과와 일치하였다(Moon et al., 2019a).

개화는 감는 굴성이 나타난 아랫 줄기 부분에서 나타나는 것으로 미루어 굴성이 나타난 줄기는 화숙 상태에 접어든 것을 의미한다고 생각된다. 따라서 개화반응은 유년성 줄기에 존재하는 개화억제물질의 감소나 성년상 변화에 따라 나타난 새로운 개화 호르몬 물질이 상부 잎에서 합성되어서 아들 순이나, 손자 순으로 전이되는 것으로 추측된다(Ream et al., 2012). 이때 Fig. 1에서처럼 마삭줄의 잎끝이 뾰족하고 엽맥이 선명한 유년성 잎이 성년성 잎이 되면 잎 끝이 둥글고 엽맥이 옅어지는 상적변환이 이루어진다. 같은 덩굴성 식물인 아이비(Hedera helix), 송악(Hedera rhombea) 등 에서는 결각이 사라지며 둥근 형태의 잎이 나타나(Hartmann et al., 2011; Park, 2019), 성년화 잎의 특성은 식물에 따라 차이를 보인다.
최근 Poethig(2013)은 이와 같은 식물의 상변화는 유전인자가 제어한다고 주장하면서 목본식물인 Acacia confuse, Acacia colei, Eucalyptus globulus, Hedera helix, Quaercus acutissima 등 6개 식물에서 유년기에는 miR156과 miR157이 발현되고 엽형이 완전히 변하는 성년기에는 miR172가 나타난다고 했다. 마삭줄에서 굴성이 나타난 하부에 성년상 잎이 나타나고 개화가 되는 것은 유년기가 종료되었음을 의미하며 그 경계는 굴성이 나타난 아랫 줄기의 마지막 유년형 잎과 첫 성년형 잎의 출현 구간으로 추측된다. 초본식물에서도 상변화에 따라 잎의 형태적인 변화가 특별한 상부 줄기에서 나타나는데 고수, horseradish, chervil, 애기장대, 수수 등에서 보고되었다(Matsoukas, 2014; Park, 2018; Hashimoto et al., 2019).
마삭줄의 유년상에서 성숙상으로 변화에 miR156, miR157, miR172인자가 관여하는지(Poethig, 2010, 2013) 그 이외 생화학적인 영향인지를 앞으로 연구를 통하여 확인할 필요가 있다고 본다. 이는 유년상에서 성숙상으로 변화하는 식물에 따라 합성하는 생화학물질이 작용하기 때문인데 자작나무에서는 peroxidase와 esterase isozymes(Brand and Lineberger, 1992), 세코이어(Sequoia sempervirens)에서 protein phosphorylation(Huang et al., 1992) 등이다. 또한, 식물 호르몬의 영향이 보고되었는데 옥신의 영향(De Zeeuw and Leopold, 1955), 아이비에서 앱시스산(abscisic acid, ABA)(Rogler and Hacken, 1975)이 관여한다고 했다. 그 이외 florigen, auxin, gibberellin(GA) 등 식물생장호르몬이 관여하는지는(Wadhi and Ram, 1967; Matsoukas, 2014; Park, 2019) 추가적인 연구를 통하여 구명할 필요가 있다고 본다.
본 연구에 공시재료로 사용된 6개체중 3개체가 5년째에 성년상을 나타냈고 그중 2개체가 6년째에 개화한 것은 50%개체에서 5년만에 상변화가 일어났고 33%는 파종 후 6년 만에 첫 개화반응을 보인 결과로 이는 자생마삭줄(T.jasminoides)의 유년성이 약 5년 지속되고 개화를 위해서는 최소한 6년의 재배기간이 필요 하다는 것을 의미 하는데 이는 유년기가 5–10년 동안 지속되는 아이비(Hedera helix), 털자작나무(Betula pubescens), 세코이어(Sequoia sempervirens)와 유사하며, 15–20년간 유년상이 지속되는 구주물푸레나무(Franxinus exseldior)와 플라타너스단풍(Acer pseudoplatanus) 보다 짧다(Park, 2019). 자생 마삭줄의 보다 자세한 화성유기를 추정하기 위해서는 품종별 파종시험과 더불어 유년성의 최대치에 대한 추가적인 연구가 필요할 것으로 생각되었다.
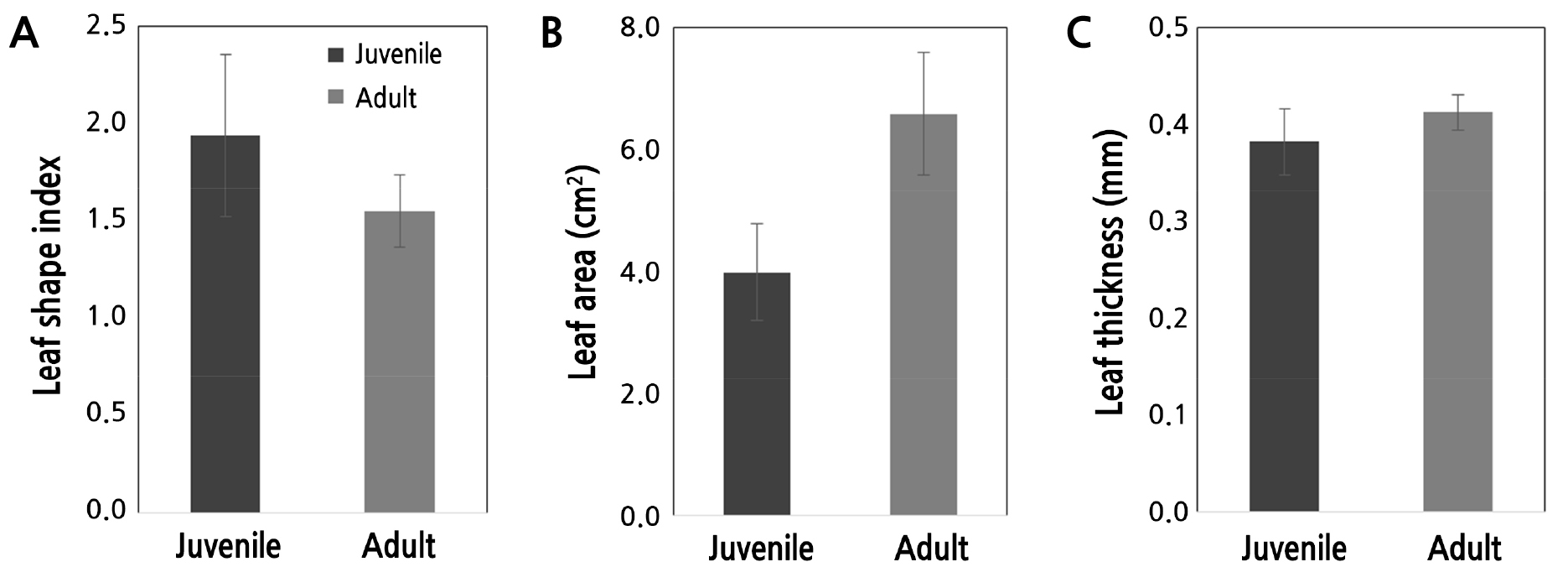
개화가 이루어진 1번과 6번 개체를 대상으로 유년기 잎과 성년기 잎의 특징을 조사한 바 엽형지수는 유년기 잎이 1.92, 성년기 잎이 1.55로 나타났으며, 엽면적은 유년기 때 3.99cm2 성년기 때 6.59cm2로 나타났고, 엽두께는 유년기와 성년기에 각각 0.38, 0.41mm이었다. 즉, 엽형지수는 유년형 잎이 약 1.2배 크고, 엽면적은 성년형 잎이 약 1.7배 크며 엽두께는 성년형 잎이 더 두꺼웠다. 유년기와 성숙상의 엽두께는 유의미한 차이가 나타나지 않았다(Fig. 2). 유년과 성년형 잎의 엽형지수의 차이는 Moon et al.(2019a)의 결과와 일치하였다. Park(2019)은 유년상과 성년상 변화와 관련하여 잎의 형태는 언급했으나 엽형지수나 엽면적에 대하여서는 다루지 않은 바 앞으로 상변화가 있는 다양한 식물을 대상으로 엽형지수나 엽면적을 비교할 필요가 있을 것으로 사료된다.

국내 자생 마삭줄의 유년기와 성년기 잎의 형태 조사
전국에 자생하는 마삭줄 유년기 잎의 형태를 조사한 결과(Table 3 and Fig. 2), 엽폭은 백아도가 2.88cm로 가장 넓고 진도가 1.20cm로 가장 좁았다. 잎 길이는 백아도 5.38cm, 굴업도 5.20cm, 경주 5.05cm이었으며 서귀포가 2.15cm로 가장 짧았다. 대체로 진도를 제외하고는 서해상의 섬에서 자란 마삭줄이 유년기 잎의 면적이 컸다. 엽형지수는 전주가 2.69로 컸고, 익산이 1.54로 가장 작게 나타났다(Fig. 3). 엽면적과 엽두께는 백아도가 가장 크고 두꺼웠으며 진도가 가장 작고 얇았다.
Table 3.
Changes in leaf width, leaf length, and leaf thickness during the juvenile and adult phases of Trachelospermum in Korea
| Area |
Leaf width (cm) |
Leaf length (cm) |
Leaf thickness (mm) | |||||
| Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | |||
| Baegado | 2.88 ± 0.03 a | 3.88 ± 0.11 bc | 5.38 ± 0.13 a | 6.43 ± 0.17 bc | 0.77 ± 0.04 a | 0.81 ± 0.08 ab | ||
| Busan | 1.30 ± 0.04 efg | 2.45 ± 0.10 fg | 3.45 ± 0.17 c | 4.88 ± 0.46 ef | 0.57 ± 0.04 bd | 0.62 ± 0.01 cde | ||
| Cheongdo | 2.13 ± 0.03 bc | 2.58 ± 0.05 efg | 4.55 ± 0.05 ab | 4.85 ± 0.06 ef | 0.48 ± 0.02 cde | 0.51 ± 0.02 defg | ||
| Ganghwado | 1.70 ± 0.13 cde | 2.33 ± 0.03 g | 3.53 ± 0.14 c | 4.30 ± 0.18 f | 0.36 ± 0.01 ef | 0.39 ± 0.02 fgh | ||
| Gureopdo | 2.63 ± 0.21 a | 4.72 ± 0.26 a | 5.20 ± 0.25 a | 7.48 ± 0.21 ab | 0.68 ± 0.05 ab | 0.84 ± 0.05 a | ||
| Iksan | 1.88 ± 0.03 bcd | 2.83 ± 0.11 defg | 2.90 ± 0.24 cd | 4.40 ± 0.22 ef | 0.52 ± 0.01 cd | 0.56 ± 0.03 def | ||
| Jeju | 1.63 ± 0.06 defg | 3.28 ± 0.13 cd | 3.60 ± 0.14 bc | 7.60 ± 0.12 a | 0.40 ± 0.04 def | 0.67 ± 0.05 bcd | ||
| Jeonju | 1.80 ± 0.07 bcd | 3.08 ± 0.08 def | 4.85 ± 0.14 a | 6.13 ± 0.25 cd | 0.44 ± 0.01 cde | 0.48 ± 0.01 efg | ||
| Jindo | 1.20 ± 0.04 g | 3.20 ± 0.09 de | 2.30 ± 0.04 d | 4.78 ± 0.18 ef | 0.21 ± 0.03 g | 0.35 ± 0.00 gh | ||
| Kyeongju | 2.17 ± 0.07 b | 3.88 ± 0.08 bc | 5.05 ± 0.38 a | 6.43 ± 0.14 bc | 0.48 ± 0.01 cde | 0.78 ± 0.03 abc | ||
| Odongdo | 1.68 ± 0.14 cdf | 3.98 ± 0.23 b | 3.30 ± 0.35 c | 7.75 ± 0.18 a | 0.27 ± 0.00 fg | 0.48 ± 0.01 efg | ||
| Seogwipo | 1.25 ± 0.03 fg | 2.55 ± 0.15 efg | 2.15 ± 0.09 d | 5.03 ± 0.27 def | 0.47 ± 0.03 cde | 0.56 ± 0.01 de | ||
| Sinsido | 1.68 ± 0.03 def | 3.25 ± 0.10 cd | 3.53 ± 0.10 c | 5.45 ± 0.17 cde | 0.48 ± 0.01 cde | 0.59 ± 0.04 de | ||
| Average | 1.84 | 3.23***z | 3.83 | 5.81*** | 0.47 | 0.58 | ||
성년기의 엽장, 엽폭, 엽두께는 Table 3과 같다. 오동도와 제주도의 엽장은 각각 7.75cm, 7.60cm로 가장 길었고, 6cm 이상 되는 잎은 백아도, 전주, 경주에서 발견되었다. Park et al.(2011)은 제주와 남해안에서 엽장이 6cm 이상인 것을 발견하지 못했다고 보고했는데 본 연구에서는 이상 5곳에서 발견되었다. 엽폭은 굴업도 마삭줄이 4.72cm로 가장 길었고 강화도가 2.33cm로 가장 짧았다. 성년기의 가장 큰 잎의 엽폭과 엽장은 유년기의 가장 큰 잎과 비교하면 각각 1.64배, 1.94배로 크게 차이를 보였다.
엽형지수를 조사한 결과(Fig. 4), 성년기는 제주가 2.32로 가장 컸고, 진도가 1.49로 가장 작았다. 제주의 경우 성년 엽형지수가 유년기(2.21)보다 커 성년형 잎이 유년보다 엽장이 더 긴 형태로 나타났고 유년형 잎도 평균(2.098)이상으로 길었다. 이는 일반적으로 성년형이 유년형보다 더 둥근 형태로 나타난 것과는 반대이며 이는 종내 유전적인 차이가 있거나 다른 요인이 있을 것으로 생각되었다. 그러나 익산과 서귀포, 제주를 제외한 10곳에서 유년성의 엽형지수가 성년에 비해 크게 나타났고 잎의 형태적 차이로 유년과 성년을 구분하는데 유의차가 나타났다. 또한, 조사지 13곳의 유년기 평균 지수 2.098과 성년기 1.818은 유의성 있는 차이를 보였다. 본 결과는 2019년도 본 연구팀이 남해안 지역 7곳의 유년기의 엽형지수가 성년성 보다 컸던 결과와 일치하였다(Moon et al., 2019a). 그러나 Davies et al.(2018)이 제시한 유칼립투스는 유년기 잎이 둥글고 성년기 잎은 버드나무잎처럼 가늘고 길게 변한다고 하였다. 따라서 엽형지수는 식물에 따라 유성년을 구별하는데 지표로 삼을 수 있지만 수치의 변화는 식물의 종류, 품종 등에 따라 다르게 나타날 것으로 생각되었다. 그러나 지금까지 본 연구의 결과로 국내산 마삭줄은 엽형을 통해 유 ․ 성년을 구분하는데 무리가 없으며 다만 유의차가 인정되지 않은 지역은 종내의 변이 등 다른 요인이 영향을 미쳤을 것으로 판단되었다.

엽면적은 굴업도에서 23.49cm2로 가장 넓었는데 이는 유년기의 백아도 잎보다 약 2.3배가 되는 면적이었다. 진도 성년기 마삭줄의 엽면적은 5.91cm2였는데 유년기 잎에 비하여 약 4.8배의 차이를 보였다. 엽면적은 조사한 13곳 모두에서 유년성 잎에 비해 성년엽이 컸고 동시에 지역 간에도 차이가 나타났는데 이들 두 조사 결과는 모두 유의성 있게 나타났다(Fig. 5). 이와 같은 엽면적의 변화는 송악에서도 나타나는데 성년기 잎이 유년기 잎보다 월등하게 컸다(Park, 2019). 엽면적의 크기는 지역이나 환경뿐만 아니라 품종에 따른 유전적 영향이 있을 것으로 추정되어 지역별 절대적 크기의 비교는 유년과 성년의 구분하는데 무리가 따르지만 같은 환경 조건이라고 추정할 수 있는 1개체 내의 다른 줄기 등에서 유 ․ 성년을 판별하기 위한 상대적 크기의 비교는 본 연구에서 나타난 결과와 같이 지역과 품종을 가리지 않고 전 지역에서 뚜렷한 차이가 나타나 유 ․ 성년을 판별할 때 변별력이 매우 크다고 할 수 있다. 따라서 유 ․ 성년간 잎의 상대적인 크기를 비교한 엽면적의 변화는 자생마삭줄의 상변화를 가늠할 수 있는 중요한 요소가 될 수 있을 것으로 판단되었다. Park et al.(2011)은 전국의 마삭줄 분포지역을 조사 하면서 엽면적의 변화 등 상변화에 대해서는 언급하지 않았다.
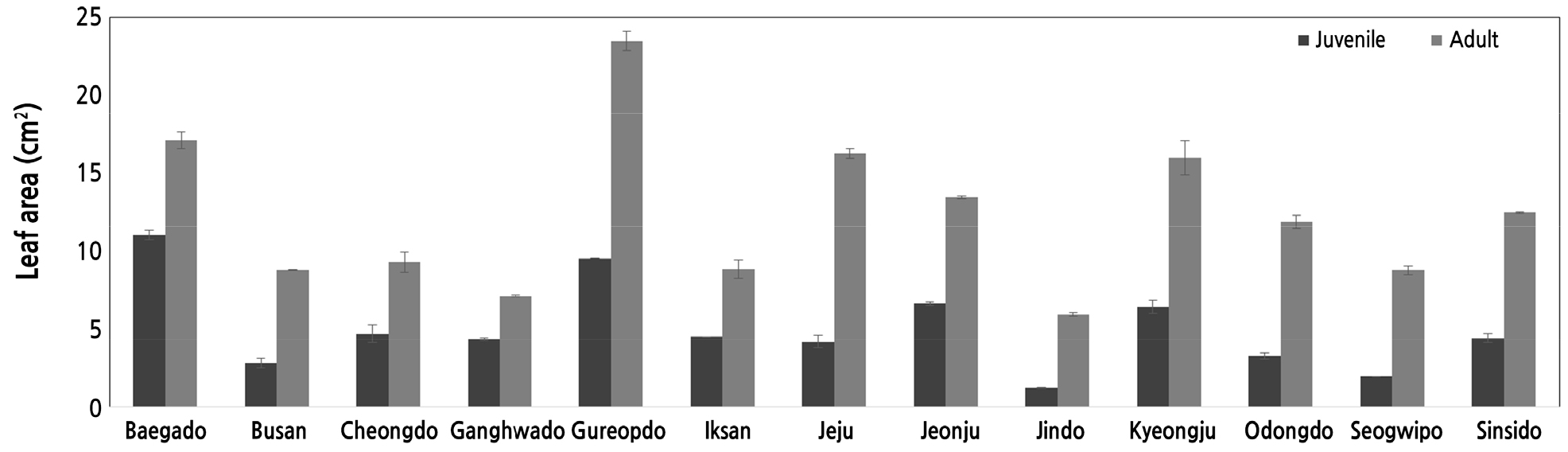
잎의 두께가 가장 두꺼운 것은 굴업도의 0.84mm, 가장 얇은 것이 진도 0.35mm였다. 그러나 이들 모두 성년 잎의 두께가 유년기 잎의 최고치 보다 커 조사한 모든 지역에서 성년형 잎이 두껍게 나타났지만 유년과 성년간 유의미한 차이가 나타나지 않아 잎의 크기에 비례하여 나타난 차이로 보이며 이러한 차이를 상변화 특성으로 보아 육안으로 구분하는 데에는 무리가 따를 것으로 생각되었다.
본 연구에서 자생마삭줄의 유 ․ 성년 잎을 특징짓는 요소로 줄기의 굴성변화와 엽맥무늬 여부, 엽폭과 엽장의 비율을 토대로 한 엽형지수, 그리고 유 ․ 성년의 상대적 엽면적과 두께를 조사하였는데 본 연구에서 재배시험의 공시재료로 사용된 마삭줄(T.jasminoides)과 달리 자생지 조사에서는 품종을 가리지 않고 국내에 자생하는 마삭줄을 대상으로 하였다. 그 결과 자생지에서 조사된 모든 개화한 개체에서 감는 줄기를 발견하였고 상변화에 따른 엽맥무늬의 변화와 엽면적의 상대적 크기의 변화도 본 연구의 재배시험 및 지역별 조사대상의 모든 개체에서 공통으로 나타났다. 그러나 지역별 엽형지수 조사시 3곳(익산, 서귀포, 제주)에서 예외가 발견된 것은 본 연구에서 엽형을 결정짓는 세부요소로 엽맥무늬, 엽면적, 엽형지수 등 3가지 요소를 조사하였는데 이중 엽형지수에서 예외적인 결과가 나타난 것으로 이는 환경적 요인 외에도 종내에 변이 등 유전적 요인이 작용한 것으로 여겨지나 자생마삭줄의 상변화에 공통으로 나타나는 특징으로 삼기 위해서는 엽형지수의 경우 추가적인 연구가 더 필요 할 것으로 판단되었다. 또한, 본 연구에서 나타난 결과를 보면 진도를 제외하고 서해상의 섬에서 잎이 크고 두꺼운 경향을 보였다. Park(2006)은 전국 11개 지역 자생 쑥의 특성을 조사한 바 해안에 가까운 전남 순천, 경기 강화에서 채취한 쑥의 엽폭과 엽장이 강원, 경북 등의 내륙 지방보다 크고 지역마다 엽의 형태가 차이가 있을 뿐만 아니라 Ca, Mg 등 무기염류 함량도 높다고 보고한 바 있다. 따라서 마삭줄도 바다 근처의 환경이 생육에 좋은 영향을 미친 것으로 추측된다. 또한 이러한 환경적 요인 외에 품종의 차이에 의한 유전적 형질이 발현한 결과임을 배제할 수 없다. 보다 자세한 원인을 밝히기 위해서는 일본처럼(Uemachi and Shimomura, 2013) 국내 마삭줄의 유전적 분포조사와 함께 추가적인 연구가 필요할 것으로 판단되었다. 이처럼 엽형과 면적 등에서 지역별 품종별 차이가 나타날 수 있지만 그럼에도 불구하고 본 연구에서는 품종을 가리지 않고 국내에 자생하는 마삭줄을 대상으로 유 ․ 성년형 잎과 줄기에 공통으로 나타나는 특징을 구명하여 상적변환을 인식할 수 있는 지표를 설정하고자 하였다. 본 연구의 결과 상적변환 전, 후의 줄기의 굴성 변화, 뚜렷한 엽맥무늬 여부, 상대적인 엽의 크기 등의 변화는 본 연구의 재배시험에 사용된 털마삭줄을 포함한 모든 조사에서 지역과 품종을 가리지 않고 유 ․ 성년을 구분할 수 있는 요소로 판명되었다. 또한 3곳에서 예외가 발견된 엽형지수의 경우 마삭줄의 유전적 분포를 함께 조사하여 품종별 형태 조사를 병행하면 보다 확실한 결과를 도출할 수 있을 것으로 사료된다.
본 연구를 통해 밝혀진 자생마삭줄의 상변화를 판별할 수 있는 특성(굴성, 엽맥무늬, 잎의크기 등)을 상변화의 지표로 활용하면 상향으로 감는 줄기가 나타나거나 엽맥이 흐려지며 큰잎이 나타나면 대부분의 자생마삭줄의 경우 상변화가 시작된 것으로 보아도 좋을 것으로 판단되었다. 이와 같이 유년과 성년형 잎의 무늬나 크기 등의 변화는 이미 아이비(Hedera helix) 등의 식물에서 보고된 바 있어서(Davies et al., 2018; Park, 2019), 유 ․ 성년의 뚜렷하게 구분되는 특성을 이용하면 분화재배 시 종자번식을 할 경우 아이비의 유식물처럼 쳐지는 줄기에 엽맥무늬가 선명한 관엽식물로 사용될 개체를 생산할 수 있고, 또한 부착하여 오르는 성질을 이용하면 담쟁이 덩굴처럼 벽면조경이나 아치 등의 장식에 이용할 수 있다. 벽면 조경시 유년상 마삭줄은 관상의 대상이 잎과 줄기인 반면 성숙상 마삭줄은 여기에 꽃과 향기를 더하고 위를 향하며 감는 줄기는 유년상과는 전혀 다른 양상을 보인다. 또한 유년상 마삭줄을 수목아래 지피식물로 식재 시 성년상이 나타나면 의도와는 반대로 감는 줄기가 발생하여 수목을 뒤덮을 수도 있다. 또한 본 연구에서 밝혀진 유년성 지속기간이 최소 5년이었던 결과를 감안하면 단시간에 개화된 마삭줄을 생산하여 출하하거나 꽃을 감상하기 위해서는 성년상 마삭줄을 이용한 삽목 등 영양번식이 반드시 필요하다. 이는 상변화가 일어난 줄기는 영양번식을 하면 지속적으로 개화 특성을 나타내기 때문이다(Park, 2019), 또한 개화시 상변화 초기에는 개화하는 줄기가 한정되고 개화수도 많지 않으므로 식용 꽃 생산 등 다량의 개화를 목적으로 재배 시에는 상변화 후에 충분한 시간이 경과하여 개화가 잘 이루어지는 개체의 개화줄기를 이용한 번식방법을 선택해야 한다고 본다. 따라서 마삭줄의 유년성 지속기간을 파악하고, 외형을 통해 유 ․ 성년을 판별할 수 있도록 본 연구에서 설정된 지표는 자생마삭줄의 연구나 재배 시 기초자료로 사용될 수 있을 것으로 사료된다.
이상의 결과를 종합해 보면 국내에 분포하는 마삭줄(T. jasminoides)의 유기는 최소 5년으로 추정되며 자생 마삭줄의 상변화시 품종에 관계없이 잎과 줄기에 공통으로 나타나는 특징으로 줄기의 굴성 변화, 잎맥무늬 여부, 잎의 크기 변화 등이 나타나며 이러한 특징은 자생 마삭줄의 상변화의 지표가 될 수 있을 것으로 판단되었다.
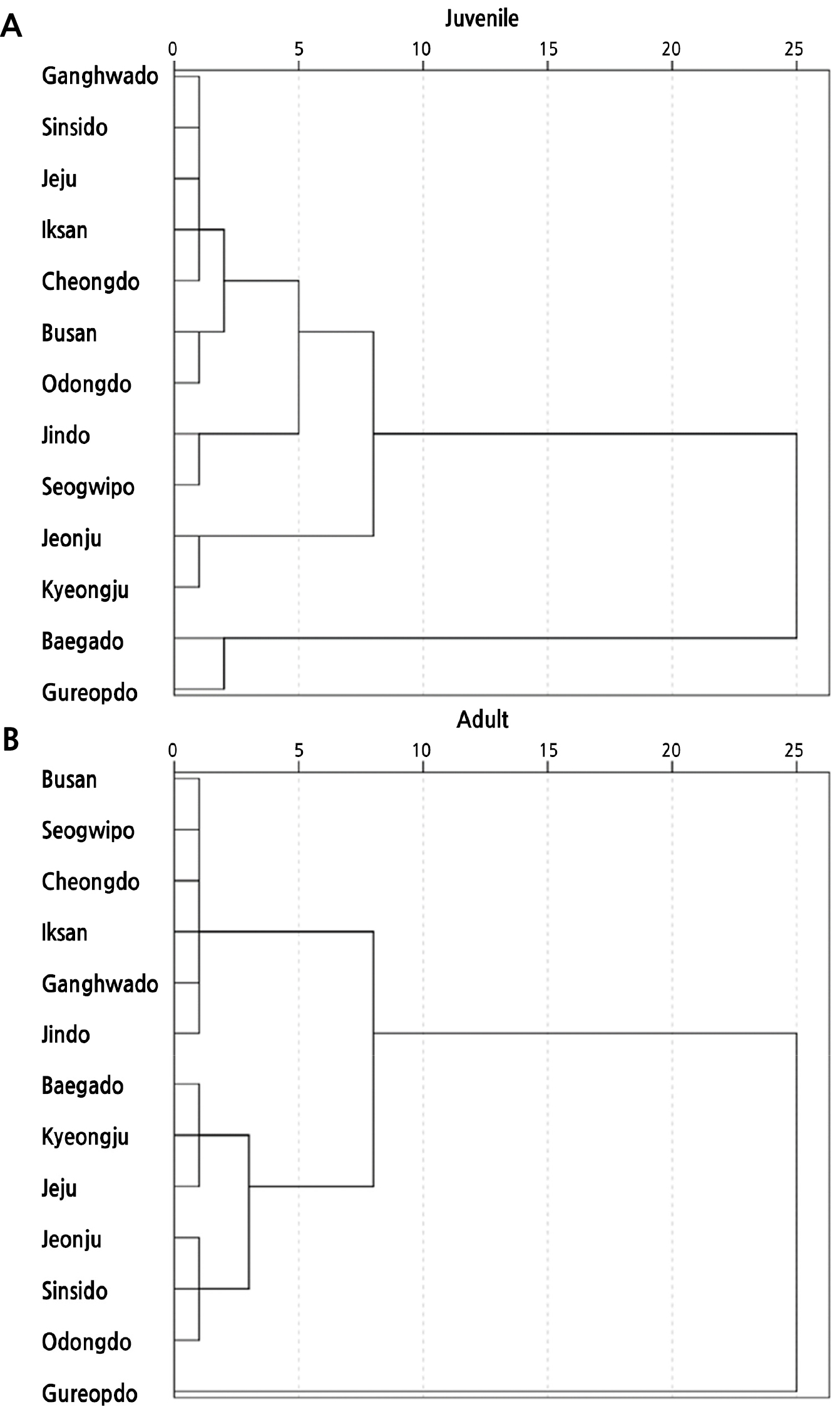
군집분석과 주성분 분석
채취 지역별 군집분석을 수지도(dendrogram)로 나타내었을 때, 재척도화 거리를 2.5로 군집화 하였을 시 유년기와 성년기 모두 4그룹으로 나누어졌는데 유년기의 경우 굴업도, 백아도가 1그룹, 청도, 부산, 익산, 강화도, 신시도, 제주, 오동도가 2그룹, 경주, 전주가 3그룹, 서귀포, 진도가 4그룹으로 분류되었다. 그리고 성년기는 경주, 백아도, 제주가 1그룹, 청도, 부산, 익산, 강화도, 서귀포, 진도가 2그룹, 전주 3그룹, 전주, 신시도, 오동도가 4그룹으로 분류되었다(Fig. 6). 지리적 집단 구분이 명확하지 않았던 팥배나무 열매의 형태적 특성(Kim et al., 2014; Kim et al., 2018)과 유사한 결과를 나타냈다. 채취 지역별 간의 구분이 명확하게 이루어지지 않은 것은 마삭줄 자생지의 기온, 습도, 그리고 토양 환경 차이 등에 의해 잎의 생장에 영향을 받아 유년기와 성년기 잎의 형태가 일정한 경향 없이 군집화되지 않은 것으로 판단된다. 또한 본 지역별 조사에서 품종을 가리지 않고 지역별로 개화한 1개체를 대상으로 유 ․ 성년의 형태적 차이를 비교 하였으나 각 지역의 자생지에는 품종이 혼재 해 있을 수 있고 종내에도 변이가 있을 수 있으므로 지리적 집단 구분이 명확하지 않았을 것으로 추측된다. 마삭줄 유년기와 성년기의 엽폭, 엽장, 엽면적, 엽두께, 엽형을 대상으로 주성분 분석(principal component analysis) 하였을 때, 제1주성분은 53.90%, 제2주성분은 17.01%을 보이며 축적 기여율은 70.91%이었다(Fig. 6). 제1주성분 중 유년기 엽면적, 성년기 엽면적, 제2주성분 중 성년기 엽형과 유년기 엽형이 높은 기여율을 보였다(Table 4). 주성분 분석의 경우 데이터 그룹의 편차 정도를 나타내는데(Bougrini et al., 2014), 유년기와 성년기 잎의 형태 요소가 유사한 위치에 놓이며 편차가 작은 것으로 나타났다.
Table 4.
Correlations of variables to the PCA factors based on factor loadings
| PC 1z | PC 2y | |
| Juvenile leaf width | 0.896 | –0.226 |
| Juvenile leaf length | 0.837 | 0.167 |
| Juvenile leaf shape index | 0.028 | 0.676 |
| Juvenile leaf area | 0.933 | –0.089 |
| Juvenile leaf thickness | 0.728 | –0.240 |
| Adult leaf width | 0.764 | 0.060 |
| Adult leaf length | 0.594 | 0.616 |
| Adult leaf shape index | –0.217 | 0.816 |
| Adult leaf area | 0.913 | 0.237 |
| Adult leaf thickness | 0.815 | –0.002 |
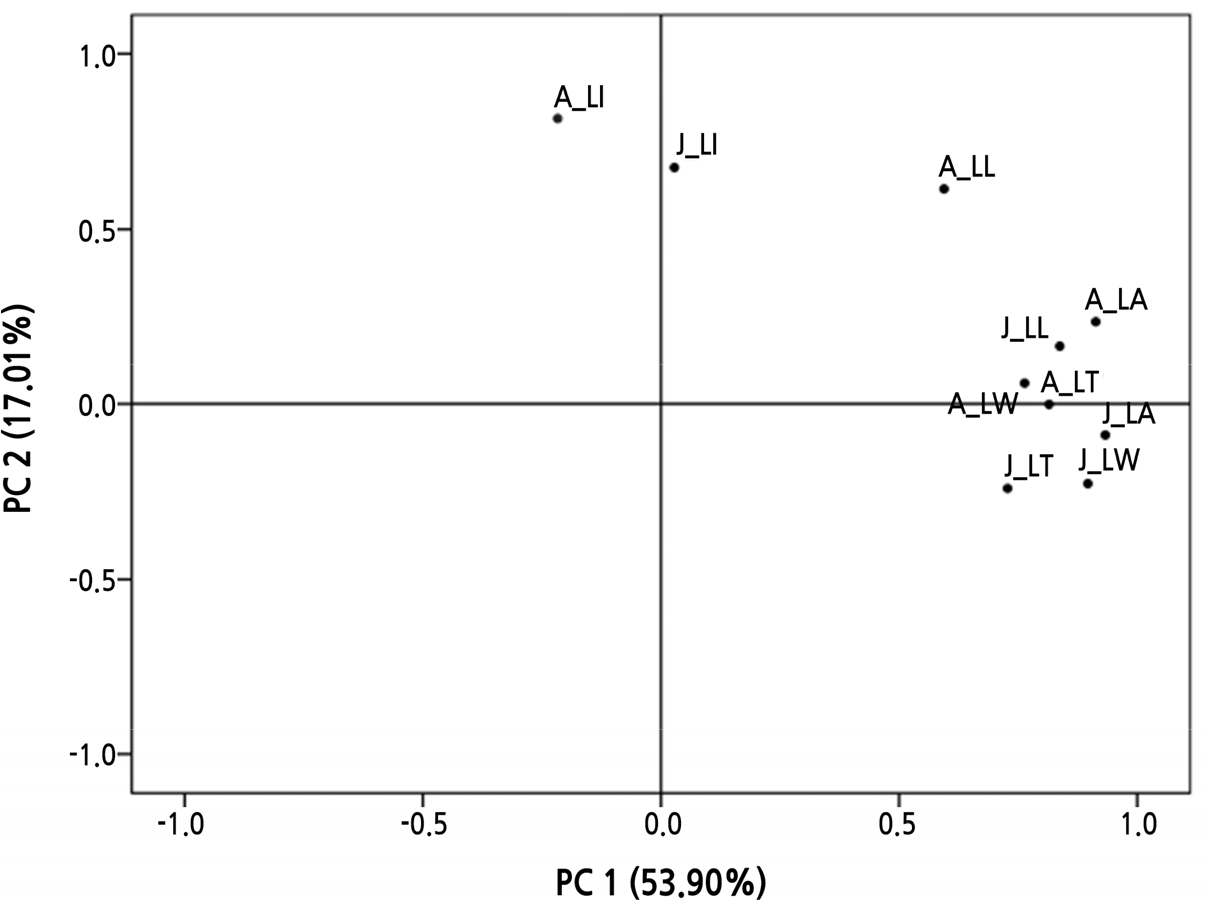
Fig. 6.
Plot of loadings for the response of the juvenile phase and adult phase of native Trachelospermum in Korea based on PCA. J_LW: leaf width of juvenile leaves, J_LL: leaf length of juvenile leaves, J_LI: leaf shape index of juvenile leaves, J_LA: leaf area of juvenile leaves, J_LT: leaf thickness of juvenile leaves, A_LW: leaf width of adult leaves, A_LL: leaf length of adult leaves, A_LI: leaf shape index of adult leaves, A_LA: leaf area of adult leaves, and A_LT: leaf thickness of adult leaves. PC 1: Mainly related to leaf width, leaf length, leaf shape index, lead area, and leaf thickness. PC 2: Related to leaf width, leaf length, leaf shape index, lead area, and leaf thickness.
군집분석 결과(Fig. 7), 마삭줄 유년기와 성년기 잎의 채취 지역에 따른 일정한 경향의 군집으로 분류되지 않았으나, 주성분 분석 결과 그래프 상 잎의 형태 요소(엽폭, 엽장, 엽형지수, 엽면적, 그리고 엽두께)가 놓인 위치가 서로 가까운 곳에 위치하며 편차가 작아 마삭줄 유년기와 성년기 잎의 형태적 주성분 요소는 유사하였다. 또한 주성분 분석 결과 제1주성분 중 가장 높은 기여율을 보인 엽면적이 자생 마삭줄의 성년상을 구분하는 지표가 될 수 있음을 알 수 있었다.