

Introduction
Materials and Methods
Plant Materials
Assessments of Fruit Quality Attributes
Statistical Analysis
Results
Discussion
Introduction
Temperate and tropical fruits can be categorized as climacteric or non-climacteric, depending on the differences in ethylene production and respiration during fruit ripening and softening (McMurchie et al., 1972). For example, apple fruit are climacteric because ethylene production coincides with a rise in respiration during fruit ripening (Knee, 1971; Yang and Hoffman, 1984). Dandekar et al. (2004) studied the physiological roles of ethylene during fruit ripening by using transgenic apples silenced for the genes encoding either 1-aminocyclopropane-1-carboxylic acid (ACC) synthase (ACS) or ACC oxidase (ACO) and observed a reduction in ethylene production rate and the maintenance of flesh firmness and peel color during storage at 20°C, compared with the wild-type ‘Greensleeves’ apple (Malus × domestica Borkh.). By contrast, melon (Cucumis melo L.) fruit show either climacteric or non-climacteric ripening for ethylene production and respiration rate (Moreno et al., 2008; Périn et al., 2002). In addition, Costa et al. (2005) reported that, for apples, the progeny of a ‘Fuji’ × ‘Braeburn’ cross showed a wide range of ethylene production during shelf-life; their results suggest that some progeny might be non-climacteric due to low levels of ethylene production.
During fruit maturation and ripening in apples, numerous biochemical pathways can be independently or dependently associated with the rise in ethylene production. Therefore, ethylene production during fruit ripening can influence many fruit quality attributes including fruit texture, flavor, peel and flesh tissue color, aroma, acidity, and sweetness. However, the conversion of starch into soluble carbohydrates is mutually exclusive with an ethylene burst during fruit ripening. Nevertheless, internal ethylene concentration (IEC) is correlated with alterations in starch pattern index; a negative correlation with starch concentration and amylose content in ‘Gala’, ‘Honeycrisp’, ‘McIntosh’, and ‘Empire’ apples has been reported (Doerflinger et al., 2015). On the other hand, alterations in fruit firmness are strongly associated with an increase in ethylene production during storage (Watkins et al., 2000). However, the relationship between IEC and flesh firmness differed between ‘Ambrosia’ and ‘Empire’ apples (Ehsani-Moghaddam and DeEll, 2009); IEC was negatively correlated with flesh firmness, SSC, and TA in ‘Empire’ apples, but only negatively associated with SSC in ‘Ambrosia’ apples.
The changes in these fruit quality attributes during shelf life and cold storage are highly affected by harvest maturity, which is in turn linked to harvest time. Advanced maturity at harvest resulted in a higher accumulation of SSC and a greater change in flesh firmness during storage in ‘Reinette de Canada’ apples (Guerra and Casquero, 2011). A late harvest time is strongly associated with loss of flesh firmness not only at harvest but also during and after storage in both ‘Gala’ (Jobling and McGlasson, 1995) and ‘Honeycrisp’ (Wargo and Watkins, 2004) apples. However, a later harvest time did not influence SSC in ‘Honeycrisp’ apples (Wargo and Watkins, 2004). The differences in ethylene production differentially affected loss of fruit firmness during storage in the progeny of the ‘Fuji’ × ‘Braeburn’ population (Costa et al., 2005). Furthermore, the physiological alteration of fruit quality attributes, such as IEC, flesh firmness, and fruit respiration, was more dramatic in early ripening summer cultivars including ‘Sunrise’ (Lau and Lane, 1998) and ‘Anna’ (Klein and Lurie, 1990) apples than in typical fall cultivars, such as ‘Fuji’(Watkins et al., 1989).
Several apple cultivars newly released from the Korean apple breeding program showed low IEC at harvest as well as during shelf life and cold storage (personal communication). Therefore, the objective of the current research was to characterize IEC and fruit quality attributes of ‘Summer Dream’, ‘Summer King’, ‘Green Ball’, ‘Picnic’, and ‘Hwangok’ apples harvested at three different harvest times then held at 20°C for shelf-life evaluation or one day after cold storage (0.5°C for up to 40 da ys).
Materials and Methods
Plant Materials
Apple (Malus × domestica Borkh.) fruit used in these experiments were harvested from ‘Summer Dream’/M.26, ‘Summer King’/M.9, ‘Green Ball’/M.9, ‘Picnic’/M.9, and ‘Hwangok’/M.9 trees (Ban et al., 2014) grown at the experimental orchard of the Apple Research Institute, National Institute of Horticultural & Herbal Science, RDA, in Kunwi, Republic of Korea (Latitude: 36°16'6; Longitude: 128°28'11; Elevation: 67 m). Three 10-year-old ‘Summer Dream’, three 6-year-old ‘Summer King’, ‘Picnic’, and ‘Hwangok’, and three 8-year-old ‘Green Ball’ apple trees were selected. Fruit were sampled on August 1, 6, and 11, 2012 for ‘Summer Dream’, and ‘Summer King’; on August 24 and September 3 and 13, 2012 for ‘Green Ball’; on September 9, 19, and 29, 2012 for ‘Picnic’; and September 13 and 23, and October 3, 2012 for ‘Hwangok’. Fruit were harvested randomly from the perimeter of each sample tree. Uniformly sized fruit of each cultivar were selected to provide two experimental units for both shelf life evaluation and cold storage studies, which were carried out at the postharvest laboratory at the Kyungpook Naotinal University campus in Daegu, Republic of Korea.
Expt. 1. Eight fruit for ‘Summer Dream’, ‘Summer King’, ‘Green Ball’, ‘Picnic’, and ‘Hwangok’ cultivars with three different harvest times were used for the daily measurement of IEC during shelfif el at 20°C.
Expt. 2. Fruit from each cultivar were stored in air at 0.5°C with 90% relative humidity. Fruit quality attributes were evaluated at 0, 20, 30, and 40 days of cold storage. IEC, flesh firmness, soluble solids concentration (SSC), and titratable acidity (TA) were evaluated from five fruit per cultivar one day after removal from cold storage.
Assessments of Fruit Quality Attributes
For IEC sampling for shelf life study, a stainless steel 18 gauge 3.8 cm long needle with a plastic luer lock was placed into the core cavity through the calyx end of the apple and then sealed with adhesive poster putty to prevent any leakage of gas during the measurement. For the cold storage study, fruit were directly used for IEC sampling one day after removal from cold storage and IEC was determined by withdrawing a 1 mL gas sample from the core cavity of each apple fruit. The sample was injected into a gas chromatograph (GC2010, Shimadzu, Tokyo, Japan), equipped with a flame ionization detector and fitted with a Porapak Q (80/100 2 m, Young In Frontier, Seoul, Korea) column. Analyses were run isothermally with an oven temperature of 90°C and injector and detector temperatures of 100°C and 200°C, respectively. The flow rate of He was 25 mL·min-1. Ethylene was quantified by peak area using an external standard for calibration.
Flesh firmness was assessed on the same fruit by taking two readings at pared regions of the equator of each apple with a penetrometer (Fruit Pressure Tester, FT 327, Alfonsine, Italy). Juice expressed from the penetrometer readings was used for soluble solids concentration (SSC) measurement with a refractometer (Model PR-201α, Atago Co., LTD., Tokyo, Japan) and for titratable acidity (TA) determination by titrating with 0.1 N NaOH to pH 8.1.
Statistical Analysis
Data were subjected to analysis of variance (ANOVA) to determine main effects and interactions (Version 9.3; SAS Institute Inc., Carry, NC, USA). Pearson correlation coefficients were obtained using the PROC CORR procedure in SAS version 9.3 to test association between fruit quality attributes from 5 apple cultivars during cold storage. The correlation analysis was performed with five replications (n = 5). Asterisks (*, **, ***, or ****) indicate a statistically significant correlation at p < 0.05, 0.01, 0.001, or 0.0001, respectively.
Results
We used 8 fruit for Expt. 1 and 5 fruit for Expt. 2 because the total number of uniform unblemished fruit was not enough to support a larger sample size, typically 18 to 30 fruit per treatment.
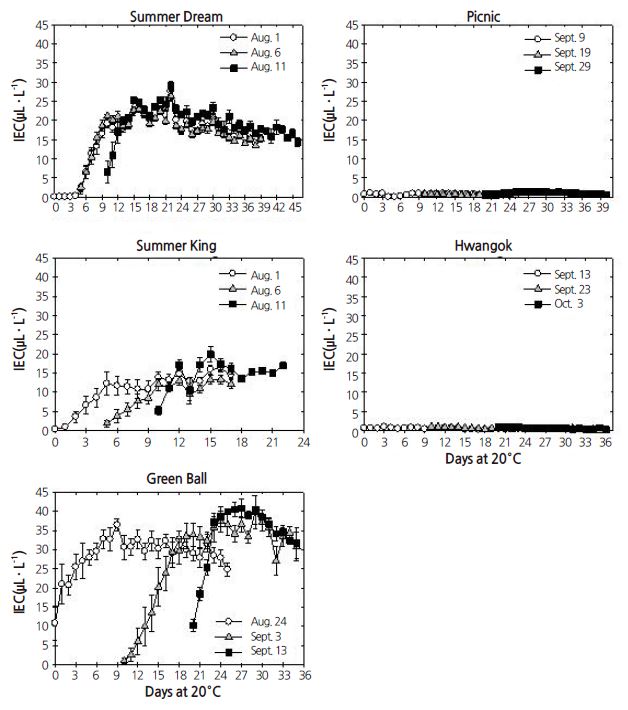
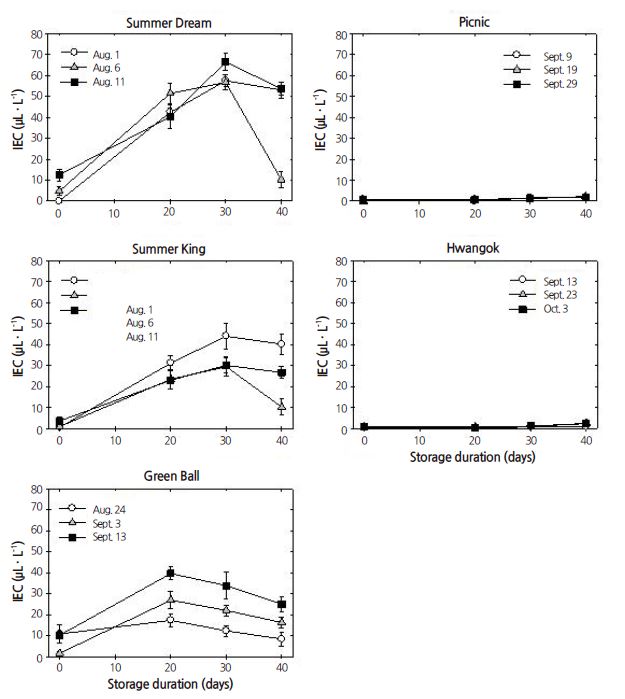
Expt. 1. The IEC patterns for ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’ responded similarly during shelf life at 20°C, compared with those in ‘Picnic’ and ‘Hwangok’, although the responsiveness of IEC to apple cultivar and harvest time were slightly different (Fig. 1). IEC was highest for ‘Green Ball’, 30-40 μL·L-1, followed by ‘Summer Dream’, 15-25 μL·L-1, and ‘Summer King’, 10-20 μL·L-1. In contrast, IEC levels for ‘Picnic’ and ‘Hwangok’ were less than 1 μL·L-1 during shelf life at 20°C, irrespective of harvest time.
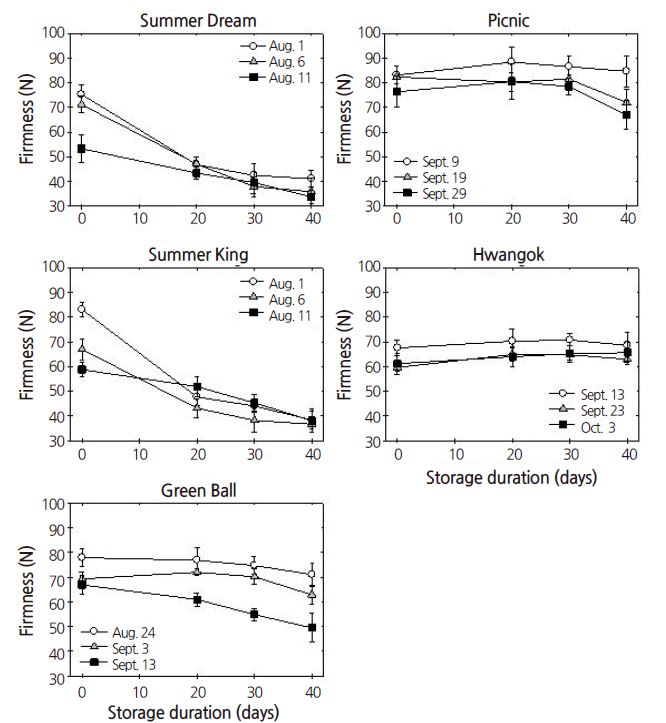
Expt. 2. Fruit flesh firmness decreased as the harvest time advanced. Firmness loss occurred in ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’ during cold storage but was relatively minor in ‘Picnic’ and ‘Hwangok’ (Fig. 2). Flesh firmness was significantly affected not only by the main effects, which are cultivar, harvest time, and storage period, but also by the two- and three-way interaction effects (Table 1).
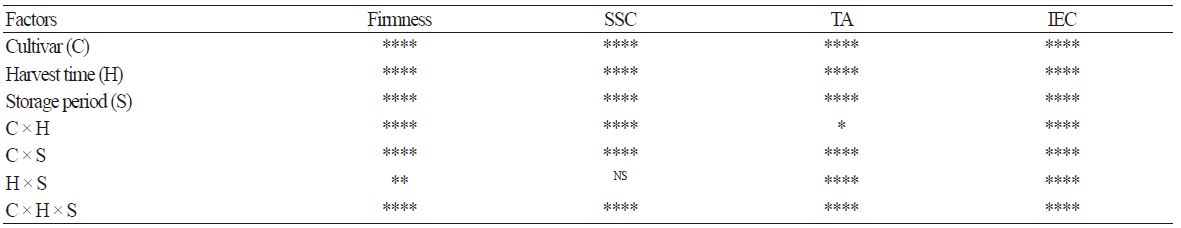
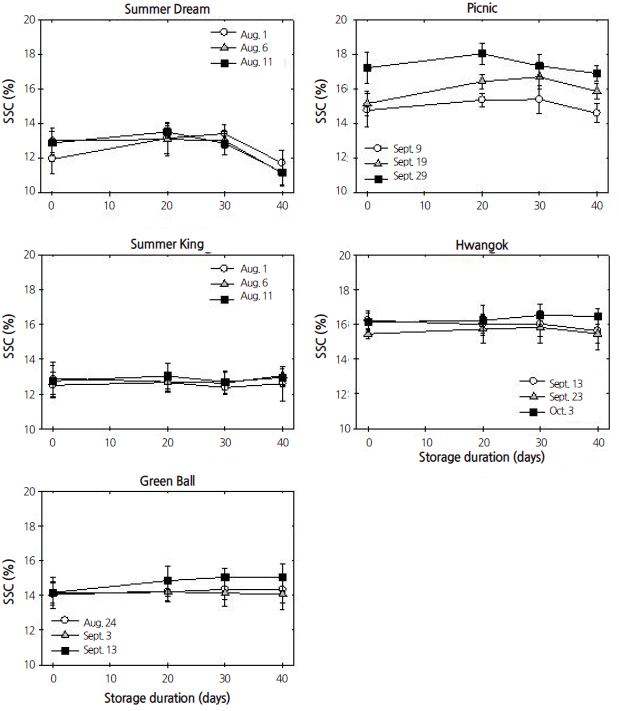
Although harvest time did not affect SSC in ‘Summer Dream’, ‘Summer King’, ‘Green Ball’, and ‘Hwangok’, SSC was highest in the late-harvested fruit of ‘Picnic’ during cold storage. SSC was not strongly influenced by cold storage except in ‘Summer Dream’, where SSC decreased with storage period. SSC was significantly affected by the main effects, cultivar, harvest time, and storage period, as well as by two- and three-way interactions, except for the two-way interaction between harvest time and storage period (Table 1).
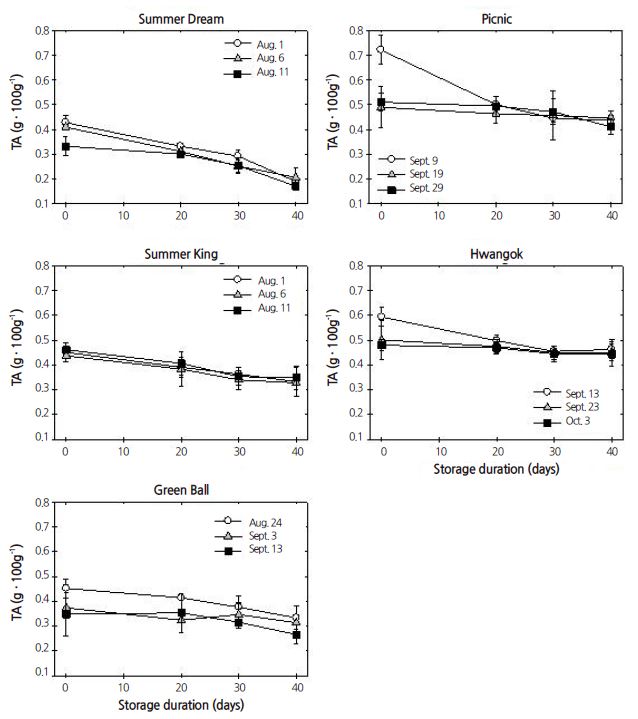
TA gradually declined or remained unchanged during cold storage, regardless of harvest time or cultivar. The decrease in TA was highest in ‘Summer Dream’ during 40 days of cold storage but inconsistent in ‘Picnic’ and ‘Hwangok’ throughout cold storage (Fig. 4). Nonetheless, the levels of TA were strongly affected by both main and interaction effects (Table 1).
Of all the parameters assessed, IEC displayed the most distinctive difference among the five apple cultivars during cold storage (Fig. 5). The pattern of IEC levels could be clearly divided into two groups. IEC in three cultivars, ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’, increased during cold storage, regardless of harvest time but remained low in ‘Picnic’ and ‘Hwangok’. In these two cultivars, the IEC level was less than 4 μL·L-1 during the whole cold storage period. Furthermore, IEC level was the attribute that was most significantly influenced by all the maina nd interaction effects (Table1).
Based on the results of a correlation coefficient test, flesh firmness positively correlated with TA in ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’ but negatively correlated with IEC in all cultivars, except in ‘Green Ball’, at different levels of significance (Fig. 6). SSC was negatively correlated with flesh firmness but positively associated with IEC in ‘Green Ball’. TA was negatively correlated with IEC in ‘Summer Dream’ and ‘Summer King’. In addition, the number of statistically significant relationships between variables was much higher in ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’ cultivars than in ‘Picnic’ and ‘Hwangok’ cultivars.
Discussion
Apple fruit ripening has a climacteric pattern due to the increase in ethylene production and CO2 evolution during ripening, although some progeny of the cross between ‘Fuji’ and ‘Braeburn’ apples showed distinctively low ethylene production during shelf life (Costa et al., 2005). In the current study, two cultivars, ‘Picnic’ and ‘Hwangok’, also showed low IEC during shelf life and cold storage, compared with the other three cultivars. The IEC results during shelf life at ambient temperature indicated that the IEC response of ‘Picnic’ and ‘Hwangok’ distinctly differed from the response of the other three cultivars, ‘Summer Dream’, ‘Summer King’, and ‘Green Ball’. In commercially well-known apples, ethylene production varies among cultivars not only during shelf life, but also during cold storage (Costa et al., 2010; Jobling and McGlasson, 1995; Tatsuki et al., 2007; Watkins et al., 1989). Apples that produce low amounts of ethylene showed less of a reduction in fruit firmness during either controlled atmosphere storage or cold storage followed by three weeks of storage at room temperature (as for shelf life), compared with apples that produce high amounts of ethylene (Colgan et al., 2006). However, two summer apple cultivars, ‘Sunrise’ and ‘Silken’, had a rapid reduction of flesh firmness after short-term cold or CA storage, compared with fruit firmness at harvest (Lau and Lane, 1998; Lu and Toivonen, 2010; Toivonen and Lu, 2005). Therefore, the different responses to IEC levels are likely cultivar-specific or dependent. The results presented here support the hypothesis that apple fruit might have two different types of ripening patterns based on the responsiveness to ethylene production, regardless of whether the fruit are stored in shelf conditions or cold storage.

Harvest time significantly influenced IEC responsiveness during both shelf life and cold storage. Advanced harvest time also contributed to an increase in the IEC responses in two summer apples (Lu and Toivonen, 2010; Toivonen and Lu, 2005), thereby decreasing fruit firmness during short-term cold or controlled atmosphere storage (Lau and Lane, 1998; Toivonen and Lu, 2005). Higher IEC was linked to a reduction in flesh firmness at harvest as the harvest time was advanced (Bowen and Watkins, 1997; Jobling and McGlasson, 1995; Nock and Watkins, 2013). Results for the new cultivars presented here indicate that IEC is negatively correlated with flesh firmness during cold storage for all cultivars except ‘Green Ball’. Regardless of harvest season, the negative relationship between IEC and flesh firmness typically appears during storage (Tahir and Nybom, 2013; Watkins et al., 2000). The effect of harvest time on IEC appears to be cultivar specific, as IEC is not influenced by harvest time in ‘Picnic’ and ‘Hwangok’.
Our results indicated that, of the fruit quality attributes, the reduction of flesh firmness during cold storage was cultivar specific, regardless of harvest time. Flesh firmness gradually declined during cold storage of ‘Summer Dream’ and ‘Summer King’ but the loss of flesh firmness was reduced in ‘Picnic’ and ‘Hwangok’, compared with the other cultivars. It is likely that the response of flesh firmness to cold storage in ‘Green Ball’ does not belong to either the ‘Summer Dream’ and ‘Summer King’ group or the ‘Picnic’ and ‘Hwangok’ group due to an inconsistent response of flesh firmness during cold storage. On the one hand, numerous apple genotypes show a wide range of flesh firmness at harvest and during storage (Iwanami et al., 2008a; Iwanami et al., 2008b; Oraguzie et al., 2007). On the other hand, lower rates of loss of flesh firmness are highly associated with low ethylene producing characteristics during cold or CA storage (Colgan et al., 2006). The reduction in flesh firmness is sharp in ‘Akane’ and ‘Silken’ but less in ‘Fuji’ (Iwanami et al., 2008b). Summer apple cultivars, such as ‘Sunrise’, and ‘Anna’, had a great reduction of flesh firmness not only due to advanced harvest time but also during short-term cold or controlled atmosphere storage (Lau and Lane, 1998; Pre-Aymard et al., 2005; Pre-Aymard et al., 2003; Toivonen and Lu, 2005). In contrast, as a late season apple cultivar, ‘Fuji’ showed less fluctuation in IEC levels within a wide range of fruit maturity based on blush degree and background color category (Watkins et al., 1993). In addition, ethylene production was greatly reduced in late season apple cultivars, ‘Fuji’ and ‘Granny Smith’ than in early and middle season apple cultivars, ‘Cox’s Orange Pippin’, ‘Royal Gala’, ‘Braeburn’, and ‘Delicious’ strains (Watkins et al., 1989). The results of Iwanami et al. (2008a; 2005) indicate that there are diverse patterns of flesh firmness loss during storage.
In contrast to IEC, the response of SSC was inconsistent. SSC gradually declined over storage period, regardless of cultivar (Kolniak-Ostek et al., 2014). In the present study, fruit were only stored for up to 40 days in air. It is assumed that this time period is not long enough to reveal any consistent patterns in SSC response as compared with SSC results from long-term storage. Fruit harvested at later times tended to have a higher level of SSC in ‘Green Ball’ and ‘Picnic’. Otherwise, SSC gradually declined in ‘Summer Dream’, irrespective of harvest time. In addition, TA gradually decreased during cold storage regardless of harvest time but did so inconsistently in ‘Picnic’ and ‘Hwangok’. Thus, the inconsistent responses in SSC and TA levels led to no significant correlation between SSC and TA except in ‘Summer Dream’. The correlation pattern was remarkably different in ‘Summer Dream’ and ‘Summer King’ compared with ‘Picnic’ and ‘Hwangok’ based on the number and significance of the correlations. This result indirectly reveals that these two types of cultivars might have different ripening or softening mechanisms.
In conclusion, several newly bred apple cultivars have been released with different physiological IEC patterns at harvest and during cold storage. Alongside the distinctive IEC patterns, other fruit quality attributes including SSC, TA, and flesh firmness also varied with cultivar. Of these cultivars, the physiological responses in ‘Picnic’ and ‘Hwangok’ are highly different from those in ‘Summer Dream’ and ‘Summer King’, with less change in IEC and firmness during shelf life and cold storage. The correlation results also show that these two groups had a diverse correlation response during cold storage. Therefore, the results indicated that these two groups of apple cultivars might have physiologically different mechanisms for fruit ripening and softening
