

서 언
여름 시설 재배는 온실 내로 방사된 복사열로 인해 외부보다 온실 내부의 온도가 높아져 작물 적정 온도 관리에 어려움이 많다. 시설 내에서는 주로 환기 또는 차광을 단독 또는 병행 처리하거나 증발 냉각, 지붕 살수 등을 이용하여 고온을 극복하고 있다. 최근 규모화된 온실에서는 유공 덕트와 열 교환기를 이용하여 외기를 흡입한 후 각 덕트의 구멍으로부터 저온 건조한 공기가 뿜어 나오는 구조를 이용한 강제 환기 방식(Na et al., 2011), 미세노즐을 이용하여 세무하는 증발냉각 방식(Lee and Kim, 2011) 등을 통해 지상부 고온을 하강시켜 안정적인 생산을 꾀하고 있다.
시설 내 온도를 하강시키는 냉방은 설치에 따른 초기 비용과 에너지 소모량에 따른 운영 비중이 높아 작물 생육에 적합한 온도로 온실 환경을 유지하기에는 한계가 있다. 따라서 야간에만 냉방을 실시하거나, 온도 하강이 필요한 부분에 부분적으로 냉방하는 국소냉방을 이용하는 방식이 온실 전체를 냉방하는 것보다 더 경제적일 수 있다(Lee et al., 2002; Morgan, 2011).
지하부 근권 냉방은 온실의 지상부 냉방에 소요되는 에너지 비용을 줄이면서 뿌리가 분포하는 배지 온도 상승을 낮추고, 뿌리의 생장을 촉진시키고, 양・수분 흡수 저해를 완화시켜 작물 생육과 수량에 긍정적임을 파프리카(Jang et al., 2010), 토마토(Lee et al., 2002), 오이(Moon et al., 2007), 엽채류(He et al., 2001) 등에서 보고(Morgan, 2011)된 바 있다. 이와 같은 근권 냉방 방식은 급액 온도를 낮추거나(Kim et al., 2011), 배지부 열 교환 매체나 증발 냉각을 이용하는 방식(Kim et al., 2010) 등이 사용되었다.
따라서 본 실험은 고온기 파프리카 수경재배에서 XL 파이프를 이용한 냉수 순환에 의한 근권 냉방의 냉방 시간이 배지 내 온도 변화와 파프리카의 뿌리 활력, 엽 수분포텐셜 및 착과에 미치는 영향을 알아보고자 수행하였다.
재료 및 방법
공시 품종은 Blocky 타입의 주황색계 파프리카(Capsicum annum. L) ‘Orange glory’(Seminis Vegetable Seed Inc., USA)이다. 암면 블록에 육묘(초장 40cm 내외)된 파프리카를 2012년 7월 16일 서울시립대 양지붕 단동 유리온실(N37.5°, E127.0°)에 정식하여 2줄기 유인하여 10월 15일까지 수경재배하였다. 정식용 배지는 유기 코코넛 코이어 슬라브(chip:dust = 5:5(v/v), W 20 × L 100 × H 12cm, Shinsung Minerals Co., Sri Lanka)이며, 배지의 화학적 특성과 조제 과정에서 집적된 염을 제거하고자 3회 이상 세척하였고, 파프리카 네덜란드 배양액(EC 2.0dS・m-1)으로 충분히 포수한 후 슬라브당 3주씩 파프리카를 정식하였다. 파프리카 수경재배 방식은 비순환식 시스템이며, 작물 생육에 따라 배양액 공급 EC 2.7-3.5dS・m-1로 조절하면서 pH 5.7 ± 0.2로 공급하였다. 1회 배양액 공급량은 주당 100-110mL씩 하루 8-9회, 일출 후 2시간부터 일몰 1시간 전까지 24시간 타이머로 생육 초기 공급하다가 점차 일일 급액량을 증가시켰다.
근권 냉방은 코이어 배지와 스티로폼 베드 사이에 XL 파이프를 매설하고, 자동온도조절기(DOV-882, Daeil Co., Ltd., Korea)와 냉각기(DA-3000B, Daeil Co., Ltd., Korea)를 사용하여 설정 온도 ± 0.5°C에서 제어가 가능하도록 하였으며, 냉수 설정온도는 20 ± 2°C로 유지하면서 순환시키는 냉수 순환형 방식을 채택하였다. 근권 냉방 효과를 알아보고자 냉방 공급 시간에 따라 연속냉방(전일, 24시간), 야간냉방(야간, 오후 5시-오전 3시) 및 냉방무처리(대조구)의 3수준으로 7월 23일부터 9월 23일까지 처리하였다.
온실 내 온도는 대기 온습도센서(SHT-110, Miraesensor Co., Korea)로 10분마다 계측하였고, 배지 온도는 FDR 센서(CoCo Sensor, Miraesensor Co., Korea)를 이용하여 작물과 작물 사이에 센서를 상부에서 5-7cm 깊이에 꽂아 10분 간격으로 계측한 후 Data logger(WP700, Miraesensor Co., Korea)에 자료를 수집하였다.
처리에 따른 배지 온도 변화는 7월 23일부터 8월 31일까지 수집된 자료를 최고, 평균, 최저 온도로 각각 정리하였고, 하루 중 배지 온도 변화는 맑은 날(일사량 600-700W・m2, 8월 1일)과 흐린 날(일사량 < 350W・m2, 8월 13일)에 계측된 자료를 주간(오전 6시-오후 8시), 야간(오후 8시-오전 6시)으로 각각 분석하였다.
배지 깊이에 따른 배지 온도는 배지 하부 기준으로 8cm(상), 5cm(중), 2cm(하)로 나누어 CR10X data logger(Campbell Scientific, USA)로 8월 1일, 10분 간격으로 수집한 자료를 시간별 평균값으로 분석하였다.
근권 냉방에 의한 파프리카의 생리적 반응을 알아보기 위하여 뿌리활력과 수분포텐셜을 측정하였다. 뿌리 활력은 Hirata (1990)의 triphenyl tetrazolium chloride(TTC)법으로 측정하였다. 처리 25일째 1cm 내외로 균일하게 절단한 0.5g 뿌리를 1% TTC가 혼합된 용액에 침지한 후 환원 반응에 따른 적색 정도를 분광광도계(Shimadzu, UV-1800, Japan) 470nm에서 흡광도를 측정하여 근활력을 계산하였다. 잎 수분퍼텐셜은 근권냉방 43일째 오전 9시부터 3시간 간격으로 다음날 오전 6시까지 상부에서 3번째 마디의 전개 잎을 채취하여 휴대용 잎 수분포텐셜 측정기(SKPM 1400, SKYE, UK)로 질소 가스 압력에 의해 수액이 엽병 밖으로 나오는 시점의 값을 3회 반복 측정하였다. 파프리카 첫 개화가 시작된 시기는 육안 조사하였고, 주당 착과수는 처리 63일과 77일에 조사하였다.
통계분석은 SAS package(statistical analysis system, version 9.1, SAS Institute Inc.)를 이용하여 Duncan’s multiple range test(DMRT)를 실시하여 5% 유의수준에서 각 처리간 차이의 유의성을 검정하였다.
결과 및 고찰
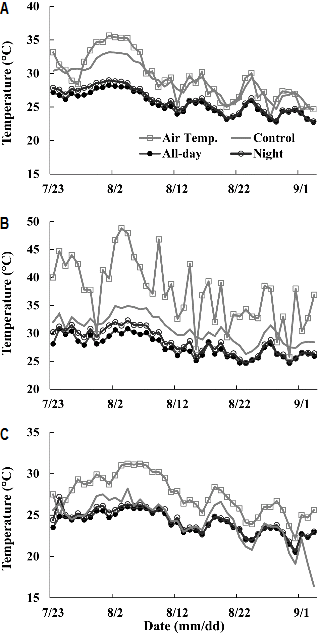
고온기(7월 23일부터 8월 31일까지) XL 파이프를 이용한 근권 냉방 방식에서 냉방 시간에 따른 배지 내 일평균, 최고, 최저 온도 변화는 대조구보다 1.8-5°C 낮아 온도 하강 효과가 관찰되었다(Fig. 1). 근권 배지의 평균온도가 전일 처리구에서는 25.6°C(22.7-28.2°C), 야간 처리구는 26.1°C(22.9-29.2°C)였으며, 대조구는 29.1°C(24.7-33.2°C)였다. 근권 냉방에 의한 배지 온도 하강이 전일 처리구에서는 평균 3.5°C(1.9-5.0°C), 야간 처리구에서는 평균 3°C(1.8-4.0°C)로 전일과 야간 냉방 시간 처리간 일 평균 배지 온도 차이는 0.5°C였다. 고온기간 중 8월 1일부터 10일까지 온실 평균 온도가 32.7°C(29.0- 35.6°C)이였을 때 전일 처리구의 배지 온도는 24.8-28.1°C로 대조구 배지 온도(28.1-33.2°C)에 비해 온도 하강(3.3-5.1°C) 효과가 크게 나타났다(Fig. 1A). 전일 처리구, 야간 처리구 및 대조구 배지의 최고온도는 27.6°C(24.7-30.8°C), 28.5°C (24.7-32.3°C), 30.8°C(26.3-34.9°C)였고, 최저온도는 24.0°C (20.5-26.0°C), 24.3°C(20.6-26.3°C), 27.4°C(24.1-31.2°C)였다. 이는 배지의 온도범위가 전일, 야간 처리구 및 대조구에서 각각 20.5-30.8°C, 20.6-32.3°C, 24.1-34.9°C를 나타냄으로써 근권 냉방 처리와 처리시간이 길었을 때 대조구보다 근권 온도는 낮아졌다(Figs. 1B and 1C).
광량이 많았던 맑은 날(일사량 최고 600-700W・m-2)과 광량이 적었던 흐린 날(일사량 < 300W・m-2)의 하루 중 배지 온도 변화를 측정한 결과는 Fig. 2와 같았다. 맑은 날 온실 내부 온도가 최고조에 도달한 시간은 오후 2시경이였으며, 배지 온도가 최고조에 도달한 시간은 전일과 야간 처리구에서는 오후 4-5시, 대조구에서는 오후 7-8시였다. 냉방 처리를 받은 배지 온도는 일몰(오후 7시) 이후 배지 온도가 감소한 반면, 대조구의 오후 8시 이후부터 오전 6시 전까지 배지 온도는 온실 온도보다 높았다. 이는 냉수 순환을 위해 스티로폼 베드 위에 슬라브를 설치함에 따라 스티로폼 재질의 특성상 주간에 상승한 배지 온도가 일몰 이후에도 낮아지지 못한 것으로 생각된다.
맑은 날 하루 평균 배지 온도가 전일 처리구에서는 27.1°C, 야간 처리구에서는 28.1°C, 대조구는 30.8°C로 근권 냉방으로 배지 온도는 2.7(야간)-3.7°C(전일) 낮아졌다(Fig. 2A). 주간(오전 6시-오후 8시)의 온실 평균온도는 37.3°C였으며, 처리구의 배지 평균온도는 각각 28.0°C(전일), 29.2°C(야간), 31.3°C(대조구)였다. 야간(오후 8시-오전 6시) 온실 평균온도는 28.7°C였으며, 배지 평균온도는 26.3°C(전일), 26.9°C (야간), 30.3°C(대조구)였다(자료 미제시). 냉방처리로 주간/야간 배지온도가 대조구보다 전일 처리구에서는 3.3°C/4°C, 야간 처리구에서는 2.1°C/3.4°C 온도가 하강하였고, 주간보다 야간 온도 하강이 컸다. 특히 야간 처리구는 배지의 야간 온도 하강으로 주간 배지 온도를 낮추었다. 전일과 야간 처리 간 배지온도 차는 오후 4시에 최고 1.9°C였으며, 주간은 평균 1.2°C, 야간은 평균 0.6°C 차이를 나타냈다.
흐린 날의 배지 온도 변화도 맑은 날과 같은 경향을 보였으며, 근권 냉방으로 일평균 2.9(야간)-3.3°C(전일) 배지 온도가 하강했다(Fig. 2B). 야간 처리구는 맑은 날에 비해 주간 배지 온도 상승이 높지 않은 반면, 야간 시간 냉방으로 인해 주간 배지 온도 하강이 더 컸으며, 전일과 야간 처리에 따른 온도 차이는 맑은 날에 비해 작았다.
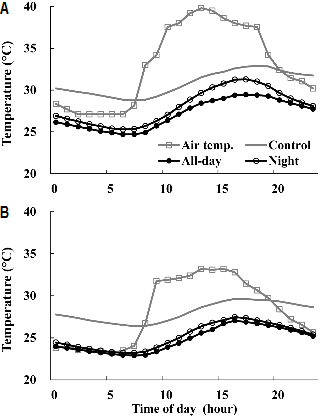
근권 냉방이 이루어지지 않은 대조구의 배지 온도는 낮에 시설로 유입된 복사에너지로 인해 일몰 이후까지 상승하였고, 야간 시간대 배지 온도가 평균 30.3°C(맑은 날)-27.5°C(흐린 날)의 고온을 유지하여 야간에도 배지 온도가 매우 높아 파프리카 근권 온도 범위에서 크게 벗어났다(Fig. 2). 이에 비해 전일과 야간 처리구의 근권 냉방은 일몰 이후 야간 시간대 배지 온도를 낮추는 데 긍정적인 영향인자로 작용하였다.
7월 23일부터 8월 31까지의 온실 내 일평균 온도는 30 ± 3.3°C, 최고/최저 평균온도는 37.7 ± 5.8°C/24.7 ± 2.1°C였으며, 8월 3일은 온실 최고온도가 48.8°C를 기록했다. 근권 냉방이 이루어지지 않은 대조구의 배지 일평균 온도는 29.1°C이였으며(Fig. 1), 맑은 날 야간 평균 온도도 30.3°C로 나타나(Fig. 2A), 이 기간 중 일평균 배지 온도가 25°C 이하로 내려간 날은 없었고, 30°C를 넘는 날은 16일이나 되었다(Fig. 1A). 이에 비해 전일 처리구에서는 일평균 배지 온도가 25.6°C, 야간 처리구에서는 26.1°C로, 일평균 배지 온도 25°C를 넘는 날이 전일 처리구에서는 23일, 야간 처리구에서는 27일 측정되었다.
파프리카 재배 온도는 21-26°C(Morgan, 2011)이며, 24시간 평균 온도 목표는 19-21°C임을 고려할 때 근권 냉방이 이루어지지 않은 대조구는 지상부와 지하부 모두 고온에서 파프리카가 재배되었음을 확인하였다. 식물 생육, 개화, 착과 및 과실 비대는 주/야간 온도뿐만 아니라 24시간 평균 온도가 크게 관여하고 있다(Bakker, 1989). 영양생장기와 착과 이후 온도 관리는 다르나, 고온(≥ 32°C)에서는 낙화 및 낙과를 조장하여 생산량을 감소시킬 수 있다(Khan and Passam, 1992; Rylski and Spigelman, 1982). 따라서 본 실험에서 고온기의 파프리카 근권 목표 최고 온도를 25°C로 가정할 때, 근권 냉방에 의해 배지의 고온 노출 기간을 전일에서는 42.5%, 야간 처리에서는 32.5% 줄일 수 있었다. 또한 본 실험에서 전일과 야간 처리구에서의 배지 내 하루 평균 온도는 0.5-1°C의 차이를 보였으나, 24시간 평균 온도를 고려한다면 전일 처리가 근권 배지의 고온 노출 일수를 감소시켜 고온기 근권 온도를 유지하는 데 보다 더 효과적이라 생각된다.
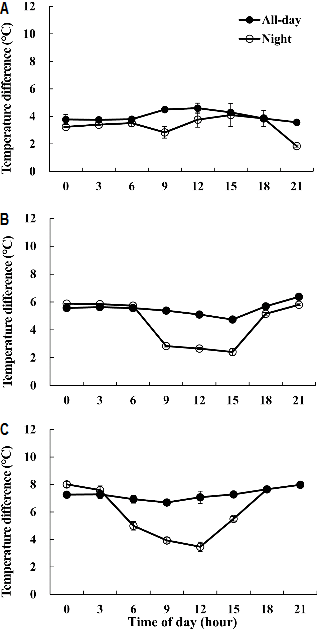
XL 냉수 순환 근권 냉방은 XL 파이프가 배지 하단부에 위치하므로 배지 깊이에 따른 온도 차[TD, TD = (대조구 - 처리구)의 배지 온도]와 평균 온도를 측정한 결과는 Fig. 3과 같다. 배지 상/중/하부의 TD가 전일 처리구에서는 4.0°C/ 5.5°C/7.3°C였으며, 야간 처리구에서는 3.3°C/4.5°C/6.1°C로, 배지 상부에서 하부로 갈수록 TD는 커졌다. 전일 처리구의 하루 중 TD 변화는 배지 상/중/하부에서 완만하였으나, 야간 처리구는 주간과 야간 시간대 TD가 배지 중, 하부에서 커졌다(Figs. 3B and 3C). 이는 냉방이 없었던 낮 시간 배지 온도 상승이 일몰과 함께 야간 냉방 공급으로 배지 온도가 하강함으로써 주간과 야간 시간 TD는 XL 파이프가 매설된 배지 중, 하부에서 큰 차이를 보인 것이다. 이러한 결과는 Lee et al.(2002)이 펄라이트 배지에서 XL 냉수순환, 라디에이터 냉풍, 패트 냉풍을 이용하여 근권 냉방 하였을 때 대조구에 비해 배지 온도가 낮아졌으며, XL 냉수순환에서는 배지 깊이별 근권 온도에 차이를 보여 본 실험 결과와도 같았다. 또한 배지 내 작물 근권 온도는 뿌리 발달뿐 아니라 양・수분 흡수에도 영향을 주며, 급액 관리에 따라 뿌리 분포는 달라질 수 있다(Kim et al., 2011; Lee et al., 2002). 따라서 본 실험을 통해 배지 깊이에 따른 근권 온도 차이가 있으므로, 뿌리 발달 등 생육과 양・수분 흡수와의 관계를 급액 관리 요인과 함께 병행하여 추후 연구해야 할 것으로 본다.
처리 25일째 전일 처리구에서의 뿌리 활력은 대조구와 야간 처리구에 비해 2배 이상 유의하게 높았으나, 야간 처리구는 대조구와 차이가 없었다(Table 1). 이는 근권온도가 높았던 시간이 야간 처리구가 전일 처리구보다 많았기 때문으로 생각된다.
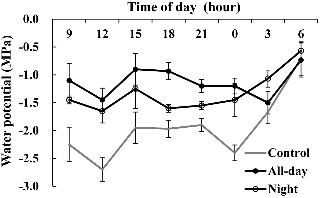
근권 냉방에 영향을 받은 전일과 야간 처리구의 잎 수분 포텐셜은 높았다(Fig. 4). 대조구의 잎 수분 포텐셜은 낮 시간 동안 전일 처리구에 비해 1.2MPa 낮았다. 하루 중 수분 포텐셜은 정오인 12시에 감소하고 오전 6시에 증가하였다. 오전 중 파프리카 엽온은 오후에 비해 낮았고 광합성과 증산율은 근권 냉방 처리에서 높았다(자료 미제시). 정오 12시는 지상부 온도와 함께 엽온이 상승하고, 한편으로 잎 표면으로부터 증발산이 증가함에 따라 오전 9시에 비해 수분 포텐셜이 감소한 것으로 보인다. 낮 시간 동안 잎 수분 포텐셜은 전일 처리구가 야간 처리구보다 높았다. 즉, 고온기 파프리카 생육기간 중 근권 냉방은 대조구에 비해 배지 온도를 낮춰(3-3.5°C; Fig. 1), 뿌리 근활력과 낮 시간 잎 수분포텐셜에 긍정적 영향을 준 것으로 보인다. 이는 Moon et al.(2007)이 근권 온도에 따른 오이의 잎 수분 포텐셜을 측정한 결과 20°C 근권 온도 처리구에서 가장 높고 고온인 35°C 포텐셜에 부의 영향을 준 것으로 본 실험결과와도 같았다.
파프리카 개화 시기는 고온기에 지연되어, 9월 7일에 전일 처리에서 첫 개화가 시작되었다(Table 2). 근권 냉방 처리구는 대조구에 비해 개화시기가 4-5일 앞당겨졌으며, 파프리카 착과 수도 유의하게 증가하였다. 이러한 결과는 Lee et al.(2002)이 근권 냉방으로 근권부를 25°C 이하로 유지하였을 때 토마토 화방별 착과율이 대조구에 비해 통계적 유의성이 인정되고 수량도 증가한 것으로, Jang et al.(2010)도 파프리카의 양액 냉각 처리로 과실 착과율과 A등급 수량이 증가하였다고 보고하여 유사한 결과를 얻었다.
한편, 주간 온도가 ≥ 32°C 고온인 경우는 착과가 감소한다는 Rylski and Spigelman(1982), Khan and Passam(1992)의 결과와 같이, 본 실험에서 근권 냉방에 의해 일평균 배지 온도 3-3.5°C 및 냉방 시간 처리간에는 평균 ≥ 0.5°C 이상 근권 온도 하강 효과를 보였으나, 고온기 온실 내 일평균 온도가 29.6°C로 높아 파프리카 착과가 늦게 이루어졌다. 이는 근권 냉방으로 배지 온도를 낮추어 뿌리 활력, 수분 포텐셜 등 생리적 반응에 유의하더라도 지상부 고온에 의한 개화 지연으로 고온기의 근권 온도 보상 효과를 찾을 수는 없었다. Behboudian et al.(1994)은 파프리카를 지온 34°C에 계속 노출시켰을 때 무기이온 흡수 양상, 뿌리 발달, 엽록소 함량은 품종에 따라 차이를 보였으며, 주/야간 고온의 배지 온도에서 계속적으로 재배되지 않으면 근권의 고온이 작물 생리적 반응에 영향을 주지 않을 수 있다고 시사했다. Gosselin and Trudel(1985)도 근권 온도에 의해 오이 생육은 증가하더라도 지상부 온도가 야간 저온에서는 생산성에 영향을 주지 못했다고 보고한 바 있다.
따라서 고온기 근권 냉방은 차광, 환기 면적 확대 등 주간 온도를 낮추면서 근권 냉방을 병행했을 때 냉방 효과를 더 높일 수 있으리라 본다.
